
Abstract
Coregulation refers to the process by which relationship partners form a dyadic emotional system involving an oscillating pattern of affective arousal and dampening that dynamically maintains an optimal emotional state. Coregulation may represent an important form of interpersonal emotion regulation, but confusion exists in the literature due to a lack of precision in the usage of the term. We propose an operational definition for coregulation as a bidirectional linkage of oscillating emotional channels between partners, which contributes to emotional stability for both partners. We propose several distinctions and raise unanswered questions that will need to be addressed in order to understand the relevance of coregulation for well-being in adulthood.
Emotion regulation refers to the processes by which we influence the occurrence, duration, and intensity of subjective experience, expression, and physiological arousal associated with emotions (Cole, Martin, & Dennis, 2004; Gross, 1999; Thompson, 1994). Empirical research suggests that the ability to modulate emotional responding in a way that supports one’s goals and maintains physiological equilibrium is critical for psychological, physical, and social well-being across the lifespan (Eisenberg, Fabes, Guthrie, & Reiser, 2000; Gross, 2002; John & Gross, 2004). The child development literature has always explicitly considered interpersonal processes as central to how infants achieve successful emotion regulation. In that context, “coregulation” refers to an adult and infant together forming a dyadic emotional system and coconstructing optimal affective states during social interactions (Feldman, 2003; Tronick, 1989). The adult literature on emotion regulation has traditionally given less attention to the role of interpersonal processes, but this has been changing and a growing literature refers to coregulation in the context of adult romantic relationships (Butner, Diamond, & Hicks, 2007; Saxbe & Repetti, 2010; Schoebi, 2008). This is an exciting shift and initial results confirm that a range of interpersonal emotional processes play a role in shaping the daily emotional experiences and physiological responses of adult romantic partners.
Research on interpersonal emotion regulation in adulthood has been hindered, however, by the lack of operational definitions (Butler, 2011). Partners in close relationships demonstrate a vast range of emotional and physiological interconnectedness, and the term coregulation has been used to refer to numerous potentially distinct phenomena. For example, romantic partners’ emotional experiences can covary day to day or show coupling in their rates of change (Butner et al., 2007); one person’s emotions experienced while separate from their partner can be transmitted to the partner upon reunion (Schoebi, 2008); and partners’ physiological responses can become correlated due to conflict (Saxbe & Repetti, 2010). Each of these possibly distinct processes has been referred to as coregulation. We suggest that this breadth of usage may blur important distinctions and obscure our understanding of these important interpersonal emotional processes.
An Operational Definition of Coregulation
We propose that it may be scientifically useful to define coregulation as one specific form of interpersonal emotion regulation, or “social affect regulation” as it is sometimes referred to. Specifically, we suggest an operational definition of coregulation as a bidirectional linkage of oscillating emotional channels (subjective experience, expressive behavior, and autonomic physiology) between partners, which contributes to emotional and physiological stability for both partners in a close relationship. In the following sections we expand and clarify these ideas and review literature relevant to each aspect of our proposed definition. One advantage of this definition is that it would promote the empirical study of whether, and how, relationship partners contribute to each others’ emotional and physiological balance. If coregulation does in fact occur in a form that matches our definition, it would represent one central mechanism by which relationship quality could contribute to health and well-being.
Bidirectional Linkage of Emotional Channels
One central point of agreement in the literature is that coregulation is a property of interpersonal systems that cannot be reduced to the behaviors or experiences of the individuals involved (Cole et al., 2004; Diamond & Aspinwall, 2003; Feldman, 2007; Sbarra & Hazan, 2008). The emphasis is not on individual emotion regulation, but rather on the ways in which both partners’ emotions influence each other in a recursive pattern of mutual regulation. Therefore, the first criteria that we and others suggest would provide evidence for coregulation is bidirectional statistical dependence between relationship partners over time on some measure of emotion (experience, behavior, physiology) (Butner et al., 2007; Feldman, 2007; Sbarra & Hazan, 2008).
