
Abstract
Avian species, as part of the higher-order vertebrates and situated at the upper trophic levels of the food chain, are particularly vulnerable to environmental pollutants owing to their potential for bioenrichment. This physiological characteristic renders birds as sensitive indicators of ecosystem health and contaminant exposure. In this study, sampling was done in the Saunders’s Gulls nesting grounds and feeding regions in the Lijin Field of the Yellow River Delta Nature Reserve in May 2018 and April 2019. Deceased individuals of Saunders’s Gulls were collected, totaling 15 nestlings (8 females and 7 males) and 3 adult female Saunders’s Gulls. We determined the concentrations of cadmium (Cd), iron (Fe), nickel (Ni), manganese (Mn), copper (Cu), chromium (Cr), and lead (Pb) in various tissues—including the heart, sternum, muscles, kidneys, lungs, intestines, and liver—of Saunders’s Gulls across different age and sex groups. Tissues were processed by drying and grinding, hydrogen peroxide digestion, and metal concentrations in different tissues were compared using atomic absorption spectrometry. Pearson correlation analysis and principal component analysis were used to understand the effects of different metals on gulls of different ages and sexes. Our findings indicated that, on average, Fe was the most abundant metal in all tissues of Saunders’s Gulls. The content of Fe in all tissues of female Saunders’s Gull nestlings ranged from 625.919 to 1,352.175 mg/kg, and in female adult Saunders’s Gulls, the content of Fe in the tissues confined 1,167.484–2,969.771 mg/kg. Notably, Ni was the least abundant in the intestines of adult birds, ranging from 0.926 to 1.572 mg/kg. We observed higher Fe concentrations in adult birds compared with nestlings, with the liver being the primary site of Fe accumulation in both female nestlings and adults. The study revealed that different tissues of Saunders’s Gulls have varying capacities for metal accumulation, with the liver, kidneys, and muscles showing the highest levels. Additionally, we found that different age and sex groups exhibit distinct metal concentration profiles in their tissues. Juvenile gulls were more likely to accumulate metals such as Mn, Cu, and Fe, whereas adult gulls were more affected by Cr and Pb. This research contributes valuable insights into metal pollution assessment in the Yellow River Delta wetlands, aiding in the ecological monitoring and management of metal pollution in this region.
Keywords
Introduction
Coastal wetland ecosystems have suffered damage recently as a result of the increased human production activities and the effects of climate change worldwide (Abbasi et al., 2015), which have enhanced the long-term effects of metals as environmental pollutants (Burger et al., 2004). Wetlands are considered as distinctive ecosystems and serve as “hotspots” for the accumulation of metals (Rossetti et al., 2016), which are generated by anthropogenic activities in production and daily life. Metals can enter wetlands through various pathways such as surface runoff and atmospheric deposition (Ramos-Miras et al., 2011). As hazardous and persistent pollutants, metals have traits like bioaccumulation (Fe, Mn, etc.), biomagnification (Hg, Cd, Pb, etc.), and resistance to degradation (Rashid et al., 2022). These properties increase their ability to harm organisms. For example, plant bodies can become migrated by metals from soil organic materials, which can impact plant physiological and ecological processes and lower wetland output (Liu et al., 2006). Additionally, metals can build up in animals through the food chain and affect their development, growth, and ability to reproduce. In addition to posing a serious threat to the ecosystem as a whole, metals can potentially cause a death of organisms (Hao et al., 2011). For example, excessive consumption of external Cu poisoned and killed eight mute swans (Cygnus olor) in Tottori Prefecture, Japan (Kobayashi et al., 1992). At many levels of biological organization, metals have detrimental effects that possess the ability to reach the population level (Das et al., 2002). Thus, maintaining a healthy ecosystem serves as protection for both human populations and biota within the ecosystem, and it would make sense to identify appropriate indicators to comprehend the effects of metals on biological groups and the ecology of wetlands.
The Yellow River Delta (YRD) wetlands are the most intact and extensive coastal wetlands in the temperate zone of China (Ning et al., 2023). The diversity of the wetland ecosystem is a defining feature, shaped by the dual influences of freshwater and brackish water from the river and various geological and anthropogenic factors. The YRD region is rich in mineral resources, primarily focused on oil and gas (Zhang and Sun, 2005). While the development of petroleum resources brings economic benefits to the local area, it also has certain impacts on the wetland environment of the YRD. The ecological and environmental issues in the region have gained more attention recently due to the rapid development of agriculture and industrial production in the YRD, increasing efforts in oil field development, rising sea levels, coastal erosion, and decreased upstream water inflow, among other factors like soil salinization. Among these issues, metal pollution has drawn widespread attention from scholars (Tan et al., 2014). Numerous researchers have studied the distribution of metals in the soil and conducted corresponding environmental assessments (Gan et al., 2019). However, examining metals just in soil, water, or even the air is insufficient in environmental monitoring studies to evaluate the possible effects of metals on locals and wildlife. Birds are highly sensitive to anthropogenic pollution and serve as valuable bioindicators for monitoring metal pollutants (Fu et al., 2014). Therefore, compared with measurements made in the physical environment, on plants, and invertebrates, measurements made on metals in birds can offer a greater knowledge of their potential impact on humans.
As top consumers in the food chain, when birds forage on benthic or planktonic organisms, metals from wetland soils can enter their bodies through the food chain, potentially affecting the health of birds (Varela et al., 2016). Previous researches have primarily focused on the distribution and sources of metals in the YRD (Miao et al., 2020), with limited studies investigating the accumulation of metals in birds (Yang et al., 2022). Hence, it is crucial to study the impact of metals on birds and implement measures to protect the bird species and their primary habitats in the YRD.
This study focused on the Saunders’s Gull (Larus saundersi) in the YRD as the research subjects. Our research aimed to (1) investigate the distribution characteristics of metals in different tissues of Saunders’s Gulls of various sex, (2) study the distribution characteristics of metals in different tissues of Saunders’s Gulls of various ages, and (3) explore the enrichment patterns of different metals in various tissues. This study establishes the groundwork for the biological monitoring of metal pollution and its management in the YRD ecosystem, in addition to offering insights into the pollution of metals in the coastal wetlands of the region and its effects on the local bird species.
Materials and Methods
Study area and study species
The YRD wetlands are located at the confluence of Laizhou Bay and Bohai Bay in the northeastern part of Shandong Province, China. It lies between ∼118°33′ to 119°20′ east longitude and 37°35′ to 38°12′ north latitude, covering a total area of 153,000 hectares. The YRD features a vast, flat terrain, faces the sea on its eastern side, and experiences relatively abundant precipitation (Huang et al., 2022). It is located in the zone of temperate continental monsoon climate, which is characterized by four distinct seasons (Jiang et al., 2013). In addition to being a typical coastal wetland ecology, YRD is a major hub for petroleum extraction (Zou et al., 2020). Rich in mineral resources, mostly concentrated on oil and gas reserves, the protected area is peppered with numerous oil wells of varying sizes (Yue et al., 2003). With a wide variety of wild plant species, the vegetation in the area has a natural succession pattern resembling coastal wetlands (Li et al., 2021). Additionally, the delta is home to a wealth of bird species, including many nationally rare and endangered birds such as the Oriental Stork (Ciconia boyciana), Golden Eagle (Aquila chrysaetos), and Red-crowned Crane (Grus japonensis) (Hua et al., 2012). Furthermore, the YRD serves as an important stopover and wintering ground, and breeding site for birds during their migration in the East Asia inland and around the western Pacific region (Higuchi, 2012).
The research object of this study is Saunders’s Gulls, which have been categorized as a vulnerable species by the International Union for Conservation of Nature and were designated as a first-level protected animal in China in 2021 (Xu et al., 2021).
Sample collection
Sampling was done in the Saunders’s Gulls nesting grounds and feeding regions in the Lijin Field of the YRD Nature Reserve in May 2018 and April 2019. Deceased individuals of Saunders’s Gulls were collected, totaling 15 nestlings (8 females and 7 males) and 3 adult female Saunders’s Gulls. Each individual was assigned a unique identification number and stored in sealed bags under freezing conditions. Detailed sample information of 18 birds is presented in Table 1. The sampling sites were illustrated in Figure 1. During the sampling period, the number of adult males in the study area was limited, resulting in adult males not being collected. This may also be related to our sampling time and the behavioral patterns of adult males. We will focus on this aspect in our subsequent research.
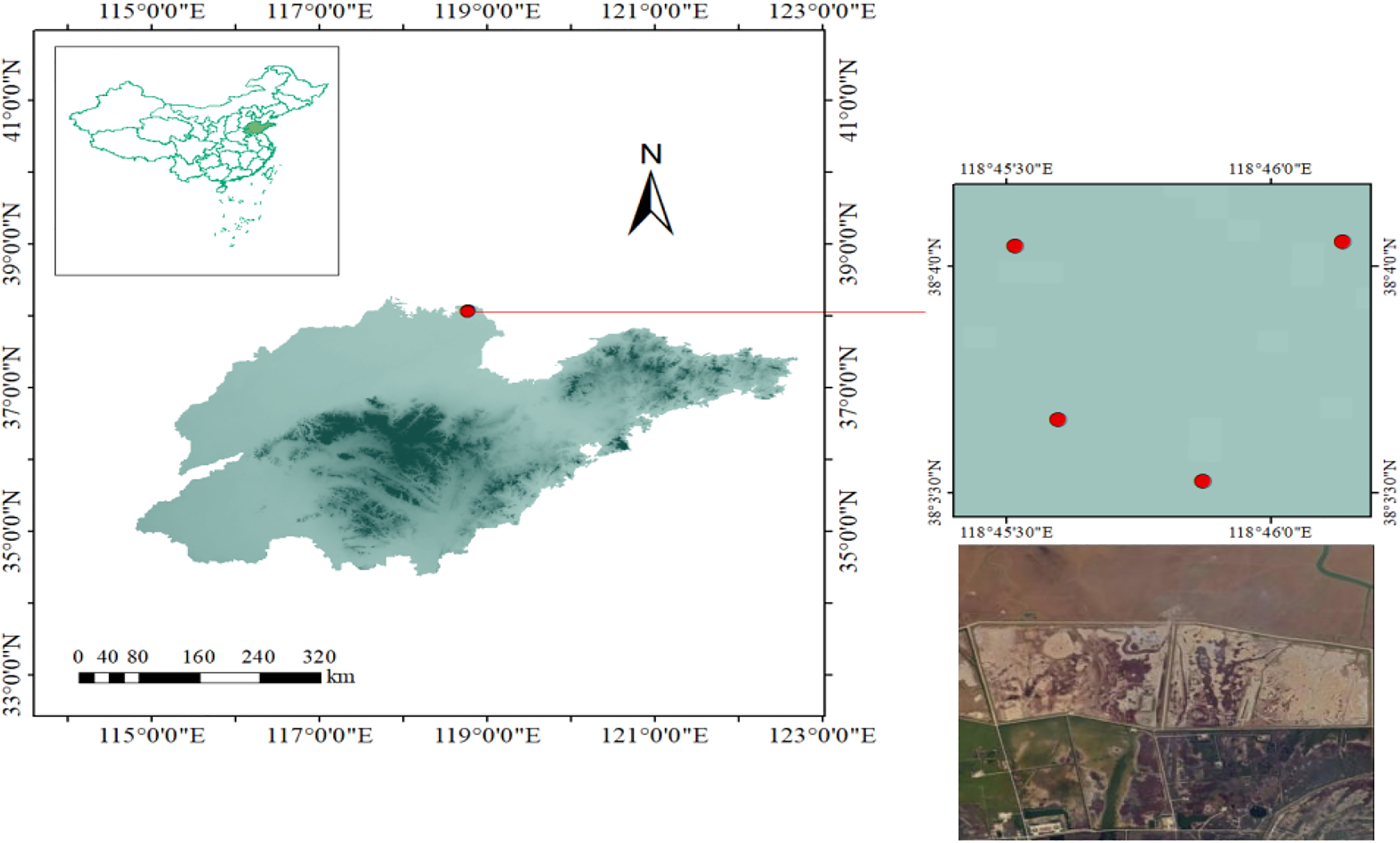
Sampling sites in the Yellow River Delta, China.
