
Abstract
During motor imagery, global inhibition and effector-specific inhibition contribute to prevent actual movements. We investigated the decay of inhibition using an action-mode switching paradigm. Participants switched between imagined and executed hand movements. Response–stimulus intervals (RSIs) were varied (200, 700, 1,300, and 2,000 ms). As inhibition (due to imagination) or activation (due to execution) in one trial affects performance in the subsequent trial, we analysed sequential effects. Evidence for the contribution of global inhibition (e.g., switch benefits in execution [E]—imagination [I] sequences compared with I-I sequences) and effector-specific inhibition (e.g., hand repetition costs after an imagination trial) was observed. Sequential effects decreased with increasing RSIs, indicating that both forms of inhibition are subject to decay. However, the decrease of sequential effects was less pronounced for global inhibition than for effector-specific inhibition. This indicates that global inhibition may decay slowly, whereas effector-specific inhibition decays rather quickly. In conclusion, global inhibition may be at least partly implemented in all contexts in which motor imagery has to be performed, whereas effector-specific inhibition may contribute to motor imagery only as soon as the exact movement parameters are known and may decay quickly after the imagined movement has been performed.
Introduction
Motor imagery refers to the mental simulation of a movement without actually executing it (Decety, 1996; Jeannerod, 1994, 1995). Motor imagery and motor execution have similar underlying processes and it is sometimes assumed that they are functionally equivalent (Jeannerod, 1995). Accordingly, motor imagery and motor execution overlap in neuronal activity (Munzert et al., 2009), motor imagery and motor execution often take the same amount of time (Guillot & Collet, 2005), and movement constraints are incorporated into motor imagery (Dahm & Rieger, 2016b; Papaxanthis et al., 2003). Nevertheless, differences between motor imagery and motor execution have also been observed (e.g., in brain activation, Guillot, Di Rienzo, et al., 2012 and movement durations, Guillot & Collet, 2005). Most notably, one apparent difference between motor imagery and motor execution is the necessity to prevent (i.e., inhibit) actual movements during motor imagery. However, inhibitory processes have received little attention in the motor imagery research. Only recently, research on inhibition in motor imagery has increased (cf. Bart et al., 2020; O’Shea & Moran, 2017, 2018; Rieger et al., 2017). So far, it is known that several forms of inhibition contribute to motor imagery (Guillot, Di Rienzo et al., 2012; O’Shea & Moran, 2018), most importantly, global inhibition (the inhibition of all motor commands) and effector-specific inhibition (the inhibition of motor commands of the effector used during motor imagery) (Bart et al., 2020; Rieger et al., 2017). Inhibition seems to last for some time after imagination is complete and affects the performance of subsequently executed movements negatively (Bart et al., 2020; Rieger et al., 2017; Scheil & Liefooghe, 2018). Most theories of cognitive architecture assume that activation of representations is a transient state that follows a time course (e.g., Anderson & Lebiere, 1998), and this temporal decay may also occur in the case of inhibition. However, in motor imagery the time course of inhibition has hardly been examined. Yet, different decay rates for global and effector-specific inhibition would further support the dissociation of the mechanisms underlying these forms of inhibition in motor imagery. Therefore, in the present study, we investigated the time course of decay of global inhibition and effector-specific inhibition in motor imagery.
The absence of actual movements during motor imagery can be explained in different ways (Jeannerod, 2001). As people know in advance that they have to imagine and not execute a movement during motor imagery, only subthreshold motor activation might be generated (Jeannerod & Frak, 1999). According to this view, motor imagery is just a weaker form of motor execution and the necessity of inhibition does not arise (Guillot, Di Rienzo et al., 2012; Jeannerod, 2001). However, recent studies provide evidence against this assumption and suggest that different forms of inhibition prevent actual movements in motor imagery (Bart et al., 2020; O’Shea & Moran, 2018; Rieger et al., 2017; Scheil & Liefooghe, 2018). Global inhibition and selective effector-specific inhibition seem to be the most important forms of inhibition (Bart et al., 2020; Rieger et al., 2017).
So far, studies indicate that inhibition of movements in motor imagery takes place early during movement preparation (Solomon et al., 2019). Inhibition continues to be present at least shortly after the imagination of a movement is complete (Bart et al., 2020; Rieger et al., 2017; Scheil & Liefooghe, 2018). However, to the best of our knowledge, nothing is known about how long inhibition continues to be present after imagination is complete and whether different forms of inhibition have a different time course of decay. We therefore investigated this in the present study.
One paradigm to investigate inhibition in motor imagery is the action mode switching paradigm as introduced by Rieger et al. (2017). The action mode switching paradigm is based on the experimental logic of task switching (for reviews see, for example, Hsieh, 2012; Kiesel et al., 2010; Koch et al., 2018; Monsell, 2003). Instead of switching between different tasks, participants switch between different action modes (imagination [I] and execution [E]), which are cued on a trial-to-trial basis. Inhibition during motor imagery should increase activation thresholds and therefore affect performance negatively in subsequently executed or imagined movements. Accordingly, the analysis of trial sequence effects, that is, reaction times (RTs) and movement times (MTs) in trial N depending on the action mode (I vs. E) in trial N-1, allows conclusions about the contribution of inhibition to motor imagery (Rieger et al., 2017). Different features of the task may be manipulated to investigate the different forms of inhibition. For instance, the comparison of action mode switches with action mode repetitions enables conclusions about global inhibition. By performing the task with different effectors, effector repetitions and effector alternations may be compared, enabling conclusions about effector-specific inhibition.
In the present study, we used the action mode switching paradigm to investigate the time course of decay of global inhibition and effector-specific inhibition in motor imagery. Participants executed and imagined hand movements from two home buttons (one for each hand, located on the left- and right-hand sides) to two vertically aligned target buttons (located in the middle between the home buttons). To investigate decay of inhibition the interval between the response in one trial and the stimulus of the next trial (response–stimulus interval [RSI]) was varied. Four different RSIs (200, 700, 1,300, and 2,000 ms) were used (in contrast to Rieger et al., 2017, who used a stable RSI of 500 ms). Participants performed the task in two sessions. In one session, RSIs were varied within experimental blocks, in the other session RSIs were varied between different experimental blocks.
We generally expected that imagination takes longer than execution, because this is frequently observed in short-lasting movements (i.e., movements lasting less than a few seconds, Guillot & Collet, 2005; Guillot, Hoyek, et al., 2012). Also, with regard to the different forms of inhibition contributing to motor imagery, we expected to observe similar data patterns as in previous studies using the action mode switching paradigm (Bart et al., 2020; Rieger et al., 2017; Scheil & Liefooghe, 2018), particularly at short RSIs (i.e., 200 ms, 700 ms), as those correspond most to the 500 ms RSI used by Rieger et al. (2017).
Global inhibition should become evident in the analysis of switch effects, that is, the comparison of action mode switches to action mode repetitions (cf. Rieger et al., 2017). Because global inhibition should take place in the previous trial in I-I sequences, we expected switch benefits in E-I sequences compared with I-I sequences. Further, global inhibition should take place in the previous trial in I-E sequences, and thus interfere with the execution of a movement in the subsequent trial (Bart et al., 2020; Rieger et al., 2017; Scheil & Liefooghe, 2018). Therefore, we expected switch costs in I-E sequences compared with E-E sequences.
Effector-specific inhibition should become evident in the data patterns within the different action mode sequences in which at least one of the movements is imagined. When movements are executed, activation of the respective effector (Adam & Koch, 2014; Bertelson, 1965; Soetens et al., 1985) should occur. Accordingly, increased activation due to the previously executed trial should facilitate performance in the subsequent trial (Adam & Koch, 2014). Thus, in E-E sequences we expected hand repetition benefits (shorter durations in movements with hand repetition than in movements with hand alternation, Bertelson, 1965). In contrast, when movements are imagined, inhibition of motor commands should prevent actual movements. Accordingly, in trial sequences in which at least one of the trials is an imagination trial, trial sequence effects should be different from E-E sequences (cf. Bart et al., 2020; Rieger et al., 2017). Because inhibition of the effector in the previous imagination trial should interfere with execution or imagination of the same effector in the subsequent trial (Bart et al., 2020; Rieger et al., 2017), we expected hand repetition costs in I-I and I-E sequences. Further, executing a movement results in an activation of this effector (Kirby, 1976; Soetens, 1998), thus more inhibitory effort should be required when a movement with the same effector is subsequently imagined. Accordingly, we expected hand repetition costs in E-I sequences.