Fairly extensive evidence suggests that such bidirectional dependence exists in parent–infant interactions (Brazelton, Koslowski, & Main, 1974; Feldman, 2003; Field, Healy, Goldstein, & Guthertz, 1990; Lester, Hoffman, & Brazelton, 1985; Stern, 1974). Terms such as mutual influence, reciprocity, matching, or affective attunement have been used to refer to the dynamic process whereby the parent and infant achieve behavioral and emotional coordination (Beebe & Stern, 1977; Tronick, Als, Adamson, Wise, & Brazelton, 1978). This affective coregulation, often indexed by emotional synchrony in dyads (covariation of emotion over time), has been argued to provide the basis for infants’ emotional self-regulation because by coconstructing optimal emotional states the mother extends and scaffolds the infant’s emerging self-regulatory capacity (Gianino & Tronick, 1985; Tronick, 1989; Tronick & Gianino, 1986). Similarly, the term psychobiological attunement refers to the psychological and physiological states of two individuals becoming linked to one another in the context of attachment relationships (Diamond, 2001; Field, 1985; Hofer, 1984, 1994; Kraemer, 1992; Sbarra & Hazan, 2008). It is argued that during social interactions securely attached partners provide contingent stimulation and soothing that helps to maintain both partners within an optimal arousal zone (Field, 1985; Sbarra & Hazan, 2008). In support of this, statistical dependencies between parents and infants have been shown across a wide range of arousal indicators, including eye gaze, facial expressions, prosody, speech rhythms, attention, and autonomic physiology (e.g., heart rate) (for a review see Field, 1985).
Recently, research has begun to explore coregulation in adulthood as indicated by statistical dependencies between romantic partners’ emotional experiences. Butner et al. (2007) used a diary methodology in which both partners in romantic couples reported daily on their positive and negative emotional experience. Coregulation was defined as both the covariation in partners’ daily affect and coupling of the rates of change between the partners. Results indicated that partners’ level of positive and negative affect covaried, above and beyond the influence of their shared daily interactions. This effect was greater on days when couples spent more time together, suggesting the partner’s presence is an important part of coregulation.
Morphostatic Oscillating Emotional Channels
One point that is implicit in much of the research on parent–child dyads is that coregulation involves an oscillating sequence of emotional stimulation and arousal modulation that enables parent–infant dyads to dynamically organize behavior and physiology within optimal bounds (Feldman, 2003; Field, 1985; Tronick, 1989). What constitutes “optimal” depends upon both the internal resources of the partners (e.g., does the infant need to sleep or is he ready to play?) and external demands (e.g., is the dyad reading quietly or playing a challenging game?). Thus coregulation would entail oscillating emotions that fluctuate appropriately for the situation around a globally stable (morphostatic) level. Although this point is rarely made explicitly, it suggests a second criterion for establishing evidence that coregulation is occurring. Specifically, coregulation should entail relationship partners’ emotions oscillating in a coupled fashion around fluctuating levels that are matched to the internal and external demands of the situation, not around an extreme level or around an escalating or de-escalating trajectory (see Panel A vs. Panel B, Figure 1). For example, if a mother and infant are successfully coregulating, their mutual emotional state should vary within the bounds of mildly negative to fairly positive, or within the range of comfortable arousal. If the dyad becomes strongly negative or overly aroused and fails to counter this with a return to a more quiescent state—in other words if their emotional oscillations move away from their optimal range of functioning—then this would imply a failure of coregulation.