Detailed Information of Each Individual
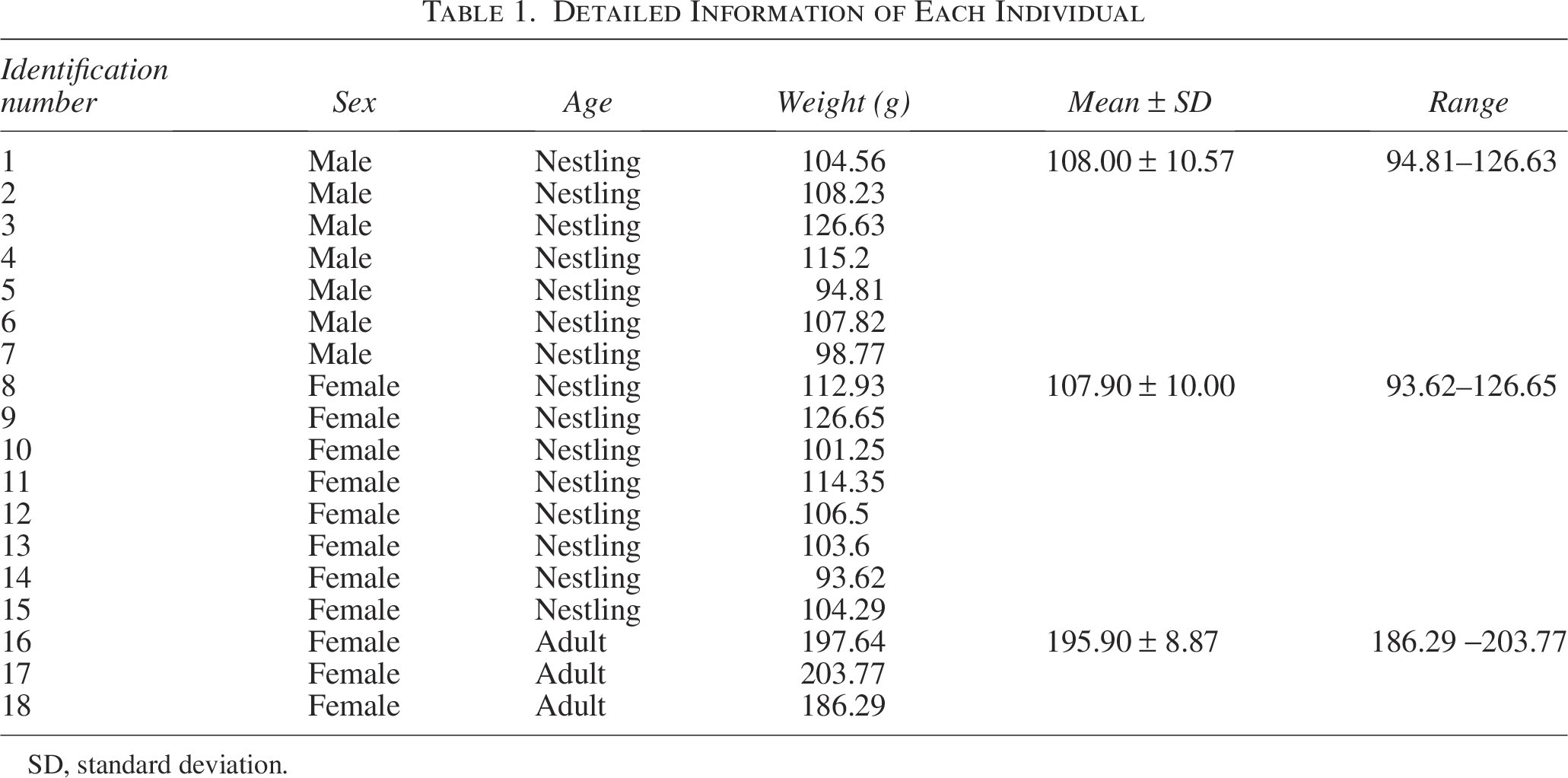
SD, standard deviation.
Laboratory analysis
The bird feathers were wetted with water and the Saunders’s Gulls were dissected with stainless steel scissors to isolate the tissues and organs such as heart, sternum, muscles, kidneys, lungs, intestines, and liver. The dissecting tools were correctly cleaned from one bird to another to ensure that there was no cross-contamination. Each dissected tissue organ was first washed with tap water to remove blood stains on the surface of each tissue, and then washed thoroughly with distilled water and ultrapure water in turn, respectively, and the tissue surfaces were drained of water. After dissecting Saunders’s Gulls and separating the heart, sternum, muscles, kidneys, lungs, intestines, and liver, and removing the bloodstains from their surface, we labeled each tissue sample from each bird, took in a drying oven, and adjusted the temperature to 105 ± 5°C for 48 h. Drying them and cooled to room temperature naturally, we ground the organ tissues into a powder in a mortar and pestle. Each sample was precisely weighed with an electronic balance to yield a 0.05 g weight before being put into a PTFE digestive tube (Yang et al., 2020). Subsequently, 12 mL of nitric acid and 4 mL of hydrogen peroxide were added to the digestion tube. After that, the PTFE digestion tube was placed on a graphite heating digestion device (model: XU-108-12).
For a 30-min predigestion phase, the temperature of the apparatus was initially increased to 90°C. After that, the temperature was raised to 150°C to continue the digestive process until the sample solution became clear, signifying that it was finished (Hseu, 2004). Then the heating apparatus was set to 160°C for acid evaporation. Acid evaporation continued until there was ∼2 mL of solution remaining. The digestion tube was then removed and allowed to naturally cool. The sample solution was filtered into a 50 mL volumetric flask using a 0.45 μm membrane. The inner walls of the digestion tube were rinsed 3–4 times with distilled water, and these rinses were combined with the filtrate. The tissue samples, filtered through a 0.45 μm membrane, were analyzed using an ice-3400 Atomic Absorption Spectrophotometer (Sun et al., 2006).
Sample quality control
To ensure the accuracy, consistency, and reliability of all morphometric data in this study, we implemented quality control procedures. All field data collection and morphometric measurements were conducted by four fixed and experienced researchers. Prior to the start of the study, uniform training was conducted to ensure consistent operational standards and minimize human error. Before the measurements, the electronic balance (model: ME204E/02) was calibrated using standard weights to ensure weight measurement accuracy to 0.01 g. Weight measurements were conducted during the same time period, and all measurements were performed on a flat, stable surface. Measurement data were recorded in real time and cross-checked after completion to avoid transcription errors. After data entry into the electronic database, sorting and conditional formatting functions were used to check for outliers, logical errors, or values significantly outside the known range for the species. No outliers were identified in this study. All laboratory equipment and solvents were tested to prevent contamination.
Statistical analysis
The data obtained from the measurements were subjected to basic analysis using Microsoft Excel 2019. These data were verified by the Shapiro–Wilk test to be normally distributed (all p > 0.05), and Levene’s test indicated homogeneity of variance (p = 0.32, 0.14). Therefore, we used the independent samples t-test to compare the mean differences between the two groups; one-way analysis of variance was used for group comparisons, and Tukey’s HSD method was employed for post hoc tests. The analyses were conducted using SPSS 22.0 and Origin software.
Results
Metal concentration in tissues of female Saunders’s Gull nestlings
A uniform distribution of metals across all tissues was observed. Notably, Fe exhibited the highest mean concentration in all tissues, with a value of 853.35 mg/kg, while Cd had the lowest, at 0.1761 mg/kg (Fig. 2). The proportional differences in concentration between each tissue type were shown in Figure 3. Statistically significant differences in Cd concentrations were detected among the kidney, heart, sternum, muscle, lung, and liver (p < 0.01) (Fig. 3a). Cr concentrations demonstrated highly significant differences between the muscle and both the kidney and lung (p = 0.004 and 0.005, respectively) (Fig. 3c). Cu concentrations were significantly higher in the liver compared with all other tissues (p < 0.01) (Fig. 3e). Pb concentrations varied significantly between the sternum and both the intestine and heart (p = 0.044 and 0.048, respectively), with liver Pb levels being significantly higher than those in the heart and intestine (p = 0.015 and 0.014, respectively) (Fig. 3f). Moreover, both Fe and Mn showed highly significant differences in concentrations between the liver and other tissues (p < 0.001) (Fig. 3d, g). While Fe concentrations in the sternum, muscle, intestine, and lung did not exhibit significant differences, they were markedly different in all other tissues (Fig. 3g).
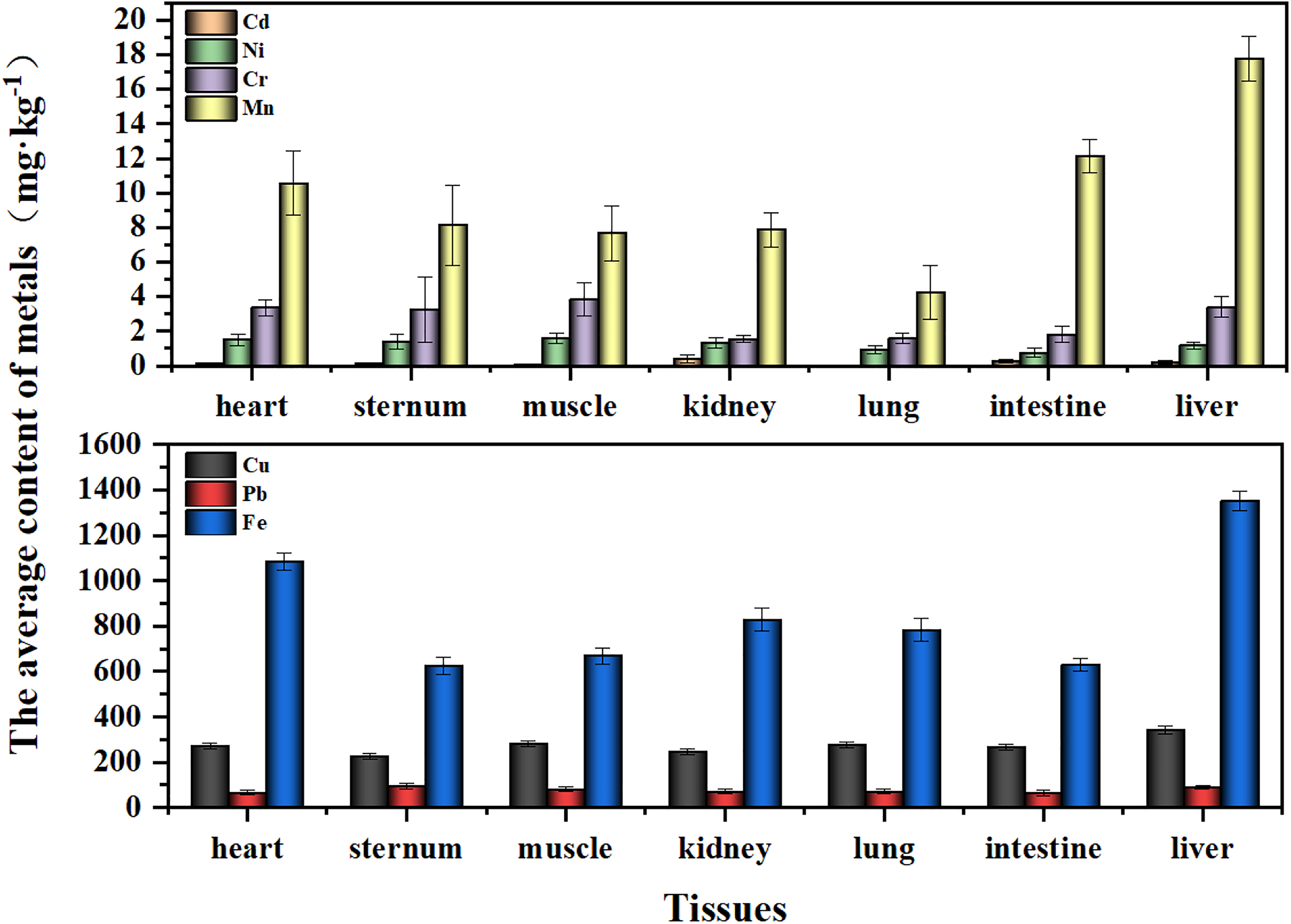
The average concentration of metals in the tissues of female Saunders’s Gulls nestlings.