The novel predictions refer to the time course of inhibition in motor imagery. We predicted that the expected sequential effects are less pronounced with longer RSIs (1,300 and 2,000 ms), as dissipation of sequential effects has been observed in various task contexts (Allport et al., 1994; Bertelson & Renkin, 1966; Egner et al., 2010; Koch, 2001; Meiran et al., 2000; Neill et al., 1992; Neill & Valdes, 1992; Wühr & Ansorge, 2005; but see also Horoufchin et al., 2011). Accordingly, we expected to observe evidence for decay of activation in sequences in which the previous trial is an execution trial (E-E sequences and E-I sequences). Furthermore, we expected that the different forms of inhibition in motor imagery are also subject to decay. Decay of global inhibition should become evident in less pronounced switch effects with increasing RSI. Decay of effector-specific inhibition should become evident in less pronounced hand repetition effects with increasing RSI in sequences, in which the previous trial is an imagination trial (i.e., I-I sequences and I-E sequences).
Although we expected that both forms of inhibition are subject to decay, the time course of decay may still differ between the different forms of inhibition. Global inhibition is brought about by two different processes. Global inhibition is partly event-based, that is, it is formed on a trial-to-trial basis and thus depends on the immediately preceding trial. For instance, switch benefits in E-I sequences compared with I-I sequences indicate that global inhibition takes place in the preceding imagination trial in I-I sequences (cf. Bart et al., 2020; Rieger et al., 2017). In addition, global inhibition is partly context-based, that is, it is higher in contexts in which motor imagery has to be performed more often. For instance, Rieger et al. (2017) observed longer durations in pure imagination blocks, consisting of imagination trials only, compared with mixed blocks, consisting of execution and imagination trials which occurred randomly (mixing benefits). This indicates that global inhibition may have been higher in pure imagination blocks. Thus, participants’ expectations about the likelihood of the occurrence of an imagination trial seem to contribute to global inhibition (Bart et al., 2020; Rieger et al, 2017). Accordingly, as global inhibition does not only depend on the immediately preceding trial but also on the task context, we speculate that global inhibition decays slowly. In comparison, effector-specific inhibition is closely related to the movement requirements in a specific trial, that is, which hand movement towards which target has to be performed. Thus, one may assume that after imagination is complete, effector-specific inhibition may decay quickly.
Participants performed the task in two sessions. In one session, RSIs were varied within blocks and in the other session RSIs were varied between blocks. It has been observed that the way RSIs are implemented influences whether or how much sequential effects dissipate with longer RSIs. For instance, it has been observed that switch costs in task switching decrease when the interval between the response of the last trial and the cue of the subsequent trial is varied within participants, but not when it is varied between participants (Altmann, 2005; Koch, 2001, 2005 see also Horoufchin et al., 2011, for a discussion). Similarly, effects of negative priming decrease with increasing RSIs when RSIs are varied within blocks (Neill et al., 1992; Neill & Valdes, 1992; Neill & Westberry, 1987; but see Hasher et al., 1996) but remain relatively stable even with long RSIs if RSIs are kept constant within blocks (Tipper et al., 1991). Thus, we expected that decay of inhibition in motor imagery may be more pronounced when RSIs are varied within blocks.
Method
Participants
22 psychology students (age in years: M = 22.7, SD = 5.7; sex: 11 males, 11 females; handedness: 19 right-handed, 3 left-handed as assessed by the Edinburgh Handedness Inventory, Oldfield, 1971) took part in the study. Participants received course credit for their participation. Originally, 24 students participated in the experiment. The data of two participants had mean durations of RTs or MTs more than three standard deviations above the mean of the other participants in several conditions. They were therefore excluded from analysis.
1
The number of participants was predetermined and based on past research (Bart et al., 2020; Rieger et al., 2017). To confirm the appropriateness of our sample size, we used G*Power 3.1.9.2 (Faul et al., 2007) to calculate the required sample size for one of the three-way interactions of interest (RSI × current action mode × previous action mode). We choose F-test (ANOVA, repeated measures, within factors) as the type of test. The number of measurements was set at four and the number of groups was set at one.
2
Statistical significance was set at p < .05. An effect-size of
Material and apparatus
The experiment was conducted using the software Presentation (Neurobehavioral Systems, www.neurobs.com, version 18.1). Stimuli were presented on an HpCompaq dc5850 monitor (screen: 23 in., vertical refresh rate: 60 Hz, resolution: 1,920 × 1,080 pixels). The background colour of the screen was white. Stimuli consisted of two vertically displayed circles with a black outline (1 cm radius, 4 cm distance between the centres) and were presented in the centre of the screen. One of the circles was coloured white, the other was coloured yellow (RGB: 255, 255, 0) or red (RGB: 255, 0, 0) indicating the action mode (imagination, execution). The action mode-colour assignment was counterbalanced across participants. The position of the coloured circle indicated the target button. In the middle of the coloured circle a right or left hand was presented indicating the hand, which should be used to perform the movement. Four buttons (radius: 3 cm) were fixed on a wooden board (length: 80 cm, width: 50 cm). Two of the buttons were located on the left and right sides of the board and served as home buttons (one for each hand, distance between the centres: 50 cm, distance from the front edge of the table: 25 cm). In the middle between the home buttons two target buttons were located on the same vertical plane (distance between the centres: 20 cm, distance from the centre to the edge of the table: lower target button: 15 cm, higher target button: 35 cm). The full set of experimental stimuli can be accessed from https://osf.io/qtvs4
Procedure
At the beginning of the trial participants were asked to press both home buttons (start position). After an RSI (200, 700, 1,300, or 2,000 ms) the stimulus was presented. In both imagination and execution trials, participants were instructed to identify the hand and the target button and to release the home button with the corresponding hand. As soon as the participants released the home button, the stimulus disappeared. In execution trials, participants were asked to move their hand to the target button, to press and to release the target button, and to move back to the home button. In imagination trials, participants were asked to imagine those movements from a first-person perspective, but to refrain from any actual movements to the target button. In both imagination and execution trials, participants were asked to press the home button at the end of the movement, which also served as the beginning of the next trial. Participants were instructed to react and move quickly while maintaining a high level of accuracy and not to correct errors. The original instructions (in German) and their English translation can be accessed from https://osf.io/qtvs4
In a session, participants first performed three practice blocks. In the first practice block only execution trials (33 trials), in the second practice block only imagination trials (33 trials), and in the third practice block imagination and execution trials (65 trials) were presented. If participants had difficulty in performing the task, the practice blocks were repeated. Afterwards, 12 experimental blocks consisting of 129 trials each (both imagination and execution trials) were presented. The trial order was fixed within blocks, such that all possible transitions from trial N-1 to trial N occurred equally often. Trial orders were created by modifying random trial sequences (cf. Rieger et al., 2017). The trial order was different for each block. The order of experimental blocks was randomised for each participant. After each block, participants had the opportunity to take a break.
Each participant performed two sessions, which were at least 1 week apart. In one session, RSIs varied within experimental blocks. The order of RSIs was created by modifying random RSI sequences, such that RSIs were equally distributed over all trial transitions. The sequence of RSIs was fixed within blocks, but was different for each block. In the other session, RSIs varied randomly between experimental blocks with the restriction that each RSI was used three times. The order of sessions was counterbalanced across participants.
Data analysis
We analysed RTs and MTs in milliseconds. RT was defined as the duration from stimulus presentation to the release of the home button at the beginning of the movement. MT was defined as the duration from the release of the home button at the beginning of the movement to the press of the home button at the end of the movement.
In all, 3.55% of the trials were excluded from the analysis for the following reasons: (a) RTs or MTs were above or below 3 SDs of the mean times of a condition in a participant, (b) participants used the wrong hand, (c) participants performed the wrong action mode (execution instead of imagination or imagination instead of execution), or (d) participants did not adhere to the instructions (e.g., corrected an erroneous movement). Movement errors in execution (i.e., movements in which the wrong target button was pressed) were included in the analysis as those errors cannot be determined in imagination trials (cf. Rieger et al., 2017).
RTs and MTs were analysed using ANOVAs. Note that because our design involves six factors, we split the analysis into two theoretically meaningful ANOVA designs, involving only four factors each (for details see below). If Mauchly’s test indicated that the assumption of sphericity was violated (which concerns the four-level factor RSI and all interactions with this factor), we report Greenhouse–Geisser’s corrected degrees of freedom, p-values and Greenhouse–Geisser’s ε. Post hoc comparisons were conducted using paired t-tests. Significance values of post hoc tests were adjusted for multiple testing using Sidak correction. As a measure of effect size, we report Cohen’s dz for within-subjects post hoc comparisons (Lakens, 2013). When several post hoc comparisons are reported together, minimum (pmin, dz_min) or maximum statistical values (pmax, dz_max) are reported.
Results
Means of RTs can be seen in Figure 1. Means of MTs can be seen in Figure 2. Standard errors of the mean and 95% confidence intervals are provided in the supplementary material.