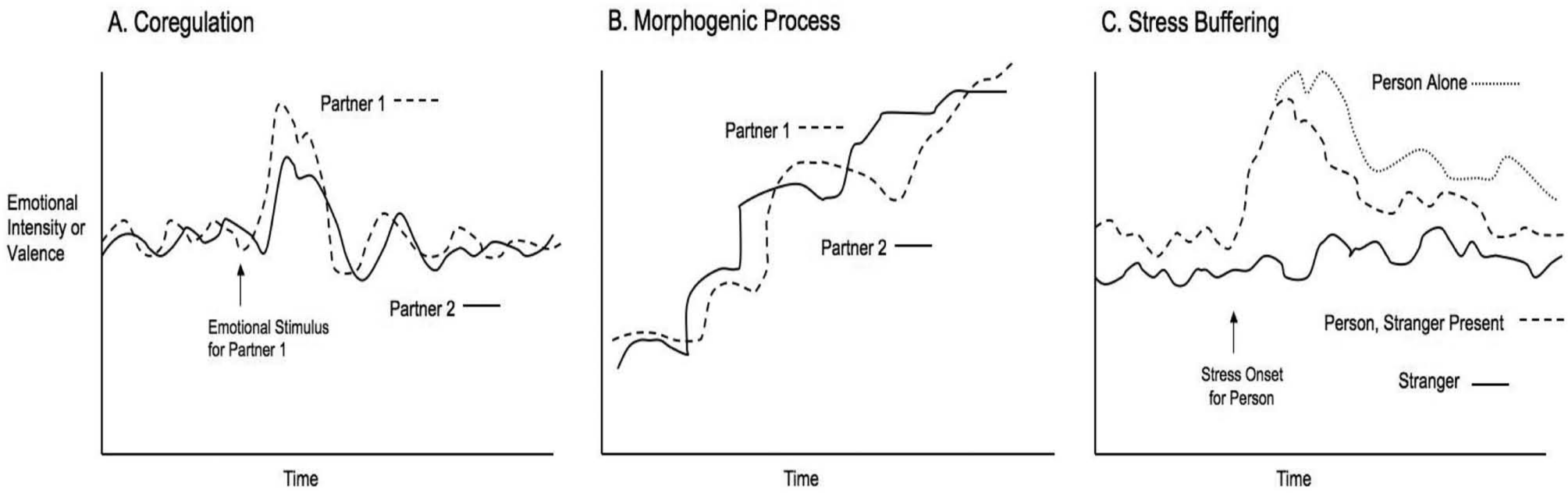
Panel A shows our operationalization of coregulation. Panel B shows an example of a morphogenic process, such as negative reciprocity, transmission, or contagion. Note that both coregulation and the morphogenic process show emotional covariation (the partners’ emotional levels are correlated over time) and coupling (one partner appears to be influencing the rate of change in the other partner’s oscillations). The processes are distinguished, however, by the presence of a linear trend in the case of morphogenesis and the presence of coupled dampened oscillations following a stimulus in the case of coregulation. Panel C shows our hypothesized pattern for stress buffering, in which there is a lack of correlation between the two people’s emotional variations, although the presence of the stranger does result in a quicker return to homeostasis following a stressor.
Several lines of research with parent–infant dyads provide information about both bidirectional linkage of partners’ emotional oscillations and the level around which they are fluctuating, thereby providing evidence for our first two criteria for coregulation. For example, Feldman (2003) focused on positive emotions and compared father–infant and mother–infant dyads. Coregulation was indicated by emotional synchrony (i.e., covariation of emotions over time). Mother–infant synchrony was inversely related to infant negative emotionality, whereas father–infant synchrony was positively related to the degree of positive arousal. This pattern suggests that both mothers and fathers provide important scaffolding for the development of infant emotion regulation, but that they do so in different ways, with mothers providing modulation of negative emotions and fathers providing coordination of high-intensity positive arousal (Feldman, 2003). In contrast, deficits in coregulation in depressed mother–infant dyads have been shown both in terms of decreased contingency of their emotional behaviors and also in their more negative and less positive affective levels during interaction (Tronick & Gianino, 1986).
Emotional and Physiological Allostatic Balance
Contemporary models of emotional and physical well-being suggest that stability, adaptability, and health are maintained through dynamic patterns of organized variability (Thayer & Sternberg, 2006). Allostatic balance refers to this stability through change, or in other words, coordinated variation of psychological and biological systems that optimize performance and minimize costs (Sterling, 2003). For example, our optimal energy expenditure is constantly changing due to internal and external demands. To support this, our heart rate fluctuates continuously around a mean level that is itself fluctuating in response to energy demands (Thayer & Lane, 2000, 2009). Healthy allostatic balance is characterized by rapid psychological and physical responses to threats, challenges, and opportunities followed by a quick return to an energy-efficient resting state (Thayer & Lane, 2000, 2009; Thayer & Sternberg, 2006).