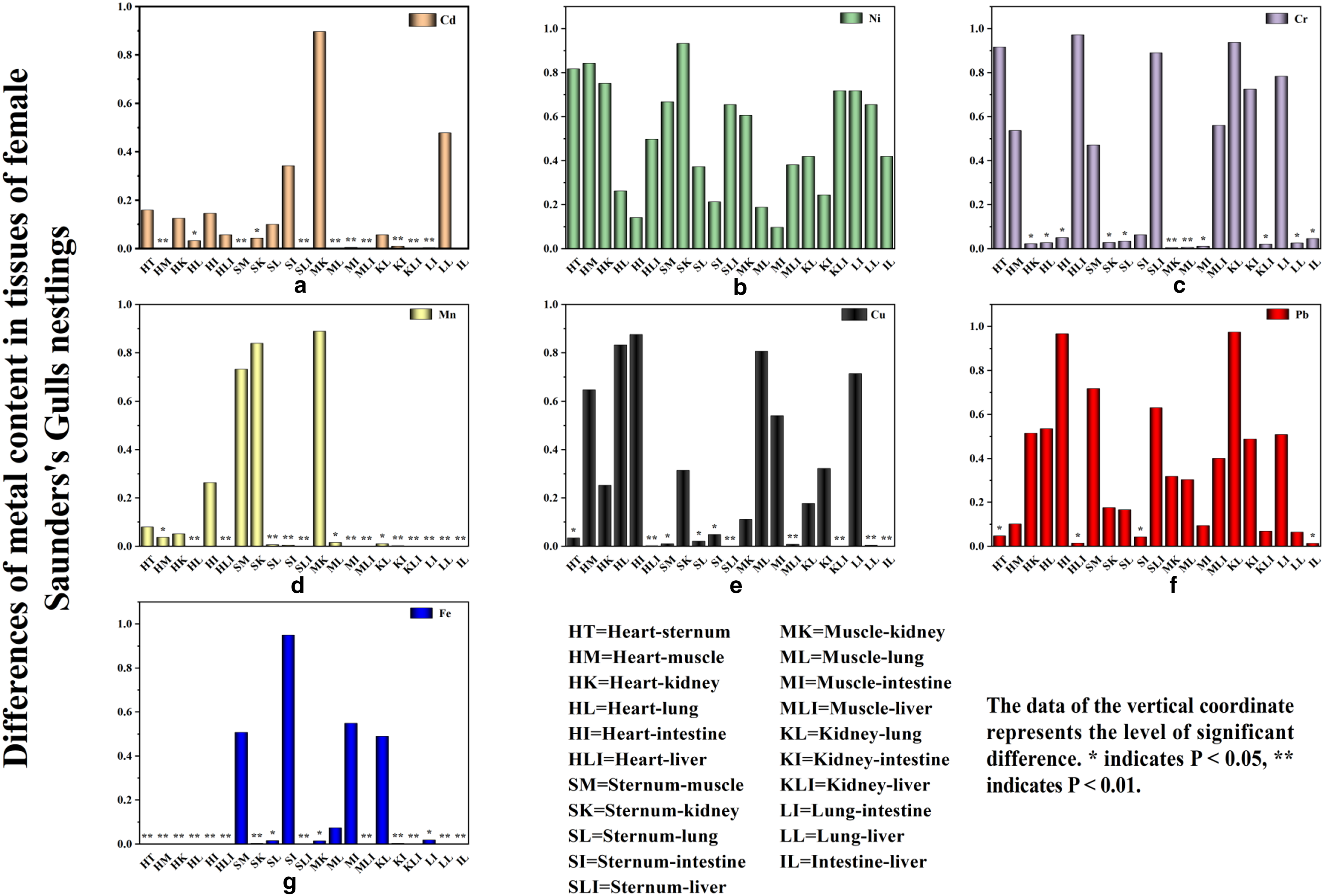
The differences of metal concentration in the tissues of female Saunders’s Gulls nestlings.
Metal concentration in tissues of female adult Saunders’s Gulls
The hierarchy of metal concentrations in tissues of female adult Saunders’s Gulls is as follows (Fig. 4). Fe was found to have the highest concentration across all tissues, whereas Cd had the lowest, which is consistent with the distribution pattern in nestlings, particularly in the heart, sternum, muscle, and lung. Notably, Ni concentrations exhibited extremely significant differences between the muscle and lung (p = 0.006), intestine (p = 0.004), and liver (p = 0.009), with a significant difference also observed between muscle and kidney (p = 0.028) (Fig. 5b). Moreover, Cr concentrations in muscle were significantly higher compared with the kidney, lung, and intestine (p < 0.001, p < 0.001, and p = 0.007, respectively) (Fig. 5c). Cu concentrations in the sternum were highly significantly different from those in the lung (p = 0.002) and liver (p = 0.01) (Fig. 5e). Pb concentrations in the sternum were highly significantly different from those in the heart, kidney, lung, and intestine (p = 0.004, p = 0.004, p = 0.006, and p = 0.001, respectively) (Fig. 5f). In addition, Mn concentrations showed significant differences between the sternum and all other tissues (p < 0.05) (Fig. 5d), and Fe concentrations displayed highly significant differences between the liver and all other tissues (p < 0.001) (Fig. 5g).
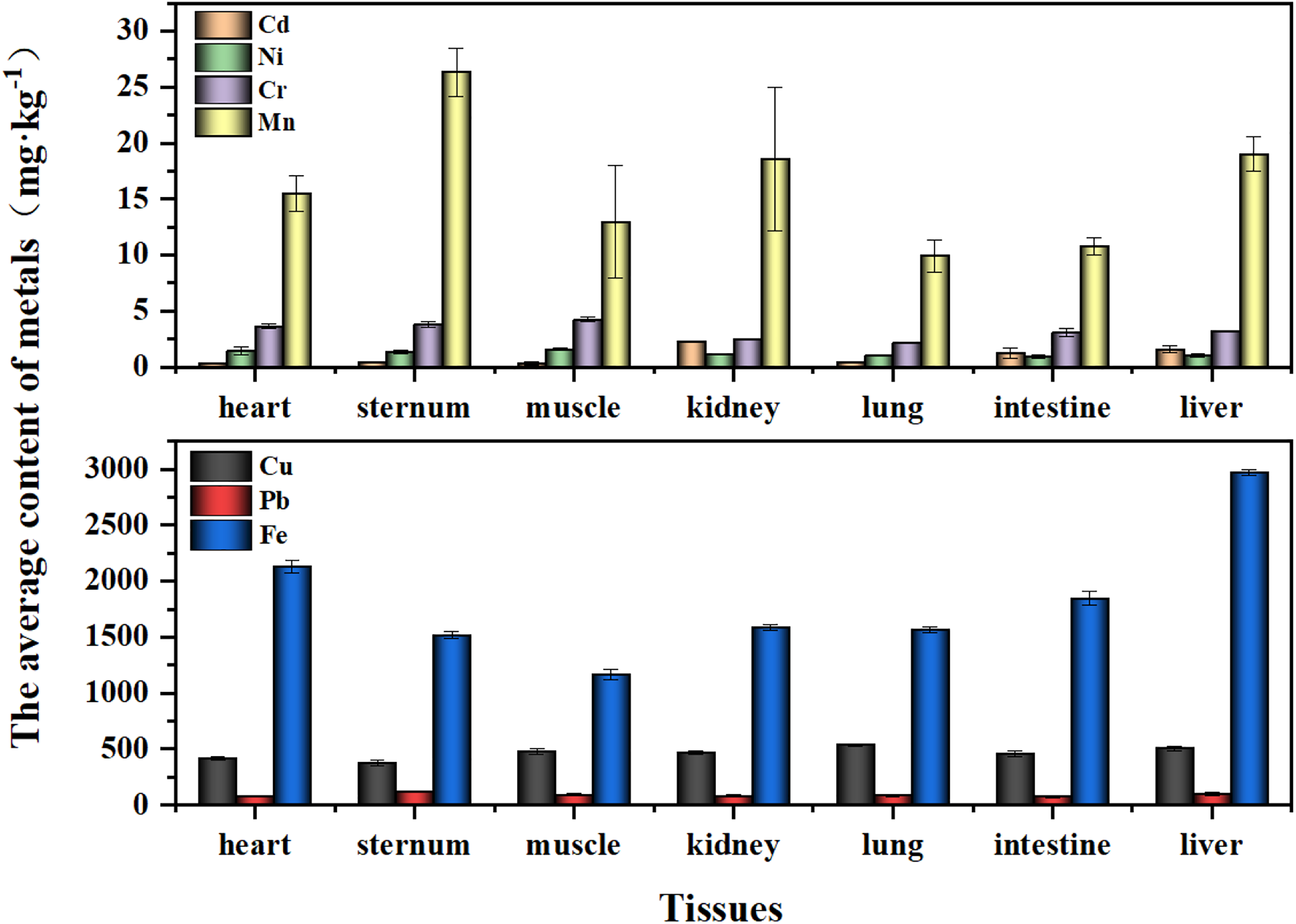
The average concentration of metals in the tissues of female adult Saunders’s Gulls.