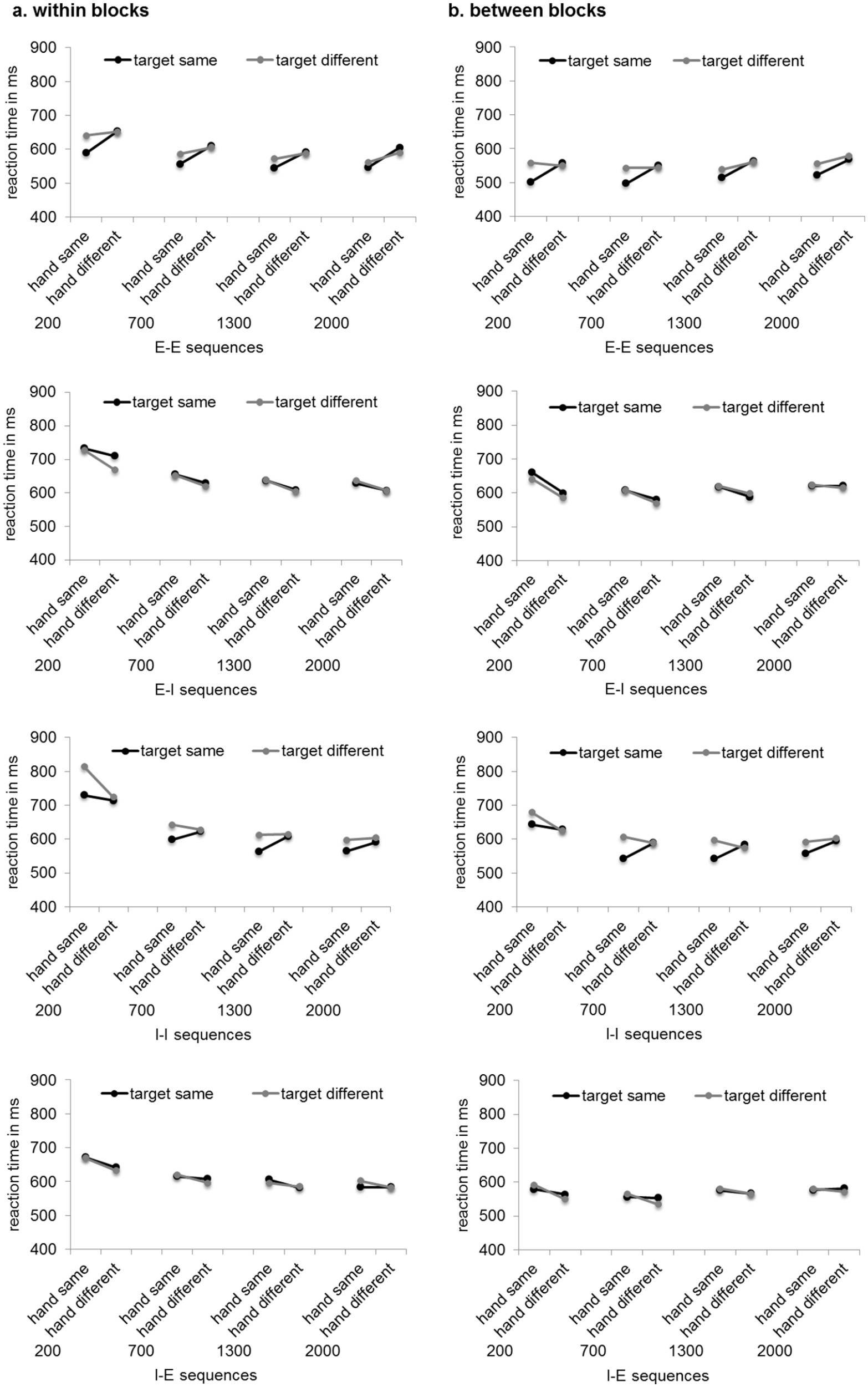
Means of reaction times in milliseconds depending on RSI implementation (panel a: within blocks, panel b: between blocks), RSI (200, 700, 1,300, 2,000 ms), hand (same, different), and target (same, different) for the different action mode sequences. Letters indicate the action mode sequence (I = imagination, E = execution) with the first letter referring to the action mode in trial N-1 and the second letter referring to the action mode in trial N.
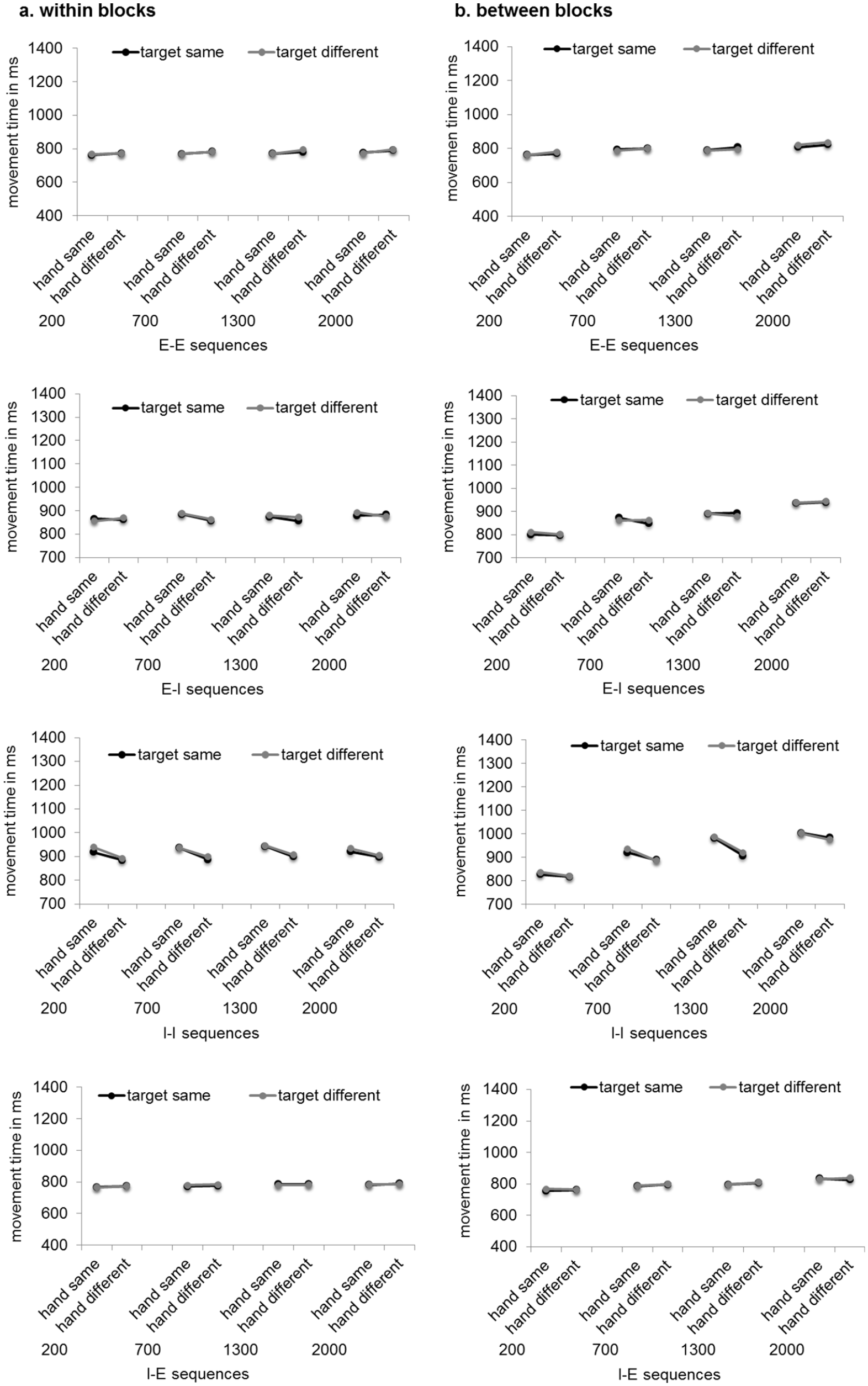
Means of movement times in milliseconds depending on RSI implementation (panel a: within blocks, panel b: between blocks), RSI (200, 700, 1,300, 2,000 ms), hand (same, different), and target (same, different) for the different action mode sequences. Letters indicate the action mode sequence (I = imagination, E = execution) with the first letter referring to the action mode in trial N-1 and the second letter referring to the action mode in trial N.