One intriguing idea is that coregulation may contribute to health and well-being by supporting allostatic balance for both partners in a close relationship (Diamond, Hicks, & Otter-Henderson, 2008; Field, 1985; Hofer, 1984, 1994; Sbarra & Hazan, 2008). The social baseline model of emotion regulation provides a framework for understanding why such a process might be highly functional across the lifespan and therefore may have evolved (Coan, 2008, 2010). The social baseline theory proposes that one central and highly effective way in which psychological and biological regulation is achieved is through simple proximity to trusted others. Humans have evolved as a social species and are hard-wired to draw on the stable support and resources of close others to reduce threat responding and promote emotional well-being. One way this might occur (although not the only way) is through coregulation, whereby psychological and biological interdependencies with a trusted partner may automatically restore emotional and physiological balance with limited cognitive or metabolic cost (Coan, 2008, 2010).
Evidence for the connection between coregulation and allostatic balance is clearest in research on parent–infant relationships. Research with humans, monkeys, and rats suggests that infants’ behavior and physiology are initially regulated by the sensory characteristics of the caregiver, such as warmth, smell, and touch. The caregiver’s behavior and physiology are in turn reciprocally responsive to stimulation by the infant, again through senses such as touch and smell, but also behaviors such as crying (Field, 1985; Hofer, 1984, 1994; Kraemer, 1992). In other words, both the infant’s and the parent’s emotional and biological dynamic stability are partially dependent upon the other. Such a state has been argued to also be present in adulthood in the context of secure attachment relationships. Repeated social contact with a rewarding partner is argued to lead to a conditioned response, whereby that partner is associated with psychological security and physiological quiescence (Sbarra & Hazan, 2008). Attachment behaviors such as cuddling, touching, and sexual contact activate oxytocin and opioid systems, which together induce pleasure and alleviate distress, thereby providing a physiological basis for felt security and contributing to both partners’ physiological and emotional balance (Diamond, 2001; Sbarra & Hazan, 2008).
Much of the empirical research relevant to coregulation and allostatic balance has been done with infant animals separated from their mothers (Field, 1985; Kraemer, 1992; Sbarra & Hazan, 2008). The central premise of this work is that specific biological and behavioral features of the mother automatically regulate the physiological and behavioral systems of the infant. These “hidden regulators” only become apparent when they are removed and the infant loses its ability to maintain biological and behavioral balance. A fairly extensive and systematic body of research with rats and monkeys supports this by demonstrating wide-ranging physiological disturbances in infants separated from their caregivers, even if the infants are provided with all other necessities. Field (1985) reviewed similar research with human infants and concluded that emotional or physical unavailability of the caregiver results in a broad range of disrupted biological and behavioral functions in the infant. Although there has been less research on adult relationships, Vormbrock (1993) reviewed work on extended separations between married partners and found that such separations were generally associated with increased negative emotions, sleep disturbances, and a variety of other behavioral and psychological dysregulations. Similarly, a recent study investigated physiology (cortisol), behavior, and emotional experience in romantic couples briefly separated due to one partner traveling and found similar effects, with the homebound partner and individuals high in attachment anxiety especially showing disturbances in sleep, subjective stress, and physiological stress responses (Diamond et al., 2008). Although these findings suggest that disrupting coregulation impairs allostatic balance, what is missing is evidence that restoring coregulation restores balance. Thus an important direction for future research is to assess allostatic balance before and after a disruption to coregulation.
Summary of Criteria for Coregulation
In summary, for the purposes of empirical study we suggest several criteria that would provide evidence for coregulation. First, it would require bidirectional statistical dependence between relationship partners over time on some measure of emotion. Second, it would entail relationship partners’ emotions oscillating in a coupled fashion around a fluctuating optimal level that is globally stable (see Panel A vs. Panel B, Figure 1). In other words, the joint emotional state of the dyad should be morphostatic, meaning that it is dominated by negative feedback processes that maintain overall emotional stability. Third, if coregulation is relevant to allostatic balance it would be most obvious when one or both partners were perturbed away from a balanced state and would appear as dampening oscillations converging on a more optimal level given internal and external demands (Panel A, Figure 1).