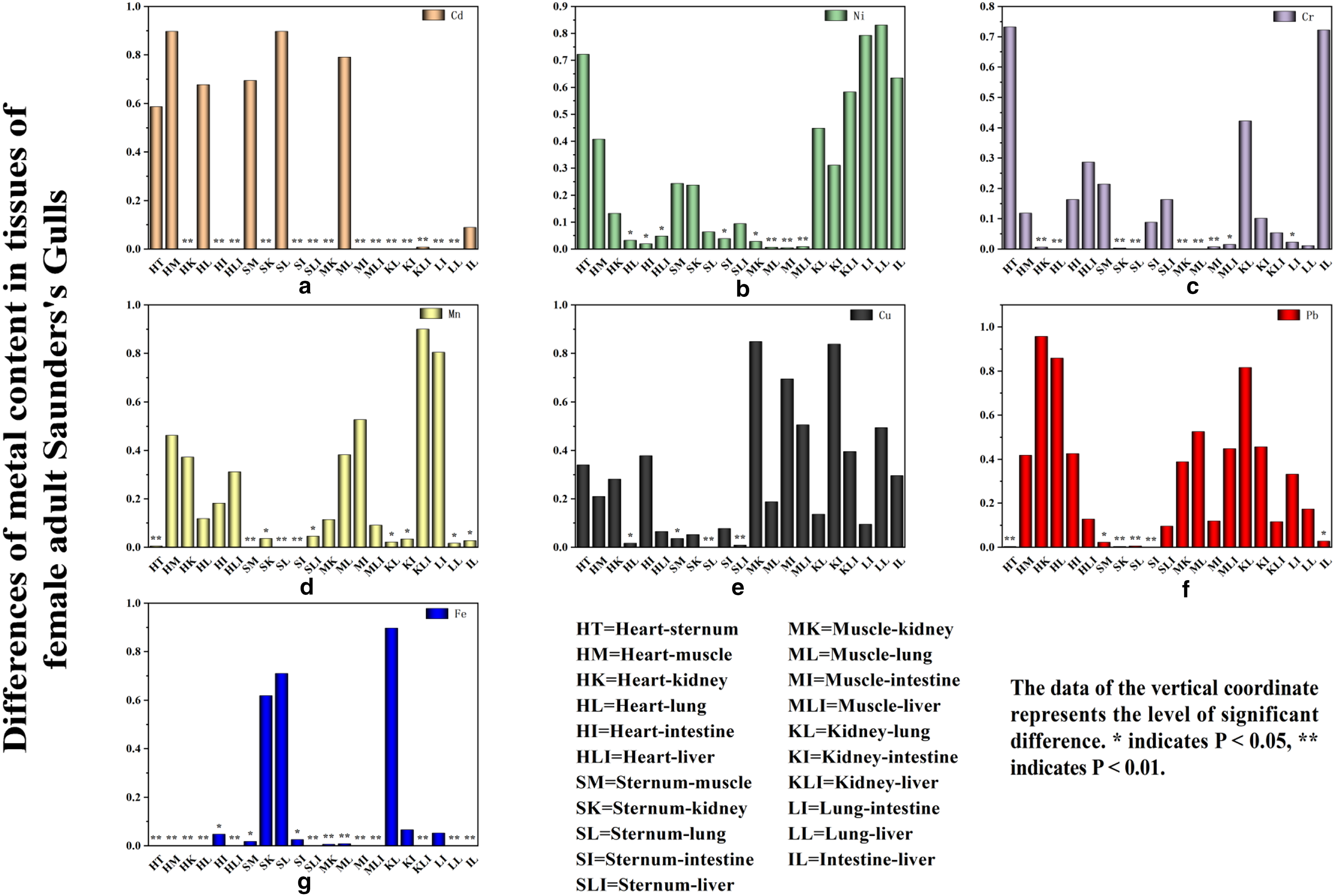
The differences of metal concentration in the tissues of female adult Saunders’s Gulls.
Comparison of metal levels in different tissues of Saunders’s Gull across different age groups
Our research has confirmed the variations in metal concentrations in tissues of Saunders’s Gulls across different age groups. Specifically, in the kidneys, intestines, and liver of adult birds, the metal concentrations were ordered as Fe > Cu > Pb > Mn > Cr > Cd > Ni (Fig. 4). In contrast, in other tissues of both juvenile and adult birds, the order was Fe > Cu > Pb > Mn > Cr > Ni > Cd (Figs. 2 and 6). Notably, in Saunders’s Gull nestlings, nickel (Ni) concentrations in the heart, sternum, muscles, kidney, and liver were found to be higher than those in adult birds (Figs. 2, 4, and 6). Conversely, metal concentrations in all other tissues were higher in adult birds compared with nestlings (Figs. 2, 4, and 6). It is noteworthy that there was no significant difference in Ni concentrations across all tissues between adult and young birds, whereas iron (Fe) concentrations exhibited significant variation in all tissues except muscle (Table 2).
The average concentration of metals in the tissues of male Saunders’s Gulls nestlings.
Comparison of Metal Concentration in Tissues of Saunders’s Gulls of Different Ages
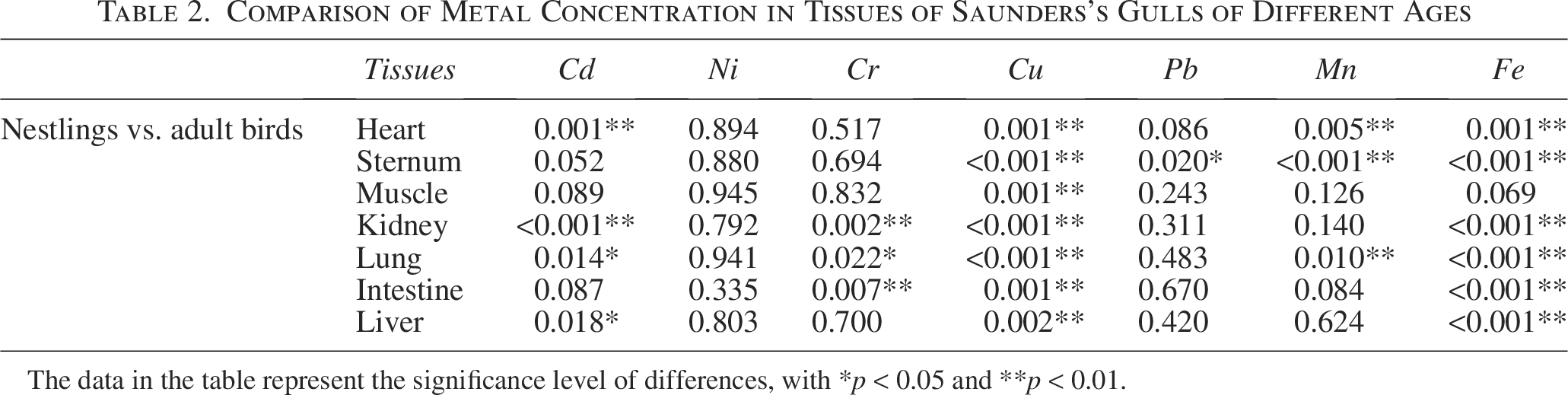
The data in the table represent the significance level of differences, with *p < 0.05 and **p < 0.01.
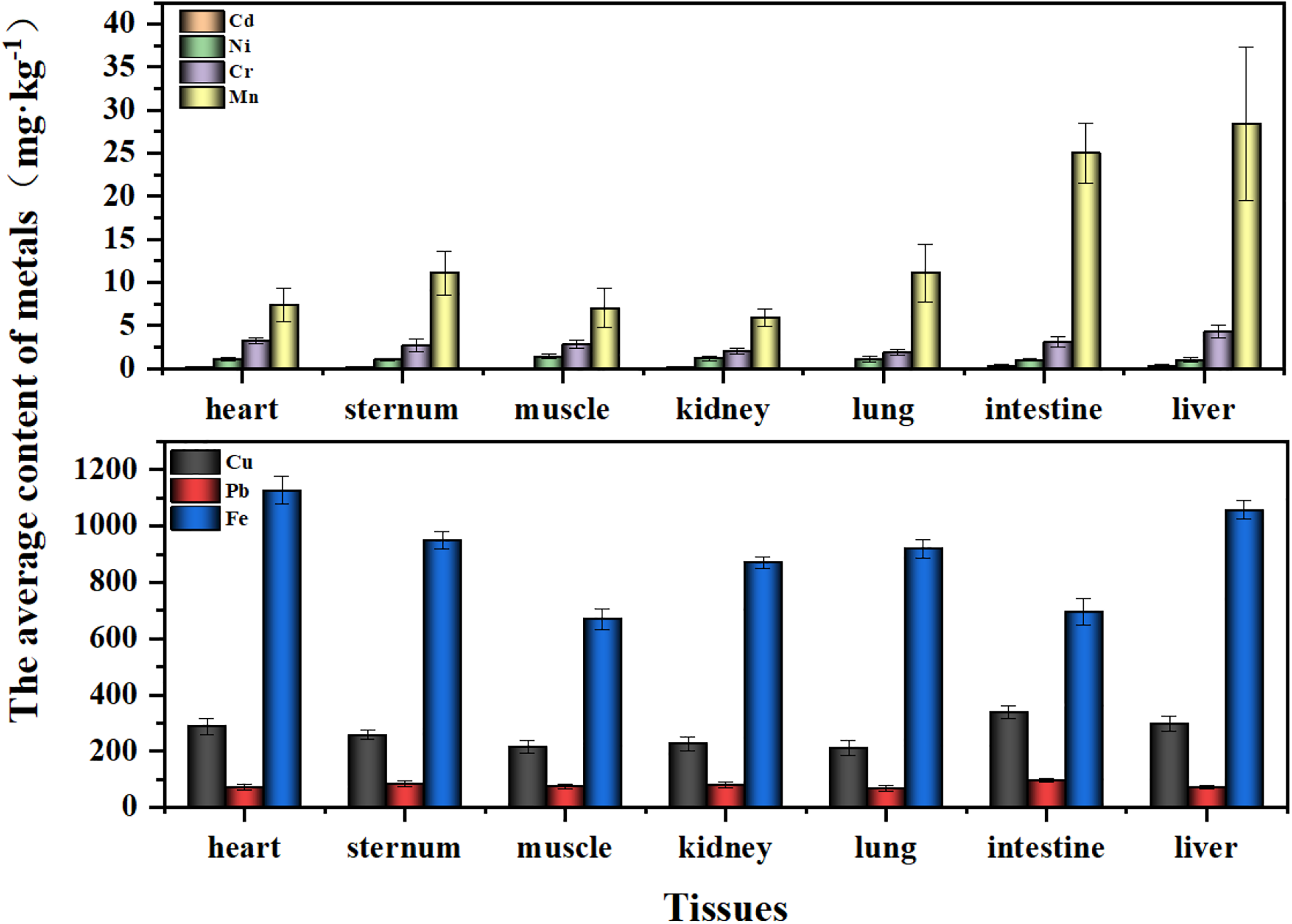
Metal concentration in tissues of male Saunders’s Gull nestlings
Analysis of metal concentrations across various tissues in juvenile male Saunders’s Gulls revealed that Fe had the highest concentration, whereas Cd had the lowest, with a uniform distribution observed across all tissues, as illustrated in Figure 6.
In the comparison of Cd levels between the intestine and liver of male Saunders’s Gull nestlings, no significant difference was observed (p = 0.635). However, both tissues displayed highly significant differences in Cd concentrations when compared with the heart, muscle, sternum, and lung (p ≤ 0.01) (Fig. 7a). Ni levels exhibited significant variation specifically in the muscle and intestine (p = 0.039) (Fig. 7b). Cr concentrations were significantly different between the liver and the sternum, muscle, kidney, and lung (p = 0.002, p = 0.005, p < 0.001, p < 0.001, respectively), with striking differences also observed between heart and lung tissues (p = 0.009) (Fig. 7c). Cu levels in the intestine showed extremely significant differences compared with the sternum, muscle, kidney, and lung (p = 0.009, p < 0.001, p = 0.001, p < 0.001, respectively) (Fig. 7e). Pb levels exhibited extremely significant differences between the intestine and the heart, muscle, lung, and liver (p = 0.003, p = 0.005, p = 0.001, p = 0.002, respectively) (Fig. 7f). Mn concentrations in the liver were significantly higher compared with all other tissues (p < 0.001) (Fig. 7d). Fe levels demonstrated extremely significant differences in the heart compared with muscles and the intestine (p < 0.001) (Fig. 7g).