Analysis of data pooled over hand and target
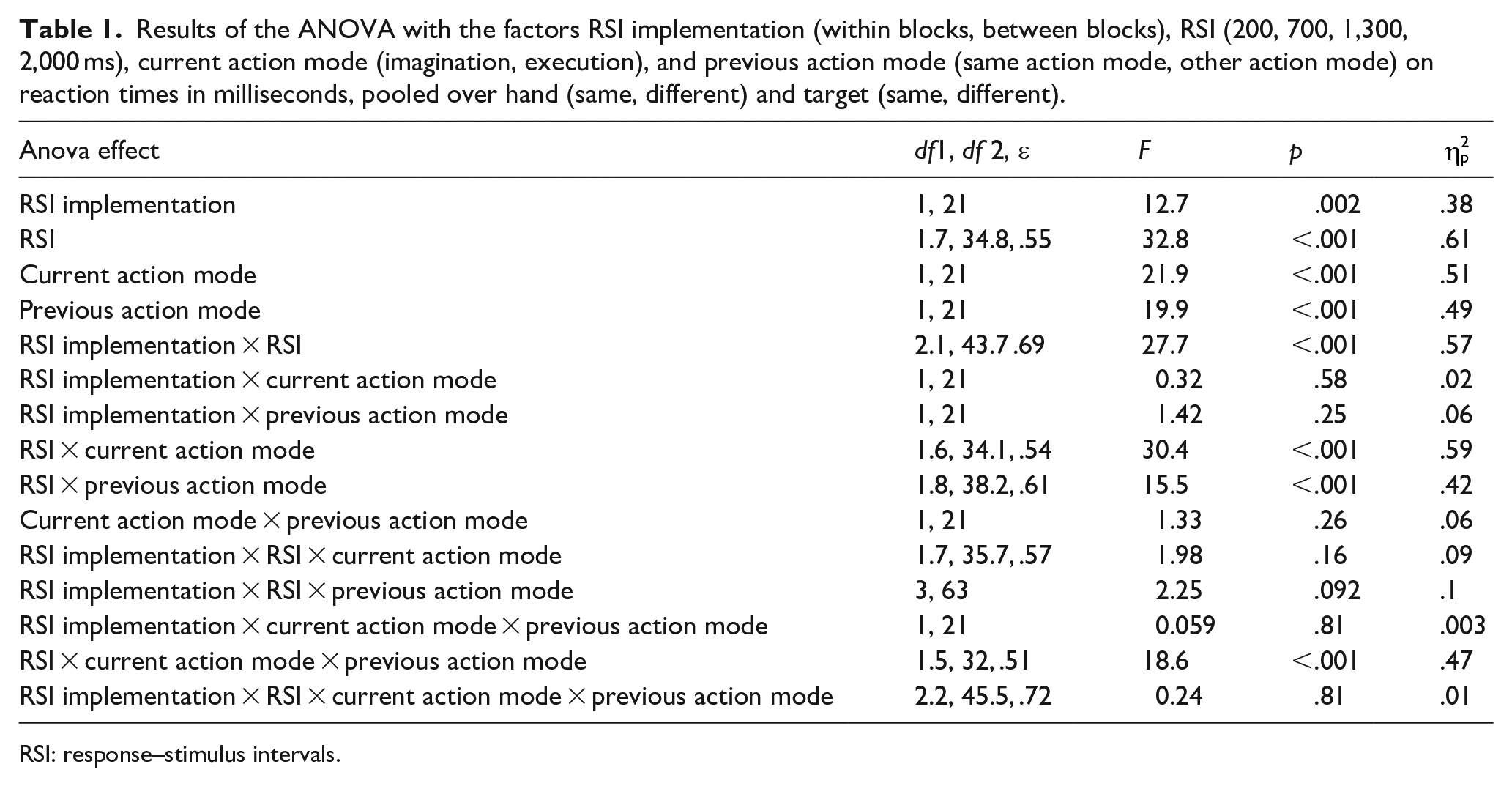
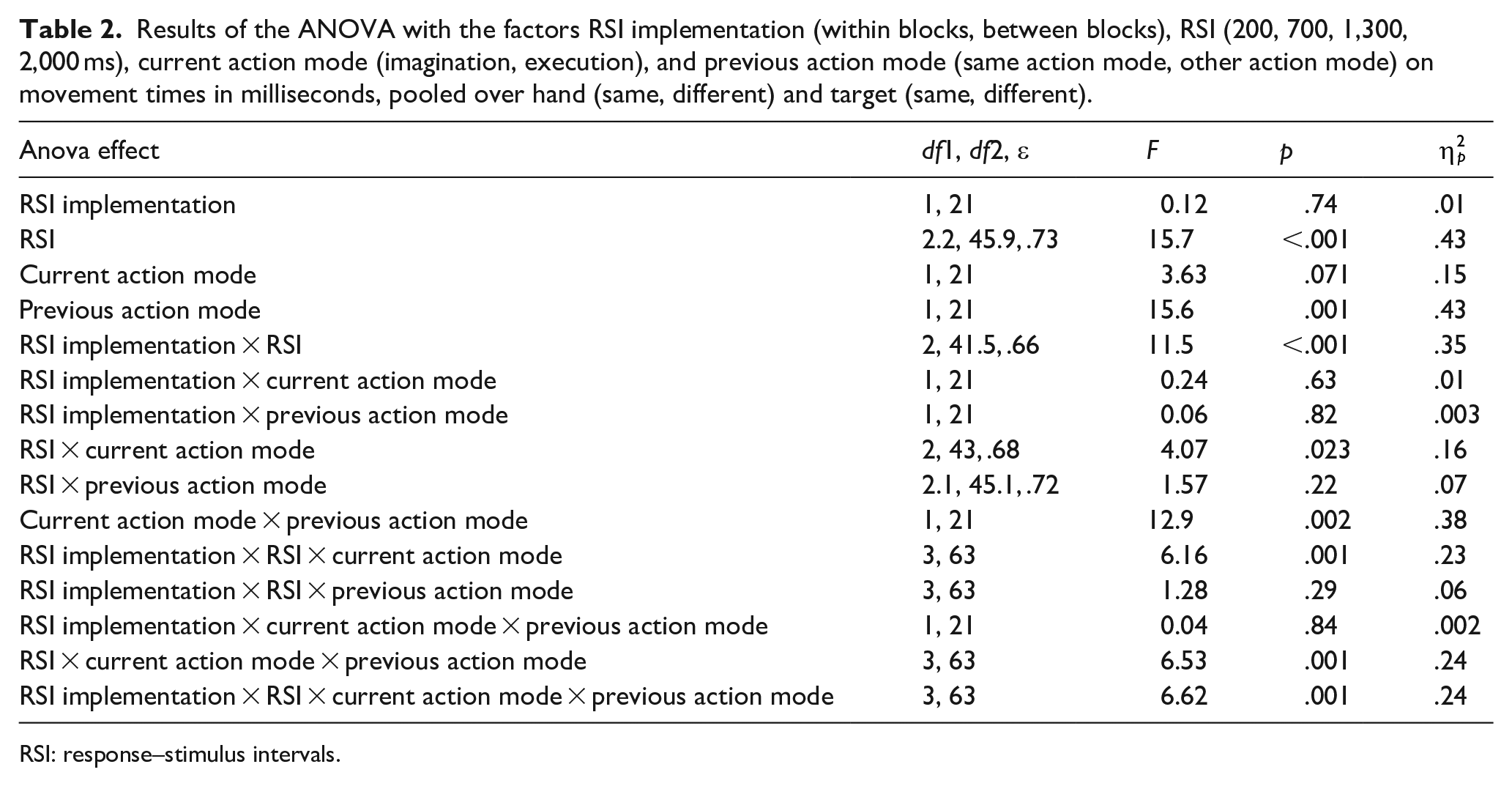
In a first step, we performed ANOVAs with the factors RSI implementation (within blocks, between blocks), RSI (200, 700, 1,300, 2,000 ms), current action mode (imagination, execution), and previous action mode (same action mode, other action mode). For this analysis, data were pooled over hand (same, different) and target (same, different). One interest in this analysis was to look at the overall differences between imagination and execution (current action mode) as well as overall effects of RSI implementation and RSI. A further interest was to look at switch costs/benefits (which result from specific combinations of previous and current action mode) as indicators for global inhibition and how switch effects interact with RSI implementation and RSI. Because previous data show that data from RTs and MTs are relevant for the effects attributed to global inhibition (cf. Bart et al., 2020; Rieger et al., 2017), we analysed both RTs and MTs separately. Results of the ANOVAs are depicted in Table 1 (RTs) and Table 2 (MTs). To increase readability, we do not mention the respective ANOVA effects in the text and ask the reader to refer to the tables.
Results of the ANOVA with the factors RSI implementation (within blocks, between blocks), RSI (200, 700, 1,300, 2,000 ms), current action mode (imagination, execution), and previous action mode (same action mode, other action mode) on reaction times in milliseconds, pooled over hand (same, different) and target (same, different).
RSI: response–stimulus intervals.
Results of the ANOVA with the factors RSI implementation (within blocks, between blocks), RSI (200, 700, 1,300, 2,000 ms), current action mode (imagination, execution), and previous action mode (same action mode, other action mode) on movement times in milliseconds, pooled over hand (same, different) and target (same, different).
RSI: response–stimulus intervals.
Current action mode
RTs were significantly longer in imagination (M = 623 ms, SE = 17 ms) than in execution (M = 579 ms, SE = 15 ms). In MTs, no significant difference between imagination and execution was observed.