Distinguishing Coregulation from Related Constructs
Morphogenic Processes versus Coregulation
One important distinction is between morphogenic and morphostatic interpersonal emotional processes (Butler, 2011). Morphogenic processes are dominated by positive feedback and entail amplified deviations resulting in an altered emotional state (see Panel B, Figure 1). In contrast, morphostatic processes depend on negative feedback to dampen or reverse deviations, returning the system to a stable state (see Panel A, Figure 1) (Butler, 2011). Based on the research reviewed before, we suggest that coregulation is best understood as a morphostatic process involving partners’ emotions oscillating around optimal levels that are globally stable.
In contrast, there are a number of emotional processes that are better understood as morphogenic, including emotion reciprocity (Cordova, Jacobson, Gottman, Rushe, & Cox, 1993), emotion transmission (Larson & Almeida, 1999), and emotion contagion (Hatfield, Cacioppo, & Rapson, 1994). Each of these are conceptualized in the literature as resulting in an elevated or altered emotional state in one or both partners (see Panel B, Figure 1). For example, reciprocity often occurs in conflict interactions and results in a high joint level of negative expression (Cordova et al., 1993). Similarly, emotion transmission and contagion both involve one partner’s emotional state affecting the other’s, with the overall effect being to shift the joint emotional state of the dyad into a new affective space (e.g., Hatfield et al., 1994; Larson & Almeida, 1999).
Much of the confusion around the term coregulation in the literature on adult romantic relationships is due to not distinguishing between morphostatic versus morphogenic interpersonal emotional processes. Several findings have been referred to as coregulation that we suggest would be better understood as examples of morphogenic processes. One example is a diary study in which married couples reported their emotional experience on the dimensions of “hard affect” (angry–calm) and “soft affect” (sad–upbeat) six times per day for 7 days (Schoebi, 2008). Analyses focused on reunion occasions, defined as time points when partners were together but they reported having been apart at the prior assessment. Individuals who scored higher on interpersonal insecurity showed greater partner influence on changes in hard affect when they reunited, and husbands who were higher on perspective taking showed greater influence from their wives’ soft affect. We suggest that this pattern of findings may be best understood as an example of emotion transmission, rather than coregulation. Further information is required, however, to establish our case. The analyses did not include a linear trajectory over time and so we do not know whether partners’ emotions were oscillating around a stable (morphostatic) or changing (morphogenic) state. We suggest that linear trajectories be routinely assessed when investigating coregulation in order to distinguish it from morphogenic emotional processes.
Saxbe and Repetti (2010) used a similar design to assess both experiential and physiological covariation in couples. In this study, both marital partners reported their positive and negative mood four times a day on 3 separate days. At each time point they also provided a salivary cortisol sample. Results showed significant covariation of negative, but not positive, emotional experience between partners. This effect was stronger for couples in which the husband was less satisfied with the marriage. In addition, partners’ cortisol levels also covaried, and again this relationship was moderated by marital satisfaction such that couples with lower marital satisfaction showed greater cortisol linkage. The authors frame their research in the context of coregulation, but also interpret their results as an example of negative reciprocity, a morphogenic process whereby less satisfied couples engage in a reciprocal pattern of escalating conflict (Saxbe & Repetti, 2010). This research provides critical insight into the ways that relationship quality might translate into hysical health outcomes through the escalation of negative emotions. Nonetheless, the results clearly do not indicate a morphostatic emotion regulatory pattern.
Interpersonal Affect Regulation versus Coregulation
Individuals are constantly engaging in a wide range of behaviors to achieve a variety of emotional goals (Diamond & Aspinwall, 2003; Gross, 1998; Thompson, 1994). Many of these emotion regulatory efforts are targeted at changing either a social partner’s emotions or the emotional tone of the relationship, and these efforts have been referred to as “interpersonal affect regulation” (Niven, Totterdell, & Holman, 2009). Many of these strategies are clearly distinct from coregulation as we have operationalized it. For example, one partner may try to induce guilt in the other to get him or her to comply with some demand. Although this would involve an attempt at interpersonal emotion regulation, it does not entail a bidirectional morphostatic process.
