
Abstract
Parkinson's disease (PD) ranks the second place among neurodegenerative diseases in terms of its morbidity, which affects 1-2% people aged over 65 years. In addition to genetics, some environmental factors may exert vital parts in PD occurrence as well. At present, more and more studies are conducted to elucidate the association between gut microbial dysbiosis and the incidence of PD. Gut microbial dysbiosis has a certain effect on both the central nervous system (CNS) and the enteric nervous system (ENS), which indicates that there is a gut-microbiota-brain axis that induces CNS disorders. Some gut microbial strains are suggested to suppress or weaken the neuroinflammation- and gut-inflammation-immune responses, which suggests the protective and pathogenic effects of certain gut microbial species on PD progression. Therefore, gut microbiome may contain plenty of targets for preventing and managing PD. Faecal microbiota transplantation (FMT) may serve as a direct and useful treatment for PD in the future. Nonetheless, there is little available scientific research in this field. The present work reviewed the latest research to examine the association of gut microbiota with PD, and the future prospects of FMT treatment.
Keywords
Introduction
Parkinson's disease (PD), a kind of multifocal and progressive neurodegenerative disorder, is mostly caused by dopaminergic neuron loss within substantia nigra pars compacta. PD shows the major symptoms of rigidity, tremor, bradykinesia (Samii et al., 2004) and numerous non-motor symptoms (NMS). Typically, NMSs more adversely affect the life quality of PD cases than motor symptoms (Martinez-Martin, 2011). In general, such NMSs are neuropsychiatric diseases (anxiety, depression, dementia, psychosis and apathy), and problems related to dysfunction of the enteric nervous system (ENS) and autonomic nervous system (ANS) (orthostatic hypotension, urinary changes and constipation). PD is a complicated disorder that involves multiple systems, which is associated with diverse mechanisms. The PD pathogenic mechanisms are mainly related to protein (α-synuclein) aggregation, endoplasmic reticulum stress (ERS), calcium homeostasis and mitochondrial dysfunction (Goswami et al., 2017; Sun and Shen, 2018), but its precise etiology remains to be further investigated.
Gastrointestinal (GI) dysfunction has been identified as a prominent contributing factor to the PD pathogenesis, typically, gut possibly provides a route for pathogens to spread into the central nervous system (CNS). Recently, the bi-directional communications taking place along the brain-gut axis are suggested to modify the neurobiological processes (Mayer et al., 2014; Sun and Shen, 2018). The linkages or interactions between brain and gut are achieved in different ways. To be specific, these may be attained by the influences of central inputs on the gut functions (like stress response), the impacts of gut-derived activity on the CNS function (GI disorder-related changes in cognition and moods), and the comparable roles of gut and brain in identical diseases (like neuronal degeneration). In traditional practice, the interactions and communications between gut and brain are regarded to be mostly regulated by inflammatory mediators and neuro-hormonal factors. Typically, it is suggested in latest studies that, gut microbiota is also a GI factor that affects the functions of brain and gut.
Lately, gut microbiota is extensively reported to affect the gut-brain axis, which also plays a certain role in the CNS-associated disorders and neuropsychiatric diseases, like multiple sclerosis (Jangi et al., 2016), autism (Strati et al., 2017), depression (Foster and McVey Neufeld, 2013; Kang and Cai, 2017a) and schizophrenia (Caso et al., 2016). Given the effects of gut microbiota and the microbial metabolites on the interference with the host behavior, cognition, immunity and metabolism (Marques et al., 2010; Cryan and O'Mahony, 2011; Foster and McVey Neufeld, 2013; P. Li et al., 2021), great attention has been paid to their impacts on the pathogenic mechanism of PD, and certain phenotypic associations are illustrated in recent years (Unger et al., 2016; Hill-Burns et al., 2017; Sun and Shen, 2018 ). For instance, the changes in the abundances or compositions of gut microbiota along with the microbial metabolites can be detected among PD cases. The gut microbial changes exert a vital part in PD induction and progression. Nonetheless, related data and scientific work are lacking at present. The emerging technologies make it possible to systemically investigate the gut microbiota and provide the practical data regarding the compositions and pathological changes (Kang et al., 2019). The present work aimed to review the latest studies concerning the association of gut microbiota with PD, together with the future prospects of the faecal microbiota transplantation (FMT) therapy.