RSI implementation and RSI
RTs were significantly longer when RSIs varied within blocks (M = 623 ms, SE = 17 ms) than when RSIs varied between blocks (M = 580 ms, SE = 16 ms). RTs were significantly longer at the 200 ms RSI than at the 700, 1,300, and 2,000 ms RSI (pmax < .001, dz_min = 1.2). All other comparisons were not significant (pmin = .85, dz_max = 0.24). RT differences between the 200 ms RSI and the other RSIs were significantly larger when RSIs varied within blocks than when RSIs varied between blocks (pmax = .006, dz_min = 0.74).
Results in MTs showed a different pattern, indicating a trade-off between RTs and MTs: MTs were significantly shorter at the 200 ms RSI than at the 1,300 and 2,000 ms RSI (pmax = .006, dz_min = 0.82). Further, MTs were significantly shorter at the 700 ms RSI than at the 2,000 ms RSI (p = .009, dz = 0.77). All other comparisons between RSIs were not significant (pmin = .09, dz_max = 0.56). In addition, MT differences between RSIs were significantly larger when RSIs varied between blocks than when RSIs varied within blocks (pmax = .018, dz_min = 0.65). 3
Taken together, RTs were significantly longer when RSIs varied within blocks than when RSIs varied between blocks. Furthermore, RTs decreased with increasing RSIs, whereas MTs showed a reverse effect.
Switch costs/benefits
Means and standard errors of switch costs/benefits depending on RSI implementation (within blocks, between blocks) and RSI (200, 700, 1300, 2000 ms) in RTs and MTs are depicted in Figure 3. Switch costs/benefits were calculated as E-I sequences minus I-I sequences when the current action mode was imagination and as I-E sequences minus E-E sequences when the current action mode was execution.
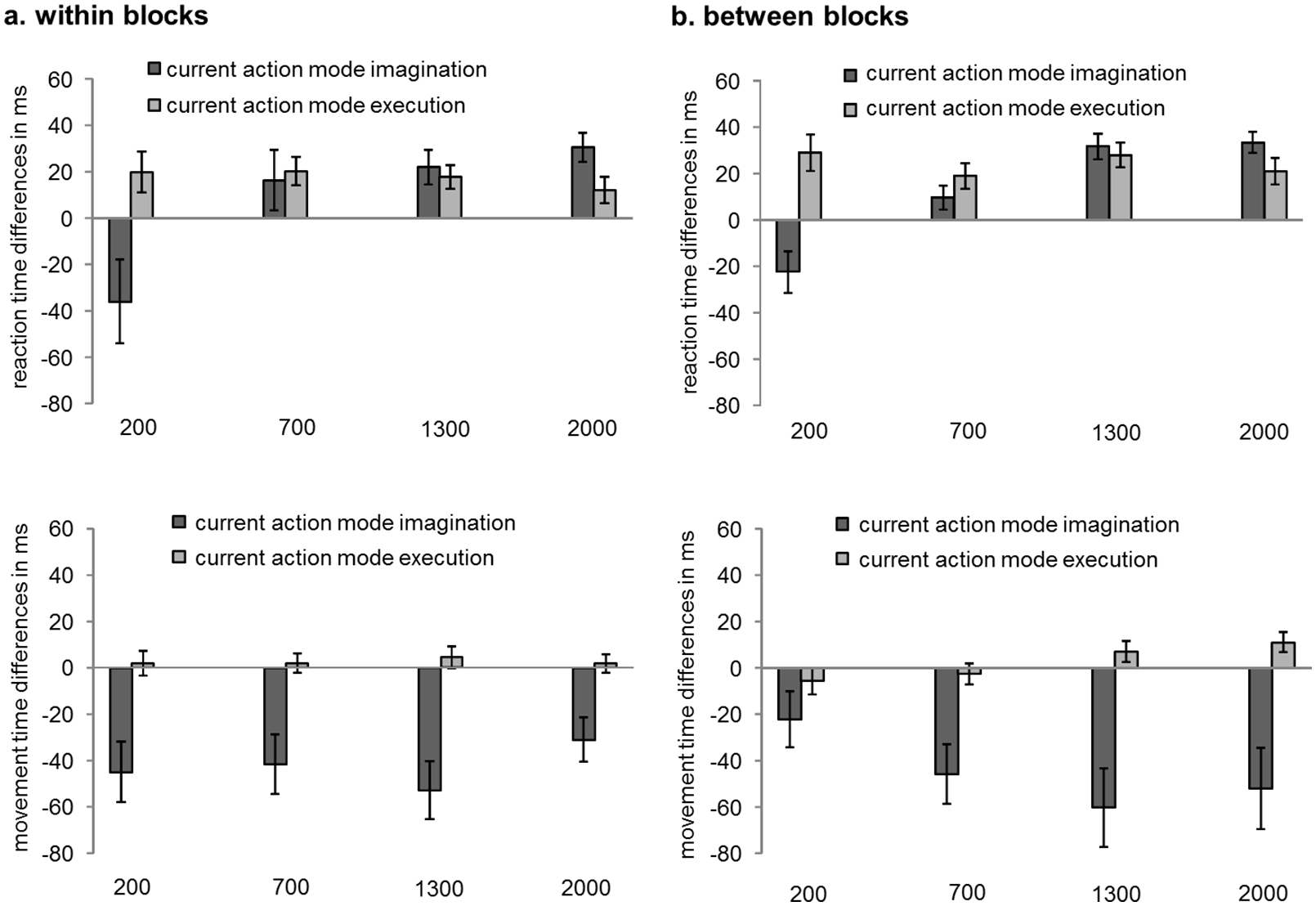
Means and standard errors of switch effects (positive values: costs; negative values: benefits) depending on RSI implementation (panel a: within blocks, panel b: between blocks) and RSI (200, 700, 1,300, 2,000 ms) in reaction times in milliseconds (upper panels) and movement times in milliseconds (lower panels). Switch effects were calculated as E-I sequences minus I-I sequences when the current action mode was imagination and as I-E sequences minus E-E sequences when the current action mode was execution.
When the current action mode was imagination, that is, in E-I sequences compared with I-I sequences, significant switch benefits were observed in RTs at the 200 ms RSI (p = .033, dz = 0.49). No significant switch effects occurred at the 700 ms RSI (p = .13, dz = 0.33) and significant switch costs were observed at the 1,300 and 2,000 ms RSIs (pmax < .001, dz_min = 1.1). Switch effects did not differ significantly depending on RSI implementation. In MTs, significant switch benefits occurred at all four RSIs (pmax = .007, dz_min = 0.64). When RSIs varied between blocks, switch benefits were significantly smaller at the 200 RSI than at the 1,300 ms RSI (p = .041, dz = 0.63). All other differences were not significant (pmin = .17, dz_max = 0.5). When RSIs varied within blocks, switch benefits were significantly larger at the 1,300 ms RSI than at the 2,000 ms RSI (p = .006, dz = 0.78). All other differences were not significant (pmin = .47, dz_max = 0.37). However, a direct comparison of the switch benefits between and within blocks showed that they did not significantly differ from each other, for none of the RSIs (pmin = .21, dz_max = 0.43). When we look at the combined effects of RT and MT, we observe significant switch benefits at all RSIs (pmax = .004, dz_min = 0.8), which do not differ significantly between RSIs (pmin = .11, dz_max = 0.54).
When the current action mode was execution, that is, in I-E sequences compared with E-E sequences, significant switch costs occurred in RTs at all four RSIs (pmax = .004, dz_min = 0.7). Switch costs did not significantly differ between RSIs (pmin = .15, dz_max = 0.51). Switch costs did not differ significantly depending on RSI implementation. In MTs, no significant switch effects were observed at all four RSIs (pmin = .071, dz_max = 0.41).
Taken together, we observed switch costs (when the current action mode was execution) and switch benefits (when the current action mode was imagination), which were relatively stable over time.
Analysis of data within the different action mode sequences
In a second step, we performed ANOVAs with the within-participants factors RSI implementation (within blocks, between blocks), RSI (200, 700, 1,300, 2,000 ms), hand (same, different), and target (same, different) separately for each action mode sequence (E-E, E-I, I-I, and I-E). In these analyses, the interactions of RSI implementation and RSI with hand repetition effects were of major interest, as hand repetition costs/benefits serve as indicators for effector-specific inhibition. Because previous data indicate that mainly RTs are relevant for the effects attributed to effector-specific inhibition (cf. Bart et al., 2020; Rieger et al., 2017), we only report the results on RTs in the text. Results on MTs can be seen in the supplementary material.
The results of the ANOVAs on RTs for each action mode sequence are depicted in Table 3. To increase readability, we do not mention the respective ANOVA effects in the text and ask the reader to refer to the table.
Results of the ANOVAs with the factors RSI implementation (within blocks, between blocks), RSI (200, 700, 1,300, 2,000 ms), hand (same, different), and target (same, different) on reaction times in milliseconds for each action mode sequence.