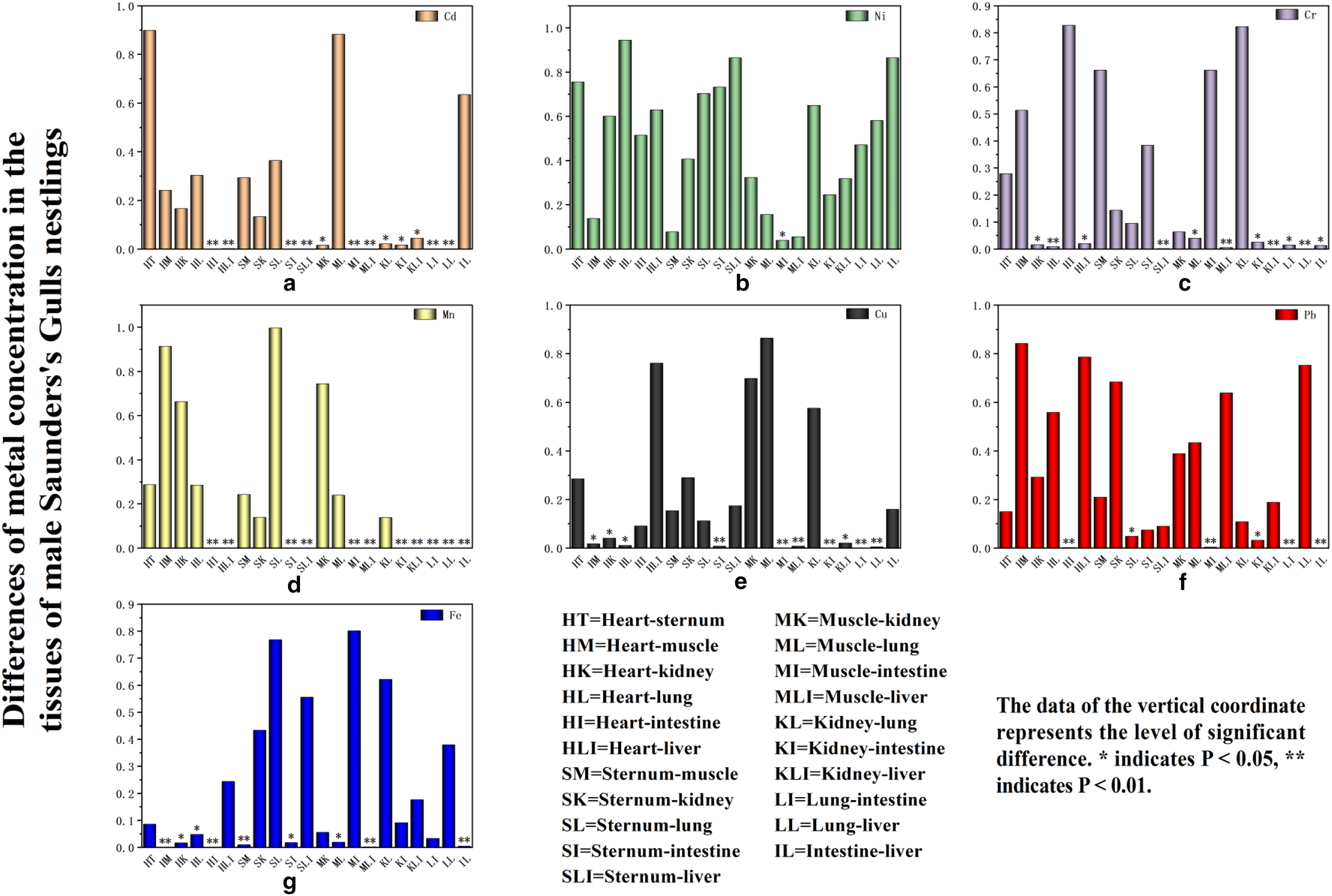
The differences of metal concentration in the tissues of male Saunders’s Gulls nestlings.
Comparison of metal concentration in different tissues of Saunders’s Gulls nestlings of different sex
Upon comparing metal concentrations in various tissues of male and female Saunders’s Gulls nestlings, a consistent pattern emerged, with the metals ordered as Fe > Cu > Pb > Mn > Cr > Ni > Cd. Notably, in the heart, female Saunders’s Gulls nestlings exhibited higher average concentrations of Cd, Ni, Cr, and Mn compared with male nestlings, with manganese (Mn) showing a significantly elevated concentration. Conversely, in male Saunders’s Gulls nestlings, the average concentrations of Cu, Pb, and Fe in the heart were higher than in females, albeit not significantly so. As detailed in Table 3, no significant differences were observed in Ni and Pb concentrations between male and female Saunders’s Gulls nestlings.
Comparison of Metal Concentration in Tissues of Saunders’s Gulls Nestlings of Different Sex
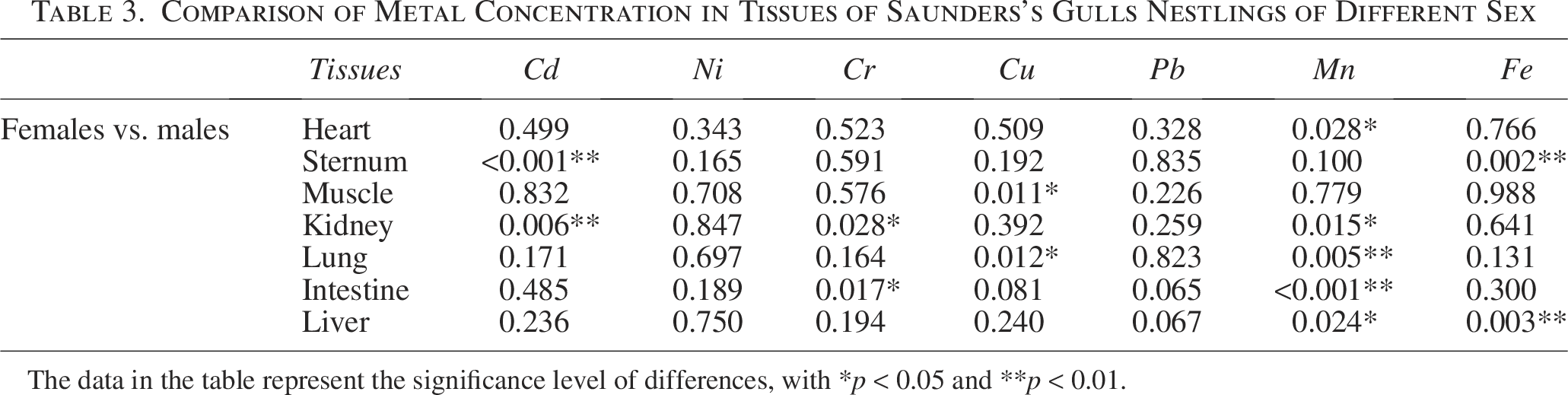
The data in the table represent the significance level of differences, with *p < 0.05 and **p < 0.01.
Pearson correlation analysis of metal concentration in different age groups of female Saunders’s Gulls’ intestines
Metals in the intestines of birds can serve as important bioindicators of wetland ecosystem health, and can indirectly reflect the condition of the wetland environment by analyzing their correlation and other characteristics. To understand the correlation of metal contents in the intestines of female Saunders’s Gulls nestlings and adult female Saunders’s Gulls, Pearson’s correlation analysis was carried out on the metal contents in the intestines of them, respectively; the Pearson correlation coefficient indicates the degree of correlation between metals. In the intestines of female Saunders’s Gulls nestlings, a negative correlation was identified between Cd and Ni, Cu, Cr, and Pb (Fig. 8a). Additionally, Ni and Cu, as well as Ni and Mn, exhibited negative correlations. In contrast, the remaining metals showed positive correlations. Among female adult Saunders’s Gulls, no significant correlations were detected among all metals. Furthermore, there was a very weak positive correlation between Fe and Pb in their intestines, with a correlation coefficient of 0.04 (Fig. 8b).
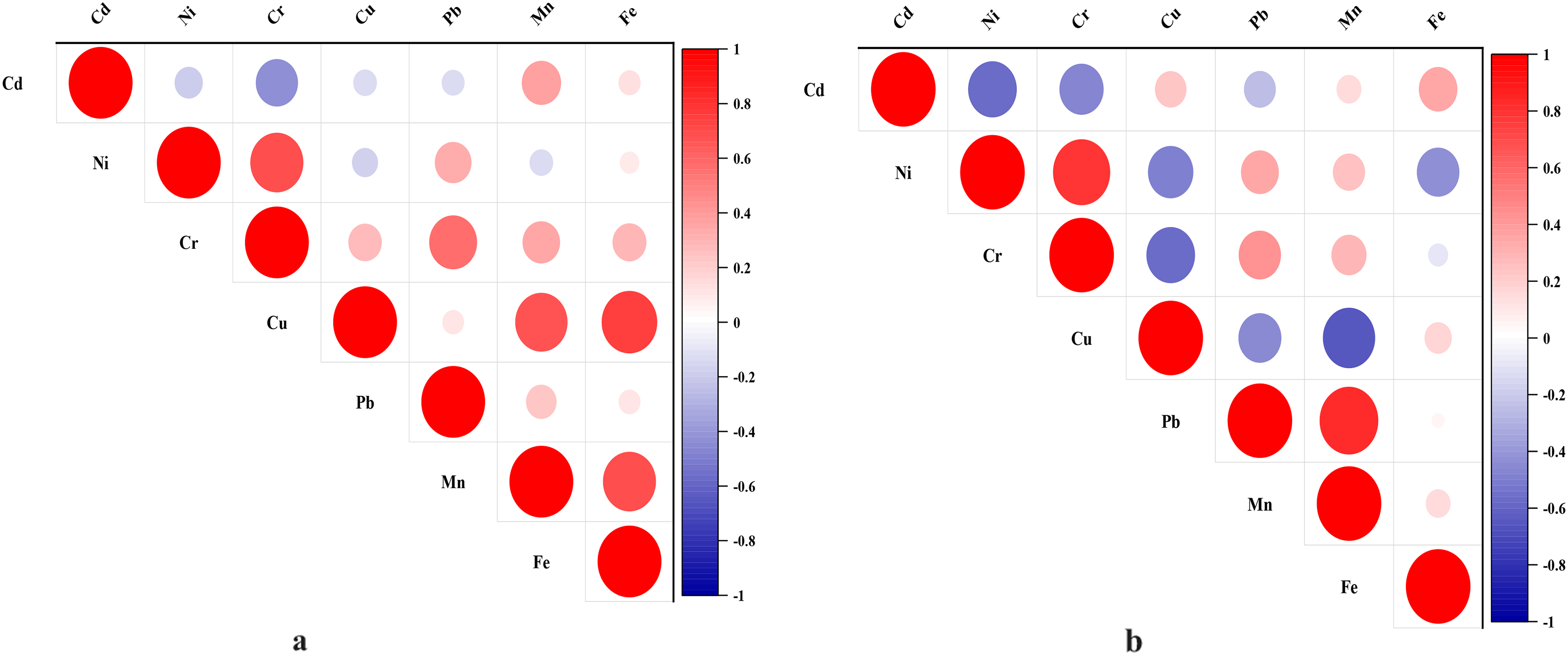
The correlation of metal concentration in tissues of female Saunders’s Gull nestlings and adults.