Other interpersonal affect strategies are less clearly distinguished from coregulation. For example, providing support, communicating empathy, or even distracting one’s partner may help both partners return to a more positive mood state (Diamond, 2001; Diamond & Aspinwall, 2003). Other examples include dyadic coping, whereby the stress management resources of partners are engaged to restore homeostasis between them (Bodenmann, 2005; Randall & Bodenmann, 2009), and capitalization, which occurs when one partner shares positive events and the other responds enthusiastically, a process that has been shown to enhance positive affect and relationship well-being (Gable, Reis, Impett, & Asher, 2004). The distinction we propose is that coregulation involves an oscillating pattern of emotional interdependency within a relationship. In contrast, supportive behaviors, dyadic coping, and other related processes all describe interpersonal behavioral patterns that partners might engage in that may, or may not, promote a coregulatory state. As such, these forms of interpersonal affect regulation may sometimes contribute to a couple experiencing coregulation, but they in themselves are not examples of coregulation.
Stress Buffering versus Coregulation
Stress buffering refers to the phenomena whereby the presence of others reduces stress responding (Uchino, 2004; Uchino, Cacioppo, & Kiekolt-Glaser, 1996). At first glance stress buffering and coregulation appear similar because they both involve a return to allostatic balance as the result of an interpersonal process, but it may be possible to distinguish them on the basis of their degree of bidirectionality and magnitude of effect. As discussed earlier, the social baseline model proposes that the mere presence of others helps distribute the risk a threat has on any given individual, because there are others that can help look for threats, the chance of being individually targeted is reduced, and shared resources can be brought to bear (Coan, 2008). This idea of risk distribution provides the basis for the well- supported stress-buffering hypothesis, which suggests that the presence of any individual, even a stranger, will result in a reduced physiological response to stressors (for a review see Uchino et al., 1996).
In contrast, a central distinction made by the social baseline model is between risk distribution and load sharing in close relationships (Coan, 2008). Load sharing refers to the fact that a close, committed relationship partner can be counted on to actively promote your health and well-being. This goes beyond risk distribution, and includes procuring and sharing resources, providing care and emotional comfort, and helping to nurture and protect your offspring. A load-sharing relationship thereby reduces the cost of many metabolically expensive activities, including maintaining physiological and emotional homeostasis. Thus, even though a stranger may reduce stress responding to some degree, the presence of a load-sharing partner should produce additional physiological and emotional stress reduction. In addition, one way that this might occur is through coregulation. Although load sharing does not necessarily entail emotional dependencies between partners (e.g. divorced partners can share the effort of childcare without sharing emotional states), it may provide a context in which coregulation is likely to occur. Due to the mutuality of a load-sharing relationship, partners in such a relationship should be more likely to show bidirectional, truly interconnected emotional and physiological responses as compared to a stranger. Thus coregulation likely implies the presence of a load-sharing relationship, even though the reverse is not true. For example, in a load-sharing relationship, if one partner were faced with a stressor we would expect him or her to be able to draw on the other’s calmer state as an emotional resource, but at the same time both partners should be responsive to the other’s increasing or decreasing anxiety, creating a coupled oscillating system. In contrast, a stranger would have much less reason to react emotionally to the other person’s stress, even though the stranger’s presence might provide some calming influence. Thus, the stress-buffering effects of a stranger may manifest in a fairly unidirectional way, as shown in Panel C of Figure 1, while the coregulatory effects of a close partner may manifest in a more bidirectional pattern of dampening oscillations following a stressor, as shown in Panel A of Figure 1.
We are not aware of any empirical evidence that speaks to whether stress reduction due to a stranger takes a different dyadic temporal pattern than that due to load-sharing partner; that question awaits future research. However, one study has addressed the hypothesis that strangers and close partners may provide separate additive stress-reducing effects (Coan, Schaefer, & Davidson, 2006). Women were mildly shocked while in an MRI scanner in each of three conditions: (a) alone, (b) holding a stranger’s hand, and (c) holding their husband’s hand. Results showed that both strangers and husbands reduced activity in neural circuits associated with threat responding. Additional reductions were observed, however, when women held their husband’s hand. Furthermore, the quality of the relationship was also important, as women in the highest quality relationships showed the lowest degree of threat.
Unanswered Questions
Scientists wishing to understand coregulation, especially in adulthood, have a lot of work left to do. The recent shift to studying coregulation in adults is an exciting foray into new territory, but it has raised as many questions as it has answered. The following are a few of the questions that appear to us to be the most pressing.