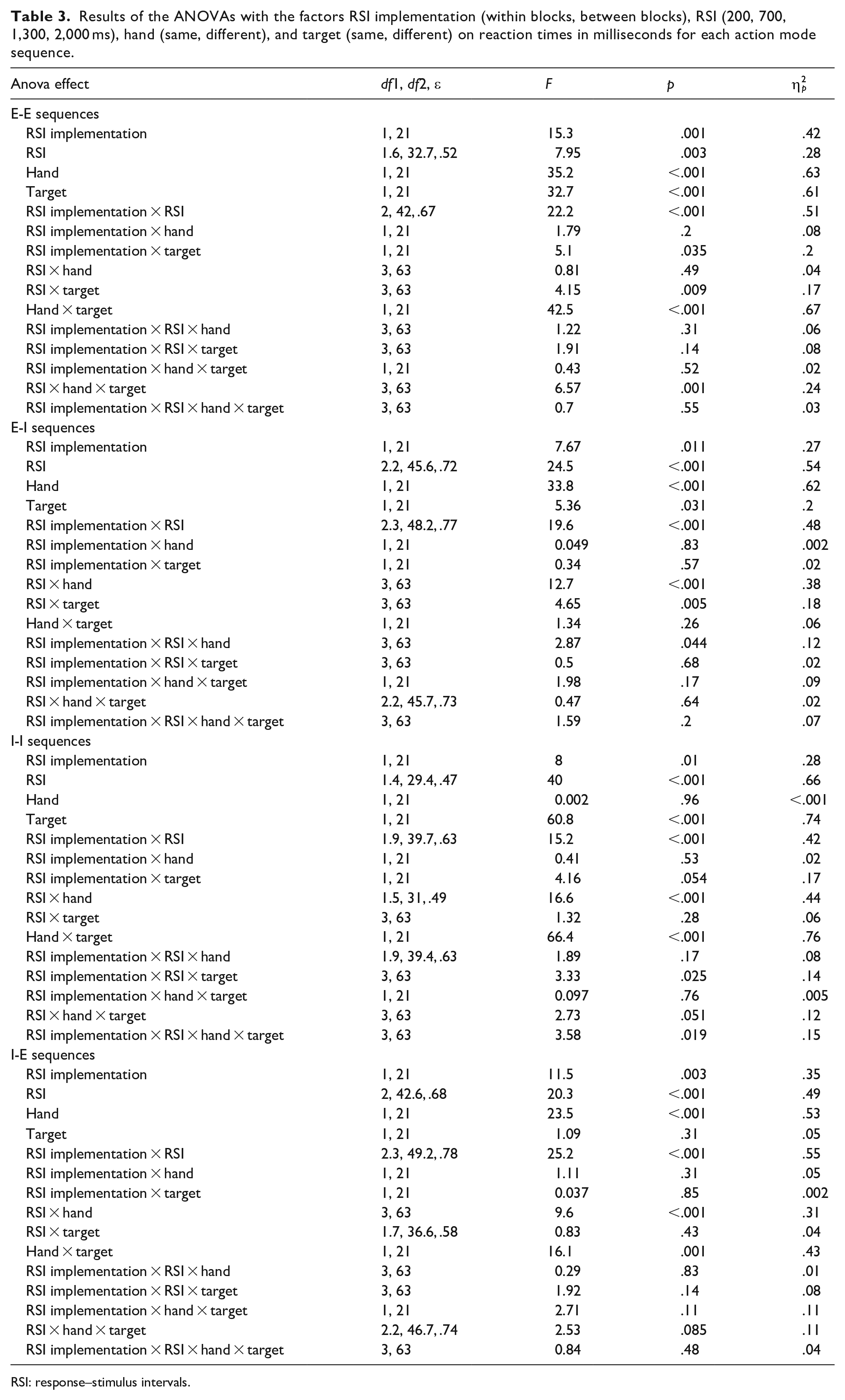
RSI: response–stimulus intervals.
Means and standard errors of hand repetition costs/benefits depending on RSI implementation (within blocks, between blocks) and RSI (200, 700, 1,300, 2,000 ms) in RTs are depicted in Figure 4. Hand repetition costs/benefits were calculated as complete repetition minus hand alternation with target repetition for trials with target repetition and as hand repetition with target alternation minus complete alternation for trials with target alternation.
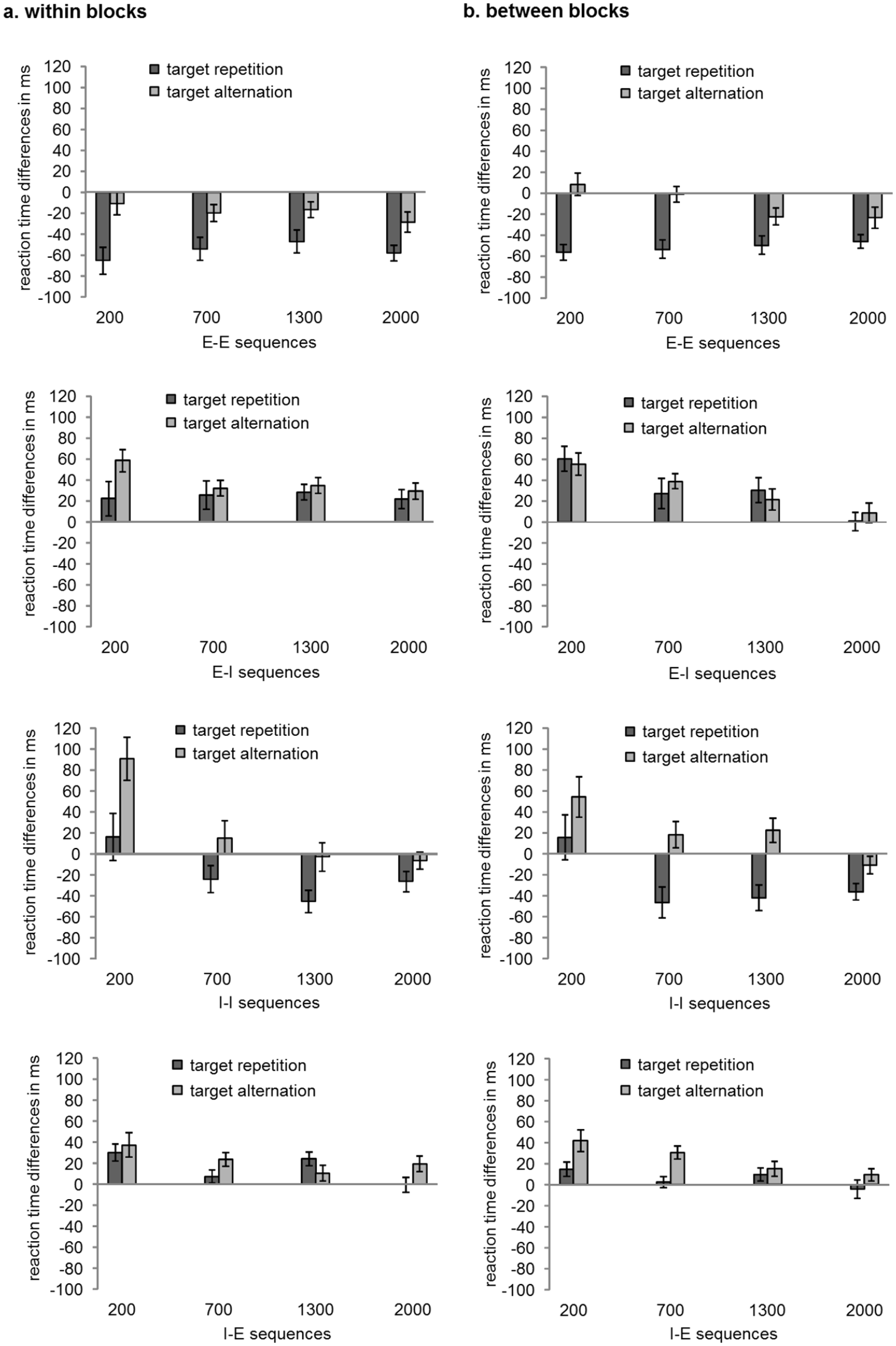
Means and standard errors of hand repetition effects (positive values: costs; negative values: benefits) depending on RSI implementation (panel a: within blocks, panel b: between blocks) and RSI (200, 700, 1,300, 2,000 ms) in reaction times in milliseconds for each action mode sequence. Hand repetition effects were calculated as complete repetition minus hand alternation with target repetition for trials with target repetition and as hand repetition with target alternation minus complete alternation for trials with target alternation. Letters denote the action mode sequence (I = imagination, E = execution). The first letter refers the action mode in trial N-1 and the second letter to the action mode in trial N.