Principal component analysis of metals in different age groups of Saunders’s Gulls’ tissues
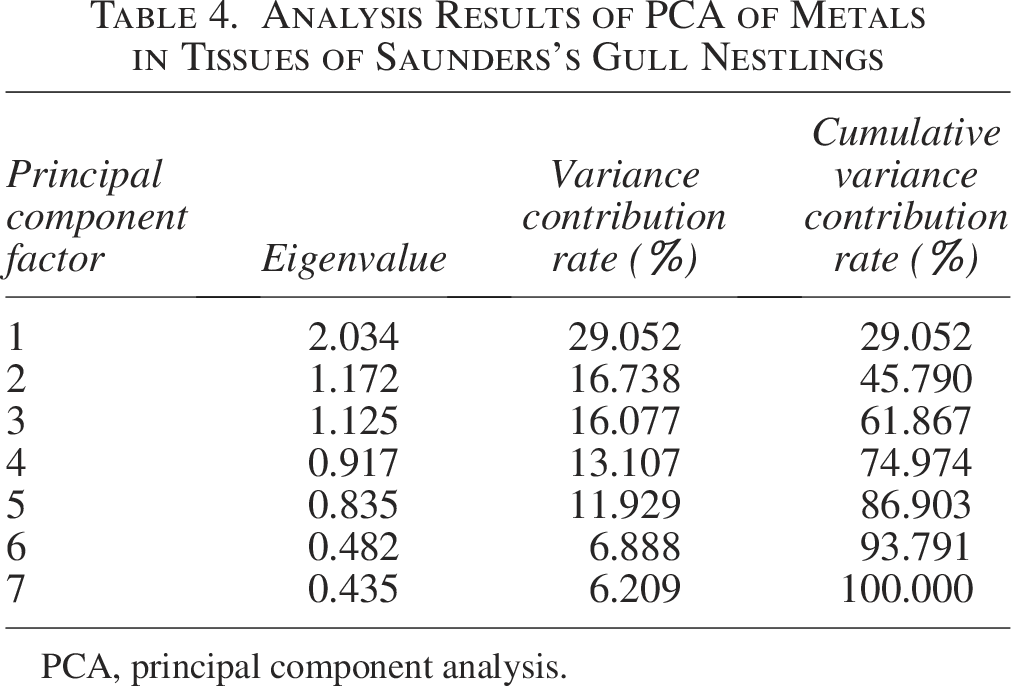
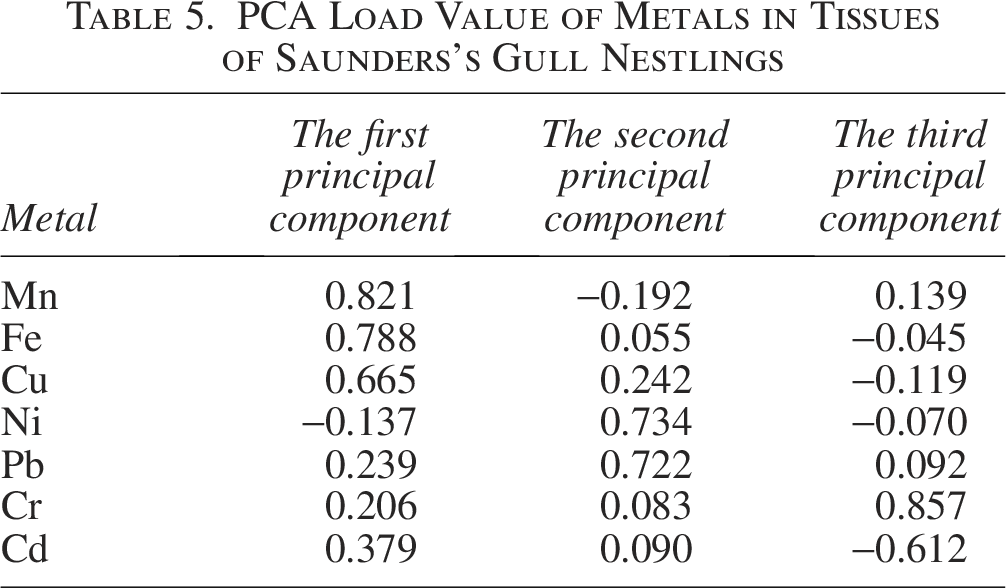
In the tissues of Saunders’s Gull nestlings, Mn, Cu, and Fe exhibited the strongest correlation with the first principal component, aligning with the observed positive correlations among these metals (Cu and Mn, Cu and Fe, Mn and Fe) as depicted in Figure 8a. The first principal component accounted for 29.052% of the variance. Ni and Pb demonstrated the highest correlation with the second principal component, which explained 16.738% of the variance, whereas Cr had the strongest correlation with the third principal component, contributing 16.077% to the total variance (Table 4). These contributions were markedly higher than those of the remaining principal components, suggesting that metal pollution in the tissues of Saunders’s Gull nestlings is predominantly governed by these three principal components.
Analysis Results of PCA of Metals in Tissues of Saunders’s Gull Nestlings
PCA, principal component analysis.
In parallel, the loading values for Mn, Cu, and Fe on the first principal component were notably higher compared with other metals (Table 5). Ni exhibited the highest loading value on the second principal component, with Pb following closely, while Cr demonstrated a relatively elevated loading value on the third principal component. The metal factor loadings in the tissues of Saunders’s Gull nestlings are graphically represented in Figure 9.
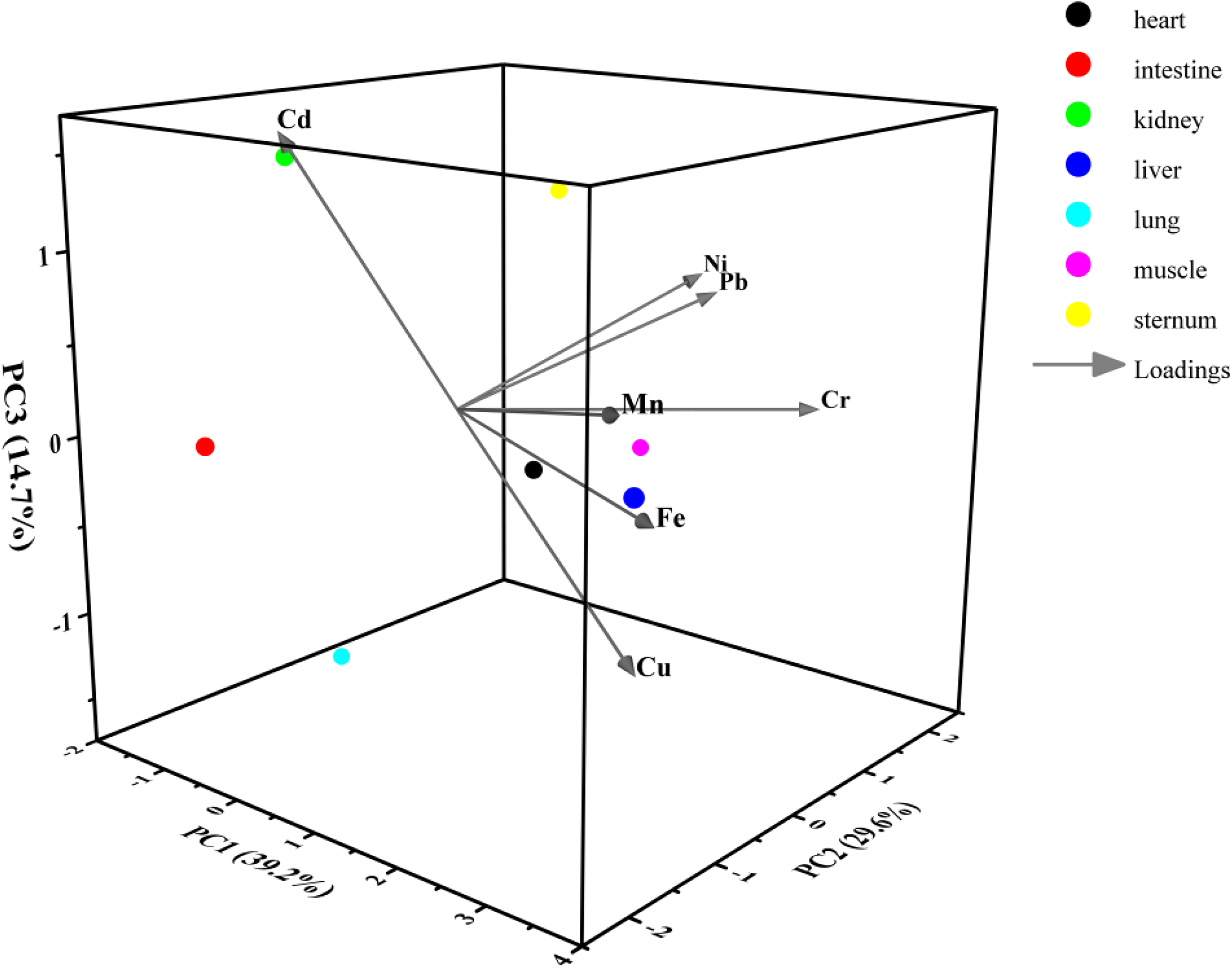
Metal factor loadings of the tissues of Saunders’s Gull nestlings.
PCA Load Value of Metals in Tissues of Saunders’s Gull Nestlings
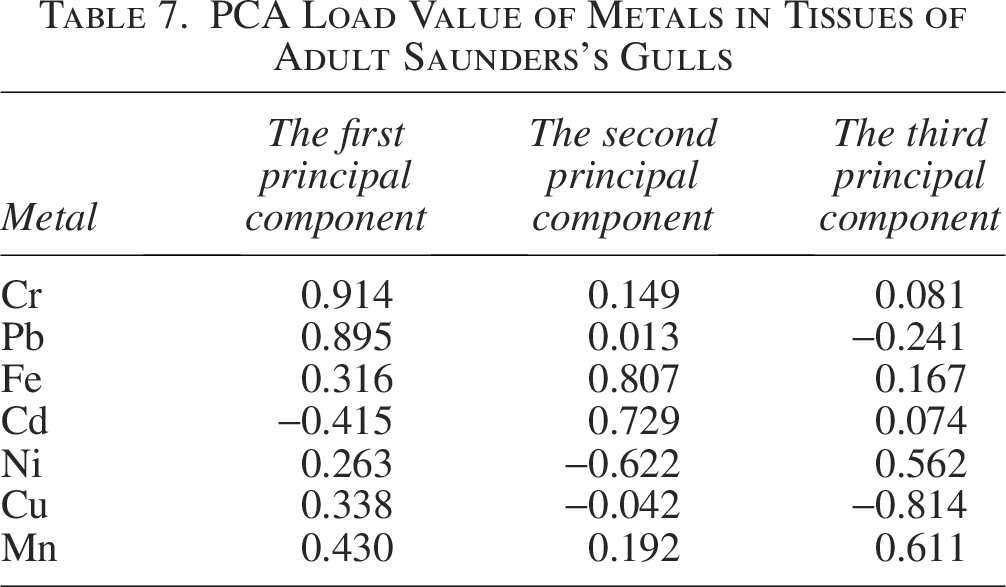
In adult Saunders’s Gulls, the first principal component accounted for a variance contribution of 32.212%, with Cr and Pb demonstrating the highest correlation with this component. This finding is consistent with the observed positive correlation between Cr and Pb (Fig. 8b), suggesting that Cr and Pb are the primary contributors to pollution within the first principal component. Fe and Cd exhibited the strongest correlation with the second principal component, which explained 23.301% of the variance. Mn showed the highest correlation with the third principal component, contributing 20.705% to the variance (Table 6 and Fig. 10). These contributions were markedly higher than those of the other principal components, indicating their dominant role in shaping the pollution profile.