Does Coregulation Exist in Adulthood?
Based on theory and prior research we have suggested that coregulation might be operationally defined as a morphostatic process which occurs in close relationships, operates across emotion channels (experience, behavior, physiology), and contributes to emotional and biological allostasis for both partners. Although we are not aware of any single study that has assessed all these criteria, enough converging evidence exists in the child development literature to lead us to conclude that coregulation, as we have operationalized it, likely occurs in parent–child dyads. The research on adult romantic relationships is much smaller, however, and almost all studies have only assessed covariation, which only satisfies one criterion. Covariation, by itself, could be driven by any of a number of interpersonal emotional dependencies, ranging from negative reciprocity, to emotional contagion, to coregulation. The work by Butner et al. (2007) goes the furthest toward establishing the presence of coregulation in adult couples by assessing both covariation and coupled oscillations. Their finding of significant coupling establishes that partners were oscillating in a coordinated pattern, but the levels and stability of their trajectories were not reported. In addition, coupling was only found for positive emotion, and the frequency of oscillations was in the range of one affective cycle every 5 days, which, as the authors point out, may suggest longer-term entrainment, rather than shorter-term coregulation. Finally, although their model includes a dampening term that indicates whether the oscillations converge over time, their study was not designed to assess situations in which partners were perturbed away from their baseline. As such, it was not intended to provide evidence relevant to allostatic balance. Thus, to the best of our knowledge, there does not exist any conclusive evidence to date that coregulation exists in adult relationships in a form similar to that suggested by theory. One pressing issue for future research, therefore, is to establish whether coregulation as conceptualized in theory (and operationalized here) does in fact extend across the lifespan.
What Is the Time Scale for Coregulation?
Establishing whether coregulation exists in adulthood will require establishing the time scale over which it occurs. The results from Butner et al.’s (2007) study show interpersonal emotional dependencies that are largely compatible with our operationalization of coregulation. The time scale that they assessed (one observation per day), however, did not allow for the observation of emotional oscillations taking place within a 24-hour period. It is currently unknown whether coregulation in couples (if it exists) transpires over the course of seconds, minutes, hours, or days. Furthermore, the issue of time scale has implications for understanding the interplay of regulatory costs and benefits involved in coregulation. Extensive evidence suggests that self-regulatory capacity is finite and that exerting self-regulation depletes our ability for subsequent regulatory tasks (Baumeister & Alquist, 2009). One of the functions of coregulation, therefore, may be to scaffold one partner’s self-regulation when he or she is fatigued, but this raises the issue of what happens to the other partner in this scenario, the one providing the self-regulatory support. For example, if one partner comes home from work exhausted and annoyed, and if coregulation is an automatic process whereby the partners’ psychological and physiological states become coupled in each others’ presence, would this exact a toll on the other partner? In keeping with this, one diary study showed that women tended to increase their work efforts in the home to compensate for when their male partners had a particularly stressful day at work (Bolger, DeLongis, Kessler, & Wethington, 1989). But even if it does exact an immediate cost, might this balance out over the longer term when it is his or her turn to gain coregulatory benefits from the other? Answers to these questions will be important for establishing if, how, and under what conditions coregulation contributes to allostatic balance in adulthood.
Which Biological Subsystems Are Relevant to Coregulation?
Research on coregulation generally does not address the fact that different biological subsystems operate differently. For example, research on coregulation in parent–infant dyads typically focuses on indices of autonomic activity (Field, 1985), while research with adult romantic partners has primarily focused on cortisol (Diamond et al., 2008; Saxbe & Repetti, 2010). Autonomic indices fluctuate over the course of minutes, however, while cortisol trajectories occur over hours. In addition, autonomic activity is typically associated with attentional and emotional responding (Kreibig, Wilhelm, Roth, & Gross, 2007; Thayer & Lane, 2000), while cortisol responses are thought to be indicative of generalized stress responding, particularly in response to social-evaluative threat (Denson, Spanovic, & Miller, 2009). Therefore, if our argument is correct that coregulation is a morphostatic process involving coupled emotions, then autonomic indices should be more sensitive for assessing it than cortisol, which itself may be a better indicator of coupled responding to longer duration stressors both within the relationship (extended conflict) and outside it (shared environmental stressors).