E-E sequences
Significant hand repetition benefits occurred in movements with target repetition at all four RSIs (pmax < .001, dz_min = 1.2). Hand repetition benefits did not significantly differ between RSIs (pmin = .23, dz_max = 0.46). Further, significant hand repetition benefits occurred in movements with target alternation at the 1,300 and 2,000 ms RSI (pmax = .008, dz_min = 0.62), but no significant hand repetition effects were observed at the 200 and 700 ms RSI (pmin = .14, dz_max = 0.33). At the 1,300 and 2,000 ms RSI hand repetition benefits were significantly higher in movements with target repetition than in movements with target alternation (pmax = .018, dz_min = 0.61). Hand repetition effects did not differ significantly depending on RSI implementation.
Taken together, in E-E sequences hand repetition benefits occurred in movements with target repetition at short RSIs and appeared independent of the target at long RSIs. Further, hand repetition benefits did not decrease with increasing RSIs.
E-I sequences
When RSIs varied within blocks, hand repetition costs were significant at all four RSIs (pmax = .002, dz_min = 0.78) and did not differ significantly between RSIs (pmin = .71, dz_min = 0.29). When RSIs varied between blocks, significant hand repetition costs occurred at the 200, 700, and 1,300 ms RSIs (pmax = .007, dz_min = 0.64), but not at the 2,000 ms RSI (p = .44, dz = 0.17). Hand repetition costs were significantly higher at the 200 ms RSI than at the 700 and 1,300 ms RSIs (pmax = .012, dz_min = 0.68), the latter two RSIs did not differ significantly from each other (p = .8, dz_min = 0.18).
Taken together, in E-I sequences we observed hand repetition costs. Hand repetition costs decreased when RSIs varied between blocks (though the decrease was slow as costs were still observable at the 1,300 ms RSI), but not when RSIs varied within blocks.
I-I sequences
At the 200 ms RSI, significant hand repetition costs occurred in movements with target alternation (pmax = .01. dz_min = 0.61), but not in movements with target repetition (pmin = .47, dz_max = 0.16). At the 700 ms RSI, no significant hand repetition effects occurred in movements with target alternation (pmin = .15, dz_min = 0.32). In movements with target repetition, significant hand repetition benefits occurred when RSIs varied between blocks (p = .005, dz = 0.67), but no significant hand repetition effects were observed when RSIs varied within blocks (p = .072, dz = 0.4). However, a direct comparison of the hand repetition effects between and within blocks, showed that they did not significantly differ from each other (p = .14, dz = 0.33). At the 1,300 and 2,000 ms RSIs, significant hand repetition benefits occurred in movements with target repetition (pmax = .011, dz_min = 0.59), but not in movements with target alternation (pmin = .073, dz_max = 0.4).
Taken together, in I-I sequences we observed hand repetition costs in movements with target alternation (primarily at the shortest RSI), which decreased over time. Further, with increasing RSIs we observed hand repetition benefits in movement with target repetition.
I-E sequences
Significant hand repetition costs occurred at the 200, 700, and 1,300 ms RSIs (pmax = .001, dz_min = 0.89), but no significant hand repetition effects occurred at the 2,000 ms RSI (p = .2, dz = 0.28). Hand repetition costs were significantly higher at the 200 RSI than at the 700 and 1,300 ms RSIs (pmax = .036, dz_min = 0.59), the latter two did not differ significantly from each other (p = .98, dz = 0.07). Hand repetition effects did not differ significantly depending on RSI implementation. Taken together, in I-E sequences we observed hand repetition costs, which decreased with increasing RSI and eventually disappeared at the longest RSI.
Discussion
In the present experiment we investigated the decay of global inhibition and effector-specific inhibition in motor imagery using the action mode switching paradigm (Rieger et al., 2017). We asked participants to execute and imagine hand movements from two home buttons to two vertically arranged target buttons and analysed sequential effects. RSIs (200, 700, 1,300, 2,000 ms) were varied within blocks and between blocks.
Action mode
RTs were longer in imagination than in execution. In the present task, during the RT phase of an imagination trial, participants have to plan an actual movement (release of the start button) and an imagined movement (movement to the target button). This may be more time-consuming than movement planning of actual movements only (release of the start button and movement to the target button in an execution trial), resulting in overall longer RTs in imagination trials. In addition, alternating between activation of the effector (indicating start of the imagined movement) and inhibition of the effector (imagined movement to the target) in imagination trials may have further prolonged RTs (Dahm & Rieger, 2016a; Rieger et al., 2017). MTs did not significantly differ between imagination and execution, even though shorter imagination durations in MTs have been observed in previous studies using similar tasks (Dahm & Rieger, 2016a; Rieger et al., 2017). However, whereas it has been observed that short-lasting movements sometimes take longer in imagination than execution, this is not always the case (for a review see Guillot & Collet, 2005).
Overall effects of RSI implementation and RSI
RTs were longer when RSIs varied within blocks than when RSIs varied between blocks. When RSIs vary between blocks participants may anticipate at which time the next stimulus will appear (Grosjean et al., 2001). Such anticipations can result in an increase of phasic alertness, thereby enhancing response readiness which results in faster responses.
Further, RTs were longer at short RSIs than at long RSIs. This is a frequent finding in reaction time tasks (Rabbitt, 1969; Welford, 1952) and may be caused by limited attentional resources and/or still ongoing processing of the preceding trial, which slows response preparation (Maslovat et al., 2013) or response execution (Bratzke et al., 2009) in the subsequent trial.
In contrast to RTs, MTs were shorter at short RSIs than at long RSIs. Such an inverse relationship between RTs and MTs is sometimes observed (Danev et al., 1971; Howell, 1953; Wühr & Heuer, 2015). One explanation for this inverse relationship between RTs and MTs might be that at short RSIs, movements are planned more thoroughly during the longer RT phase, allowing shorter MTs, whereas part of movement planning is postponed into MTs at the other RSIs (even though they were instructed otherwise). However, it seems unlikely that participants changed their response criterion during the experiment in a way that in some trials movement planning was completed prior to responding, whereas in others it was postponed into MTs. Therefore, another explanation may be that participants tried to keep the overall speed of their response (i.e., from stimulus onset to the press of the home button at the end of the movement) constant over trials. Thus, when RTs were longer at short RSIs, participants may have tried to compensate for this by speeding up MTs and vice versa.
Note that the decay of both global inhibition and effector-specific inhibition (which will be discussed below) was hardly affected by whether RSIs varied between or within blocks. We had speculated that decay of inhibition might be less pronounced when RSIs are varied between blocks as similar results have been previously observed for sequential effects in task switching (Altmann, 2005) and negative priming (Tipper et al., 1991). However, this was not the case.
Time course of global inhibition
Note that even though data from RTs and MTs are both informative about the effects attributed to global inhibition, it has previously been observed that effects in RTs are sometimes less clear than effects in MTs. RTs are additionally affected by costs due to action mode switching (cf. Bart et al., 2020; Rieger et al., 2017). This also became apparent in the present study.
We observed shorter RTs (switch benefits) when the current action mode was imagination, that is, in E-I sequences compared with I-I sequences, at the 200 ms RSI. Switch benefits disappeared at the 700 ms RSI and switch costs occurred at the 1,300 and 2,000 ms RSIs. In MTs, switch benefits were relatively stable over all RSIs. We have no good explanation of why switch benefits decreased in RTs but were stable in MTs. When we looked at the combined effects of RT and MT we found switch benefits at all RSIs, and switch benefits did not significantly differ from each other. Thus, overall, the results indicate that in I-I sequences global inhibition took place during imagination of the movement in the previous trial, thus slowing down performance in the subsequent imagination trial (Bart et al., 2020; Rieger et al., 2017). Other effects, such as the requirement to switch between actions may partly modulate switch benefits, but it is not exactly clear how those mechanisms interact with each other.
When the current action mode was execution, that is, in I-E sequences compared with E-E sequences, we observed longer RTs (switch costs) at all four RSIs. Two processes may contribute to this effect: switch costs may occur because global inhibition took place in imagination, which interfered with executing the movement in the subsequent trial (Bart et al., 2020; Rieger et al., 2017; Scheil & Liefooghe, 2018). Further, the requirement to switch between actions may have contributed to this effect as well. As no significant switch effects were observed in MTs, it could be that switch costs in RTs were mainly caused by the latter process when the current action mode was execution.
Time course of effector activation
Data patterns of action mode sequences, in which the previous trial was an execution trial (i.e., E-E sequences and E-I sequences), provide evidence for effector activation. In E-E sequences, hand repetition benefits occurred. Such repetition benefits are commonly observed when movements are repeated and indicate that activation of the effector (Adam & Koch, 2014; Bertelson, 1965; Soetens et al., 1985) in the previous trial facilitated performance in the subsequent trial.
At the 200 and 700 ms RSIs, hand repetition benefits occurred only in complete repetitions (i.e., movements with hand and target repetition). With increasing RSIs hand repetition benefits appeared independent of the target. Those results may indicate that different task features like the used effector and the movement towards a certain target may become bound to an integrated “event file” (Hommel, 1998; see also Frings et al., 2020), such that partial repetitions (i.e., only the effector) do not facilitate the response, at least at short RSIs. Over time, the previously integrated task features may unbind (Stoet & Hommel, 1999), which may be the reason why at long RSIs repetition benefits occurred also for partial repetitions.