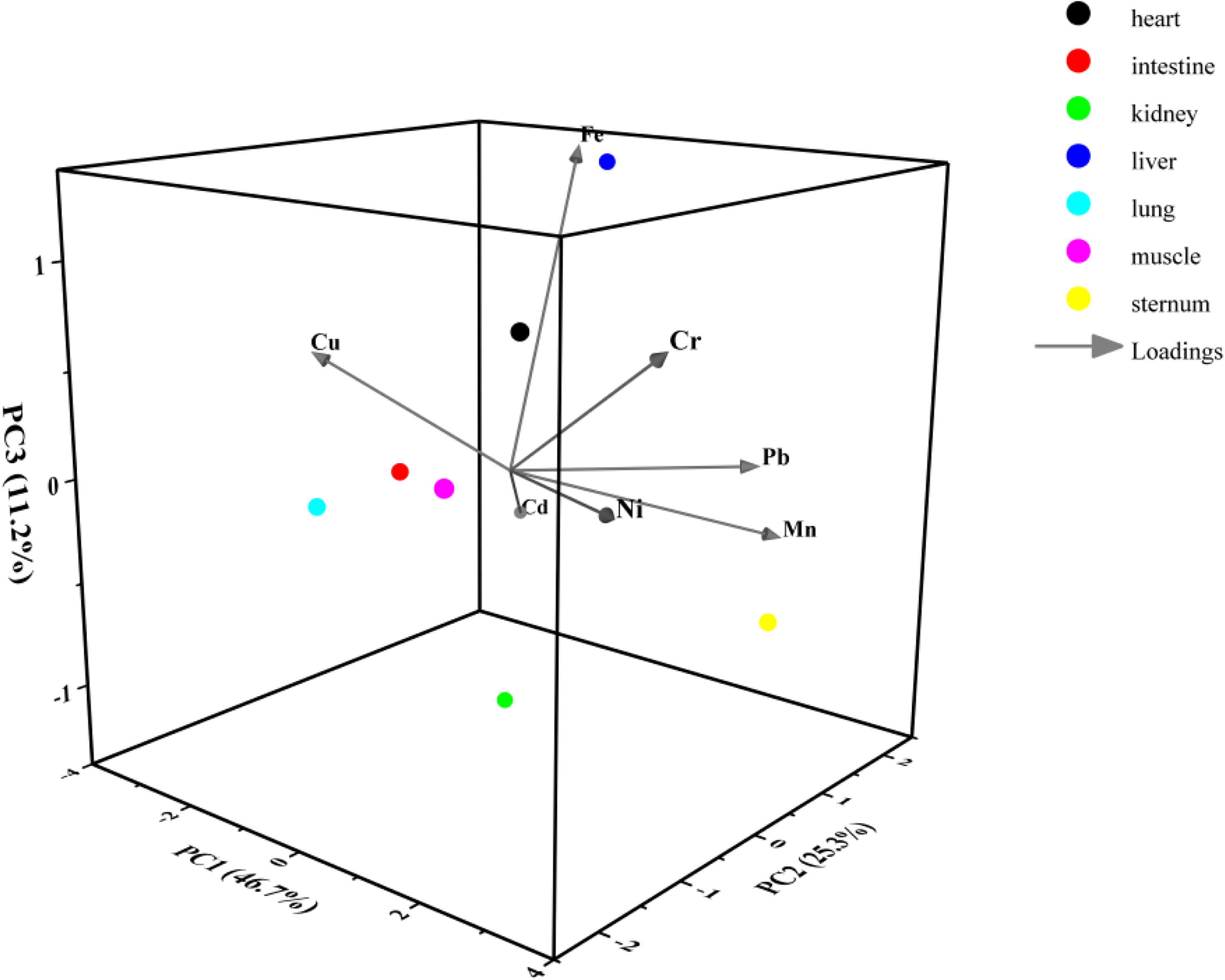
Metal factor loadings of the tissues of adult Saunders’s Gulls.
Analysis Results of PCA of Metals in Tissues of Adult Saunders’s Gulls
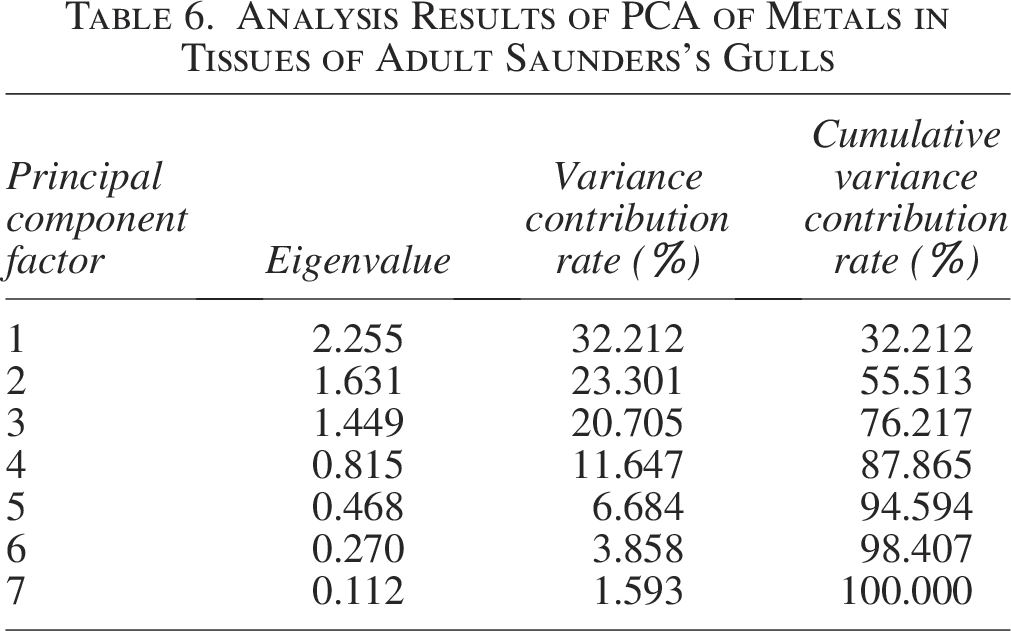
Furthermore, the first principal component showed relatively elevated loadings for Cr and Pb, while Cd displayed the highest loading on the second principal component, followed by Fe (Table 7). Mn likewise presented a relatively high loading on the third principal component.
PCA Load Value of Metals in Tissues of Adult Saunders’s Gulls
The principal component analysis (PCA) and loading results for Saunders’s Gulls have elucidated the metal pollution profiles within juvenile tissues, with Mn, Cu, and Fe being significantly associated with the first principal component, Ni and Pb with the second, and Cr with the third. The first principal component, which accounted for the highest variance, suggests that juvenile Saunders’s Gulls are predominantly influenced by Mn, Cu, and Fe pollution. Conversely, adult Saunders’s Gulls appear to be more affected by Cr and Pb pollution, with Fe, Cd, and Mn following as secondary contaminants.
Discussion
Metal levels in tissues of Saunders’s Gull across different age groups
Metal concentrations within identical tissues exhibit age-dependent variations in avian species. The investigation of Saeki et al. into Cd levels in tissues of cormorants (Phalacrocorax carbo) from Lake Biwa and the Tokyo region of Japan revealed that Cd concentrations in the kidneys and liver demonstrated a significant age-related increase (Saeki et al., 2000). For instance, in the White-throated Laughingthrush (Pycnonotus sinensis), concentrations of Cu, Pb, and Cd in the cardiac, hepatic, and pulmonary tissues of adult birds were markedly elevated compared with those in juveniles. Additionally, in Beijing, Cd, Pb, and Cu concentrations in adult sparrows were found to be consistently higher than those in nestlings (Chao et al., 2003). Burger further highlighted that Pb concentrations in the majority of adult tissues exceeded those found in the analogous tissues of juveniles (Burger, 1993). Our results align with the findings reported in preceding studies, underscoring the robustness of our observations.
The study on Saunders’s Gulls has uncovered variations in metal concentrations within identical tissues across different age cohorts. Specifically, Cr concentrations in the heart, sternum, muscle, kidney, and liver are notably elevated in juvenile birds when compared with their adult counterparts. Conversely, metal concentrations in certain other tissues are markedly higher in adult birds than in nestlings. These observations imply that metal concentrations in tissues are age-dependent, with an incremental accumulation of metals in various anatomical regions as the birds mature. Several plausible explanations may account for this observed phenomenon. On the one hand, the body necessitates specific concentrations of essential metal elements, including Cu, Mn, and Fe, to sustain normal physiological metabolism and growth. Consequently, adult individuals exhibit elevated metal concentrations, particularly Mn, which is crucial for skeletal ossification in homeothermic vertebrates. As the extent of bone ossification in birds intensifies, the Mn concentration in the sternum correspondingly augments. On the other hand, the excessive intake of nonessential elements, such as Cd and Pb, can result in their accumulation within tissues. Organisms possess detoxification mechanisms; in the case of birds, they can mitigate the harmful effects of certain metals and preserve the stability of their internal environment through liver detoxification, renal excretion, molting, and oviposition. However, the extent of metal accumulation is contingent upon the equilibrium between absorption and excretion processes. Under conditions of low environmental metal concentrations, avian excretion mechanisms are adept at sustaining lower metal concentrations within their bodies. Conversely, when environmental metal concentrations surpass a critical threshold, birds may be overwhelmed in their excretory capacity, resulting in substantial metal accumulation. Notably, as migratory species, Saunders’s Gulls are subject to variations in metal distribution within adult tissues, potentially influenced by the metal background levels in their wintering grounds or along their migratory pathways. Such variations may account for the observed discrepancies in metal distribution between nestling and adult bird tissues.
Elevated Ni concentrations in the hearts, sternums, muscles, kidneys, and livers of juvenile Saunders’s Gulls could potentially be ascribed to the transfer of metals from adult birds during birth, a mechanism that adult birds may employ to facilitate metal excretion. In contrast, the increased metal concentrations in adult Saunders’s Gulls relative to juveniles imply that adults may accumulate various metals more substantially during their migratory journeys. Notably, the fact that juvenile Saunders’s Gulls remain in the YRD postnatally underscores the potentially pivotal role of adult birds in amplifying metal accumulation throughout their migration.
Metal concentration in tissues of Saunders’s Gull across different sex
Sex influences metal concentration disparities within the tissues of organisms. Studies have demonstrated that in certain instances, male individuals exhibit higher accumulations of specific metals compared with their female counterparts (Hunter et al., 1987). For example, studies have revealed that male grasshoppers accumulate higher concentrations of Hg compared with their female counterparts. Similarly, in leaf-cutting bees (Gilpinia socia Klug) and European pine sawflies (Neodiprion sertifer Geoffroy), males have been observed to have elevated Ni concentrations relative to females (Heliövaara et al., 1990). The disparity in metal accumulation between male and female insects could be attributed to the dilution effect associated with body mass (Köhler and Triebskorn, 2013). It is commonly observed that females possess a greater body mass compared with males, which results in a more pronounced dilution effect and consequently lower metal concentrations within their tissues (Devkota and Schmidt, 2000).
In avian species, metal levels are modulated not only by hepatic detoxification and renal excretion but also through the process of feather molt. The regeneration of feathers demands a substantial investment of energy, with nutrients being supplied exclusively from the blood vessels that connect the feathers to the internal organs. This physiological process can lead to a reduction in metal levels within the bird. Furthermore, female birds possess the additional mechanism of excreting metals through the production of eggs. Research on American White Pelicans (Pelecanus erythrorhynchos) has indicated that, on average, male individuals exhibit higher metal concentrations in liver tissues than their female counterparts (Donaldson and Braune, 1999). In the Beijing region, male Eurasian Tree Sparrows (Passer montanus) have been observed to have elevated mercury concentrations compared with their female counterparts. This observed sex-based discrepancy in mercury levels is believed to be due to the oviposition process in females, which endows them with a distinct mechanism for metal excretion not present in males. Consequently, the physiological differences between the sexes lead to variations in their capacity to eliminate heavy metals from their systems (Pan et al., 2008). The concentration of metals in avian eggs is intricately linked to the metal levels within the reproductive systems of female birds, thereby serving as an indicator of the overall metal burden in the female’s body. This relationship underscores the capacity of female birds to eliminate certain metals through the process of egg-laying. However, it is important to note that research has also documented instances where females exhibit higher metal concentrations than males. For instance, the study of Gochfeld on metal concentrations in duck tissues revealed sex-based disparities, with female Greater Scaup (Aythya marila) displaying elevated levels of Pb and Mn compared with their male counterparts. These findings highlight the complexity of sex-based differences in metal accumulation and excretion among avian species (Gochfeld and Burger, 1987). This indicates that sex differences in metals could vary depending on the species.