A related issue is that some biological subsystems such as oxytocin or parasympathetic tone may provide a supporting substrate for coregulation, but they do not show between- artner couplings themselves. Oxytocin is associated with affiliation, social bonding, and soothing (Taylor et al., 2000), and its inherently rewarding properties have been suggested to reinforce attachment behaviors, which could facilitate coregulation (Sbarra & Hazan, 2008). High parasympathetic tone, as indexed by heart rate variability, is associated with greater self-regulatory capacity, emotional flexibility, and social competence and connectedness (Porges, 2003, 2007; Smith et al., 2011; Thayer & Lane, 2000, 2009). These characteristics may contribute to individual differences in the capacity for engaging in coregulation. In support of this, in a study of mother–infant dyads greater behavioral synchrony was predicted by higher infant parasympathetic tone at a prior time, suggesting that the development of parasympathetic control within infants is critical for supporting behavioral coregulation at a later stage (Feldman, 2006).
If Coregulation Exists in Adulthood, How Is it Related to Attachment Security?
Throughout the literature, coregulation and attachment security are assumed to be related. Fairly strong evidence suggests a circular relationship in childhood, such that early biological and behavioral coregulation provides the scaffolding for the emergence of attachment security, which in turn reinforces coregulation (Evans & Porter, 2009; Feldman, 2007). It is possible that this circular relation between felt security and coregulation is carried throughout life and repeated within each attachment relationship, but we are not aware of any research tracking the development of attachment security and coregulation across time in adult romantic couples. In addition, the situation is more complicated in adulthood because both partners enter the relationship with a previous attachment history and habitual attachment style (Mikulincer, Shaver, & Horesh, 2006; Pietromonaco & Barrett, 1997; Pietromonaco, Barrett, & Powers, 2006). It is unknown how this might influence the dynamics of coregulation and security. Does coregulation promote secure attachment? Does insecure attachment alter the quality of coregulation, or even preclude it? The only study we know of that has investigated coregulation and attachment styles in adulthood found that anxiety and avoidance did in fact moderate indicators of coregulation, but the results were not always in the predicted direction (Butner et al., 2007). Thus the jury is still out regarding if and how attachment security and coregulation are related in adulthood.
Recommendations and Conclusions
Clearly, relationship and emotion researchers still have a lot of work to do to understand the fundamentals and implications of emotional coregulation, especially in adulthood. To make progress, we argue that future research should include observations at several levels of resolution (second-to-second, hour-to-hour, day-to-day) and adopt an analytic strategy that includes: (a) average emotional levels (to establish the presence of a nonextreme set point), (b) linear trends (to investigate the presence of morphogenic processes in addition to coregulation), (c) covariation and coupling (to investigate both forms of between-partner interdependency), and ideally (d) dampening in response to a perturbation (to show relevance to allostasis). The statistical models suggested by Boker, Butner, and colleagues could be adapted to address all these issues (Boker & Laurenceau, 2006, 2007; Boker & Nesselroade, 2002; Butner, Amazeen, & Mulvey, 2005; Butner et al., 2007).
Understanding if and how partners coregulate, and when this occurs, would provide us with important information about the role of interpersonal emotion regulation in promoting health and well-being. Coregulation should be adaptive to the degree that it tailors the emotional state of relationship partners to achieve their goals in a metabolically efficient manner. A morphostatic process such as coregulation may help to prevent undesirable emotional escalation from occurring or to return the dyad quickly to quiescence when a stressor is removed. If this were true then coregulation may contribute to flexible allostatic balance, which is known to be critical for physical and psychological well-being (Thayer & Brosschot, 2005; Thayer & Lane, 2007; Thayer & Sternberg, 2006), but this issue awaits empirical verification.
Footnotes
Author note:
The authors would like to thank Noel Card, Michael Rohrbaugh, and David Sbarra for their comments on earlier versions of this manuscript. This research was supported in part by the Frances McClelland Institute for Children, Youth, and Families, in the Norton School of Family and Consumer Sciences at The University of Arizona. Information about the Frances McClelland Institute is available at: ![]()