Further, hand repetition effects did not decrease at long RSIs, indicating that effector activation may last longer than 2,000 ms. This observation stands in contrast to previous studies showing a decrease of response repetition benefits with increasing RSIs (e.g., Bertelson & Renkin, 1966). However, repetition effects also depend on participant’s expectations (Duthoo et al., 2012; Kirby, 1976; Soetens et al., 1985) and participants often expect repetitions rather than alternations, even when they are equally distributed (Duthoo et al., 2012). Thus, participants may have expected repetitions rather than alternations in the current task resulting in benefits even at long RSIs (Kirby, 1976)
In E-I sequences, hand repetition costs (in contrast to benefits in E-E sequences) provide further evidence that activation of the effector occurred in the previous trial, thus requiring more inhibitory effort to imagine the movement in the subsequent trial. In E-I sequences hand repetition costs decreased when RSIs varied between blocks, though the decrease was slow and hand repetition costs were still observable at the 1,300 ms RSI. When RSIs varied within blocks hand repetition costs did not decrease. When RSIs are blocked, prediction of the stimulus onset is possible (Grosjean et al., 2001). Such predictions may have fostered faster decay of effector activation to prepare for the next trial.
Time course of effector-specific inhibition
Data patterns of all action mode sequences in which at least one of the trials was an imagination trial provide evidence for effector-specific inhibition. In E-I sequences hand repetition costs were observed, indicating that executing the movement in the previous trial resulted in an activation of the effector, thus requiring more inhibitory effort to imagine the movement using the same effector in the subsequent trial. Further, in I-I sequences at the 200 ms RSI (only in movements with target alternation) and I-E sequences at the 200, 700, and 1,300 ms RSIs hand repetition costs occurred. This indicates that inhibition of the effector in the previous trial had to be overcome to execute or imagine the movement using the same effector in the subsequent trial (Bart et al., 2020; Rieger et al., 2017). Taken together, the data indicate that effector-specific inhibition contributes to motor imagery.
Only sequences, in which the previous trial is an imagination trial (I-I sequences and I-E sequences) are informative regarding the time course of effector-specific inhibition. In both, I-I sequences (in movements with target alternation) and I-E sequences, hand repetition costs decreased, indicating that effector-specific inhibition is subject to decay. Further, hand repetition costs eventually disappeared at longer RSIs. Thus, even though effector-specific inhibition continues to be present for some time after imagination is complete (cf. Rieger et al., 2017; Scheil & Liefooghe, 2018), it is not a very long-lasting form of inhibition, but instead decays relatively quickly within the time frame of 2 s.
In complete repetitions (i.e., movements with hand and target repetition) in I-I sequences, we observed no significant hand repetition effects at the 200 ms RSI. Furthermore, hand repetition benefits occurred with increasing RSIs. Why did we not observe hand repetition costs? Complete repetitions are a special case, as the stimulus and the response are the same in trial N and trial N-1. When a stimulus is not repeated, it is necessary to pass through various processing stages during information processing. Complete repetitions allow bypassing those processing stages, which results in a fast response (Horner & Henson, 2008; Marí-Beffa et al., 2012). Thus, effector-specific inhibition and mechanisms behind complete repetition may have contributed simultaneously to the observed effects. Effector-specific inhibition and complete repetition benefits may initially be of similar strength, resulting in no significant hand repetition effects at the 200 ms RSI. However, costs due to effector-specific inhibition may decrease relatively fast (as indicated by the results of movements with target alternation in I-I sequences and the results of I-E sequences), whereas complete repetition benefits may decrease relatively slowly (as indicated by results from E-I and E-E sequences), resulting in hand repetition benefits with increasing RSIs.
How do global inhibition and effector-specific inhibition work together?
Although both global and effector-specific inhibitions were subject to decay, they differed in their time course. Global inhibition decreased over time, but the decrease was rather slow and switch benefits were still observable at the 2,000 ms RSI. Two different processes seem to contribute to global inhibition. Our results and previous results (Bart et al., 2020; Rieger et al., 2017) indicate that global inhibition is partly event-based, that is, formed on a trial-to-trial basis. For instance, switch benefits in E-I sequences compared with I-I sequences indicate that in I-I sequences global inhibition took place in the previous imagination trial, affecting performance in the subsequent trial. We will refer to this form of global inhibition as phasic global inhibition, which reflects the temporary increase of global inhibition as a response to a certain event, for example, a stimulus indicating that a movement has to be imagined (cf. Posner, 2008, for similar assumptions regarding phasic alertness). In addition, previous studies show that global inhibition is higher in contexts in which motor imagery has to be performed more often (pure imagination blocks compared with mixed imagination and execution blocks, cf. Bart et al., 2020; Rieger et al., 2017). This indicates that global inhibition is also partly context-based and may be akin to a proactive form of inhibition (Elchlepp et al., 2016). We will refer to this form of global inhibition as tonic global inhibition, which reflects the overall readiness to prevent all actual movements at any time in contexts, in which movements have to be imagined (cf. Posner, 2008, for similar assumptions regarding tonic alertness). Participants may always maintain a certain level of tonic global inhibition during the present task (as they know they may have to imagine a movement any time), slowing down the decay of phasic global inhibition.
In contrast, effector-specific inhibition decayed quickly within a time frame of 2 s, which indicates that it is mainly formed on a trial-to-trial basis and strongly depends on the movement requirements within a certain trial. When a trial is over, effector-specific inhibition quickly dissipates.
Those differences between global and effector-specific inhibition resemble differences between various forms of motor command inhibition in other inhibitory tasks. For instance, for Go-No Go tasks, it has been proposed that a non-selective (global) form of inhibition may temporally slow down the response, which is later followed by a more selective form of inhibition (Coxon et al., 2007). A similar model can be assumed for inhibition in motor imagery. Due to tonic global inhibition (Bart et al., 2020; Rieger et al., 2017), the motor threshold for any actual responses may be increased in contexts in which motor imagery may be performed. Once participants know (i.e., after the stimulus for a trial appears) that they now have to imagine the movement, global inhibition is increased further (phasic global inhibition, as indicated by the observed trial-to-trial effects in the present study). As effector-specific inhibition is tightly bound to the specific movement requirements, it may only contribute to motor imagery as soon as the exact movement features (e.g., which hand performs the movement) are known. After imagination is complete, effector-specific inhibition decays fast and phasic global inhibition decays as well. However, a certain level of (probably tonic) global inhibition is maintained as participants know that they may have to imagine a movement any time, slowing down the decay of phasic global inhibition.
Limitations and perspectives
A limitation of the present study is that we investigated very simple hand movements. Movements performed in everyday life are often more complex and involve the whole body. One may speculate that different forms of inhibition are not in all instances equally effective and that their contribution to motor imagery may therefore vary as a function of movement complexity and depend on which effectors are involved. For instance, if an imagined movement involves the simultaneous use of the right hand and the left foot, global inhibition might be more efficient and thus more pronounced than effector-specific inhibition. Which form of inhibition is more pronounced may in turn affect how quickly inhibition decays over time. Thus, further research is necessary to investigate which factors may influence inhibition in motor imagery and its decay over time. This should lead in the long term to a more detailed model of how different forms of inhibition work together during motor imagery.
With regard to applications of motor imagery, it may be of interest to investigate whether the necessity to inhibit actual movements in motor imagery may cause undesired effects. For instance, one may raise the question of whether imagination of a movement shortly before its actual execution (as sometimes used by athletes prior to a competition as a pre-performance routine) may have detrimental effects on performance. Based on the present results a tentative assumption is that this is not the case (as long as there is a break of approximately 2 s between imagination and execution) because inhibition seems to decay rather quickly. However, more thorough research, which should also involve more complex movements than the ones used in the present study, is needed to satisfactorily answer this question.
Conclusion
In conclusion, our results indicate that global inhibition and effector-specific inhibition contribute to motor imagery. Global inhibition seems to decay more slowly than effector-specific inhibition. Global inhibition may not only be formed on a trial-by-trial basis, but may be at least partly implemented in all contexts in which motor imagery has to be performed. Effector-specific inhibition may contribute to motor imagery as soon as the exact movement parameters are known and decays fast after the imagined movement has been performed.
Supplemental Material
2020_06_25_Bart,_Koch,_Rieger_supplement_time_course – Supplemental material for Decay of inhibition in motor imagery
Supplemental material, 2020_06_25_Bart,_Koch,_Rieger_supplement_time_course for Decay of inhibition in motor imagery by Victoria KE Bart, Iring Koch and Martina Rieger in Quarterly Journal of Experimental Psychology
Footnotes
Acknowledgements
The authors thank Alexander Bäuerle, Andrea Brasik, and Maximilian Hausmann for their help with data collection.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.
Data accessibility statement
Notes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.