In our examination of metal concentrations in Saunders’s Gulls, we observed sex-specific disparities within tissue metal levels. Males exhibited significantly or markedly higher concentrations of Fe in the sternum, Cr in the kidney, and Mn in both the lung and liver compared with females. Conversely, females showed significantly or markedly higher levels of Mn in the heart, Cd in the sternum, Cu in the muscle and lung, and Cd and Mn in the kidney, as well as Fe in the liver, compared with males. The tissue distribution patterns for Ni, Cu, and Mn were consistent between sex, with Ni primarily in muscle tissue and Cu and Mn predominantly in the liver. However, the distribution patterns for Cd, Cr, Pb, and Fe varied between sex. These findings suggest that sex may influence the distribution of certain metals. The differential metal concentrations between sex could be due to variations in tissue accumulation capacities, with females potentially utilizing reproduction to eliminate excess metals. At the same time, males and females may have different foraging behaviors, physiological characteristics, or migration patterns, which may also lead to differences in heavy metal accumulation. Since the birds in our study were nestlings and had not yet reached sexual maturity, the observed metal concentration differences are likely attributable to sex differences rather than reproductive processes. Further research on juvenile birds is warranted to elucidate the specific mechanisms underlying these disparities.
In summary, the distribution of metal concentrations within tissues is age-dependent, with an increase in metal enrichment observed as the birds age. The accumulation of metals in the environment is influenced by various factors, including sex and environmental conditions. While there are sex-based differences in metal concentrations within the same tissues, the distribution patterns across different tissues in male and female Saunders’s Gulls are not markedly distinct. This obscurity may stem from the fact that the sampled male and female birds were nestlings, with immature reproductive systems. To more accurately discern the metal distribution differences between sex, studies involving mature Saunders’s Gulls would be more suitable. In the future, we will focus on research in this area to fill the gap.
Comparison with toxicity thresholds and health risk implications
Currently, there is no unified limit standard for all bird species internationally. However, based on extensive ecological toxicology research, scientists have proposed some guideline thresholds, which are mainly used to assess whether wild birds are at potential risk of heavy metal exposure. These thresholds are typically targeted at the liver and kidneys, as they are the main organs for heavy metal accumulation.
In this study, the average concentrations of Pb in the kidneys of female and male Saunders’s Gull nestlings were 71.91 and 80.17 mg/kg, respectively, which were significantly higher than and far beyond the recognized threshold for kidney toxicity risk (20–30 mg/kg) (Franson and Pain, 2011); the average concentrations of Cu in the livers of female and male nestlings were 340.19 and 296.97 mg/kg, respectively, and the average concentrations of Pb were 90.21 and 70.47 mg/kg, respectively, all of which were higher than the recognized threshold for liver toxicity risk (Eisler, 2000; Franson and Pain, 2011; Scheuhammer, 1987); while the average contents of Mn in the livers of female and male nestlings were 17.76 and 28.44 mg/kg, respectively, although this value was higher than the background concentration, it was still far below the recognized threshold for liver toxicity risk (Eisler, 2000). This indicated that the current exposure levels of Cu and Pb in the Saunders’s Gull nestlings may already pose a direct risk of kidney and liver damage to the nestlings. The current exposure level of Mn may not yet cause direct liver damage to the individuals.
In female adult birds, the average concentration of Pb in the kidneys was 81.45 mg/kg, which was the same as that in the young birds and significantly higher than the recognized threshold for kidney toxicity risk (20–30 mg/kg) (Franson and Pain, 2011); the average concentrations of Cu and Pb in the liver were 503.93 and 99.94 mg/kg, respectively, far exceeding the recognized thresholds for liver toxicity risks of Cu and Pb (Eisler, 2000; Franson and Pain, 2011; Scheuhammer, 1987); while the average concentrations of Mn and Fe were 19.00 mg/kg and 2,969.77 mg/kg, respectively, although they were higher than the background concentrations, they were still far below the recognized thresholds for liver toxicity risks (Beyer and Meador, 2011; Eisler, 2000). This indicated that the current level of Pb exposure in adult female Saunders’s Gulls may pose a direct risk of damage to their kidneys; the levels of Cu and Pb exposure may pose a direct risk of damage to their livers. The levels of Mn and Fe exposure may not yet pose a direct risk of liver damage to adult female individuals.
Effects of excessive accumulation of metals on birds and the ability of different tissues of birds to accumulate metals
The accumulation of excessive metals can adversely impact multiple facets of avian physiology, encompassing hormonal regulation, reproductive capabilities, immune response, and overall health, thereby potentially jeopardizing their survival. Specifically, in a study conducted in a mining region of southwestern Finland, it was observed that exposure to metals such as Pb, Cd, Zn, Ni, and Cu resulted in a diminished hematocrit level in Great Tits (Parus major) and Pied Flycatchers (Ficedula hypoleuca). Concurrently, these birds exhibited reduced levels of alkaline phosphatase and total protein in their plasma, indicating a significant disruption in their physiological homeostasis (Espin et al., 2016). Metal contamination in Florida’s marshes has been associated with a reduction in estradiol levels among female White Ibises (Threskiornis melanocephalus) and an increase in testosterone levels among males. This hormonal imbalance is likely induced by the metals’ activation of the hypothalamic–pituitary–adrenal axis, which in turn may alter neurological, reproductive, and immune functions in these birds (Heath and Frederick, 2005). Blue Tits (Cyanistes caeruleus) living in heavily polluted areas experience reduced sperm count and motility and thinner eggshells (Dauwe et al., 2004).
The capacity for various bird tissues to accumulate specific metals can differ significantly. For example, mercury (Hg) exhibits a preference for accumulation in the lungs, hearts, and gizzards of piscivorous avian species, which may be attributed to their diet and the bioaccumulation processes within these organs (Agyemang, 2013).The sequestration of metal elements within the tissues of the ring-necked pheasant (Phasianus colchicus) exhibits tissue-specific patterns. Cd and Mn are predominantly found in bone and liver tissues, while Zn is chiefly accumulated in bone. Cu is primarily concentrated in the liver. Notably, elevated levels of Pb and Cd are observed in testicular tissue. In the context of Saunders’s Gulls, the liver, kidney, and muscle tissues are the primary sites for metal accumulation, with the highest concentrations of these elements. This distribution is closely linked to the physiological roles of these organs in metal metabolism and detoxification processes.
Comparing the elemental concentrations in the tissues of Saunders’s Gulls reveals that the liver accumulates relatively high levels of Cd, Cu, Pb, Mn, and Fe, making it the primary organ for metal accumulation. Among these elements, Cu, Mn, and Fe are essential for the organism’s health. Cu acts as an activator in hemoglobin synthesis, contributing to hematopoietic functions; Mn is a crucial component of various enzymes; and Fe is a vital part of hemoglobin, involved in oxygen transport and storage. The accumulation of these elements in the liver plays a positive role in the biochemical metabolism of birds. However, excessive levels of metals can also have toxic effects. Studies have reported that Se can induce adverse biological effects in the metabolism of glutathione and oxidative stress in mallards, leading to slowed growth and weakened immune function in ducklings (Hoffman, 2002). Furthermore, Cd and Pb are metal elements that pose serious hazards to animal organisms, and their accumulation in the liver is consistent with the liver’s detoxification function.
The accumulation of Cd and Fe in the kidneys is relatively high, with Cd exhibiting the highest concentration. Cd, when it enters the body, can induce excessive free radical production through various pathways. It has the ability to displace metals such as Fe and Cu in antioxidant enzymes, such as glutathione and peroxidase, significantly reducing the activity of these antioxidant enzymes within the body. Consequently, this weakens the body’s ability to clear free radicals (Wu et al., 2016). Cd also forms covalent bonds with thiol groups in proteins, such as cysteine residues, disrupting the thiol homeostasis within cells. This disruption can deplete reduced glutathione, inhibit antioxidant enzyme activity, and induce lipid peroxidation reactions (Bharavi et al., 2011). Additionally, a study has found that different concentrations of Cd lead to lipid peroxidation damage in kidney tissues, and this damage exhibits a dose-dependent relationship (Dabas et al., 2014).
The muscle tissue of Saunders’s Gulls accumulates relatively high levels of Ni and Cr. The toxicity of Cr is closely related to its valence state, with hexavalent Cr being significantly more toxic than trivalent Cr. Moreover, hexavalent Cr is readily absorbed by the body and accumulates within it. Due to its strong oxidative properties, hexavalent Cr readily penetrates biological membranes and exerts its toxic effects (Zayed and Terry, 2003).In this study, the highest chromium (Cr) concentration was detected in the muscle tissue, suggesting that muscle may be the primary site for Cr accumulation. It is likely that the absorbed Cr is primarily excreted through the kidneys.
The correlation analysis revealed significant positive correlations among Cu, Mn, and Fe, suggesting that these metals may share a common pollution source. Moreover, Pb and Cr pollution also appear to influence these metals. This pattern could be attributed to the fact that juvenile birds, from the moment of birth, feed and reside in the YRD area, where their tissue metal concentrations are shaped by the local metal environment. A study examining metal concentrations in feather and eggshell samples of C. boyciana and L. saundersi—two representative wetland bird species in the YRD region—demonstrated that these birds are predominantly contaminated by Cd, followed by Cr, with Mn being the least prevalent (Yang et al., 2020). Saunders’s Gulls, C. boyciana, and L. saundersi all reside in the YRD, yet they exhibit varying levels of metal pollution exposure. Such discrepancies may stem from the distinct bioaccumulation capacities of these species or from their divergent dietary habits.
Our findings also align with previous studies highlighting significant regional disparities in metal distribution within avian tissues. Notably, a circumpolar survey examining metal concentrations in willow ptarmigan livers and kidneys has disclosed substantial variations in Cd levels between Canada and Scandinavia (Pedersen et al., 2006). The observed differences in metal concentrations among Saunders’s Gulls are likely attributable to the varying degrees of environmental metal pollution in their respective habitats. Our study indicates that juvenile and adult Saunders’s Gulls are exposed to distinct primary metal pollutants, potentially a consequence of their migration patterns and the environmental conditions of their wintering grounds. This divergence in metal exposure could account for the observed variations in metal concentrations within their tissues. Juvenile birds, which are resident in the YRD breeding ground year-round, are likely predominantly affected by local metal pollution. PCA and associated loading values for adult Saunders’s Gulls suggest that they are primarily exposed to Cr and Pb pollution.
Conclusions
Variations in metal concentrations within avian tissues can be attributed to factors such as tissue type, age, sex, and environmental influences. Avian tissues demonstrate differential capacities for metal accumulation, with the liver, kidneys, and muscles in Saunders’s Gulls accumulating the highest levels of metal elements, a pattern that is closely associated with their physiological roles. Age-related differences in metal concentrations have been observed across various tissues, with adults typically exhibiting higher concentrations than nestlings, particularly for certain metals in specific tissues. Sex also plays a role in the tissue distribution of metal concentrations. Correlation and PCAs indicate that juvenile Saunders’s Gulls are predominantly affected by pollution from Mn, Cu, and Fe, which show a positive correlation among themselves. In contrast, metal pollution in adult Saunders’s Gulls is primarily attributed to Cr and Pb, exhibiting a highly significant correlation, followed by pollution from Cd, Fe, and Mn.
Authors’ Contributions
Y.Y.: Conceptualization, data curation, and writing—review and editing. X.Z.: Conceptualization, methodology, data curation, software, and writing—original draft. F.S.: Visualization and data curation. S.W.: Methodology and data curation. J.C.: Conceptualization, data curation, and writing—review and editing. M.T.: Methodology and writing—review and editing.
Footnotes
Author Disclosure Statement
The authors declare no competing interests.
Funding Information
The authors gratefully thank the financial support provided by the National Natural Science Foundation of China (31672314).
Data and Materials Availability
The authors declare that the data and materials supporting the findings of this study are available within the article.
Ethics Approval
The authors followed all the valid national rules for the use and care of animals, and the study was conducted after the ethical committee approval of the Department of Biomedical ethics approval document of Qufu Normal University.