The Microbiota-Gut-Brain Axis
As a bidirectional communication between GI tract and the CNS, the microbiota-gut-brain axis is associated with immune, endocrine mechanisms and neural pathways (Cryan and O'Mahony, 2011; De Palma et al., 2014; Sun and Shen, 2018; Dogra et al., 2021 ). It modulates the digestion events (such as secretion and motility), immunologic function, and perception or emotional response to visceral stimuli under normal condition (Mayer et al., 2006). Its dysfunction is related to several disorders (Cryan and O'Mahony, 2011; Aziz et al., 2013; Bonaz and Bernstein, 2013; Davari et al., 2013; Felice et al., 2016; Grenham et al., 2011; Mayer, 2011; Parashar and Udayabanu, 2017), like depression, anxiety, hepatopathy (Iwaki et al., 2021), intestinal inflammation, GI motility disorder, and chronic abdominal pain syndrome (Cryan and Dinan, 2012; Strati et al., 2021). Besides, this axis is linked with stress behaviors and responses (Clarke et al., 2013; O'Mahony et al., 2015; Parashar and Udayabanu, 2017). At present, it is suggested that microbiota involved in bidirectional communication that takes place along the brain-gut axis may serve as the significant contributor (Rhee et al., 2009; Bercik, 2011; Grenham et al., 2011; Mayer, 2011; Collins et al., 2012; Felice et al., 2016; Parashar and Udayabanu, 2017). Apart from the neuroimmune and neuroendocrine systems, this axis also involves the CNS, together with the parasympathetic and sympathetic branches of ENS and ANS (Grenham et al., 2011; Parashar and Udayabanu, 2017). The afferent fibers that travel from gut to cortical centers (including anterior/posterior cingulate, cerebral, amygdalar and insular cortices), together with the effector fibers that project to gut smooth muscles, stand for the main routes of bidirectional communications along the microbiota-gut-brain axis (O'Mahony et al., 2011; Parashar and Udayabanu, 2017). The complicated network may possibly affect the GI and cerebral functions (Rhee et al., 2009; Mayer, 2011; Felice et al., 2016).
Roles of Gut Microbial Dysbiosis in PD
Increasing studies prove that PD is linked with gut microbial dysbiosis in both human beings and animals. At the same time, some mechanisms may be related to the effects of microbial alterations on the gut-brain interaction in PD. Figure 1 presents the mechanisms of gut microbial dysbiosis in affecting PD occurrence.
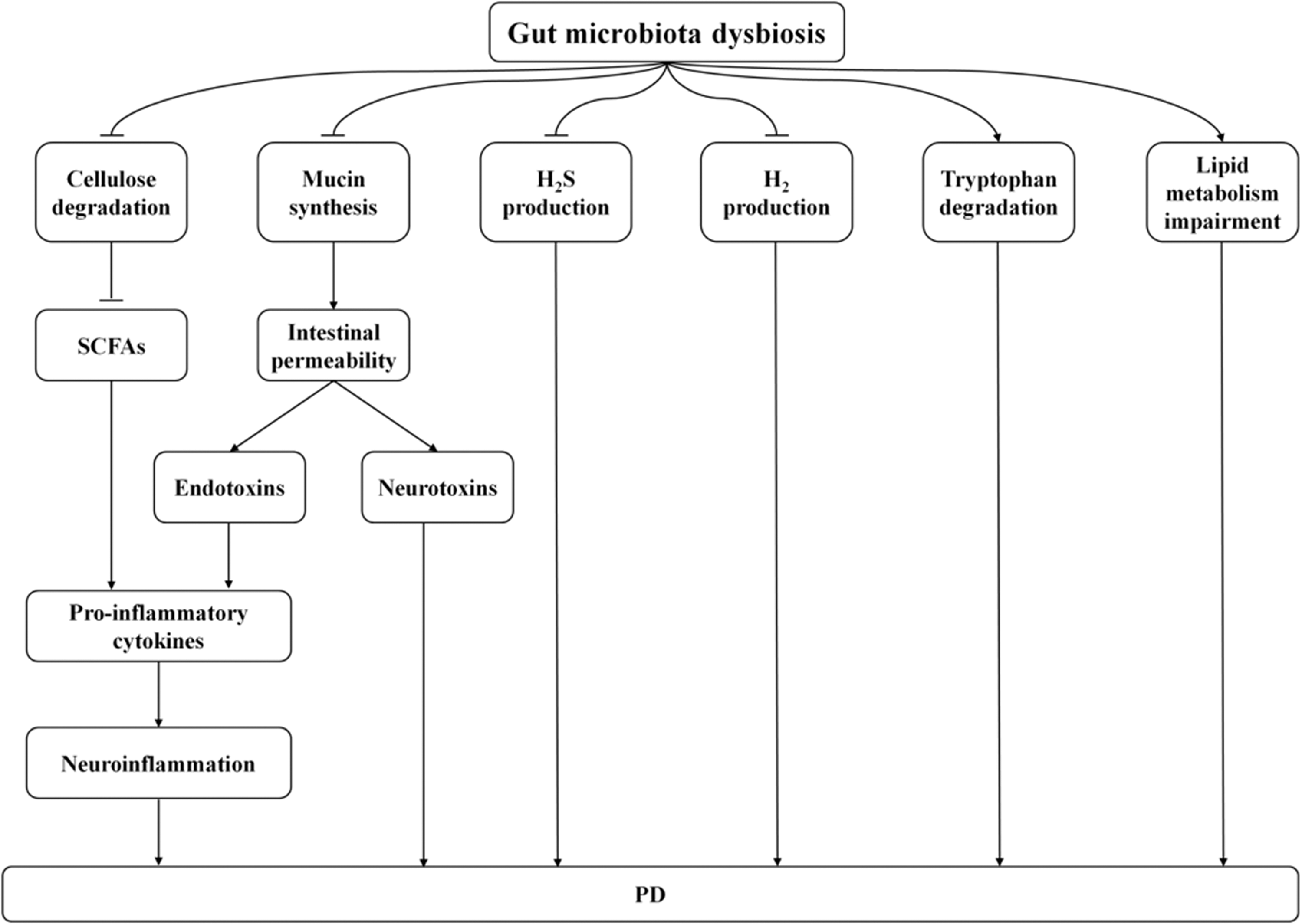
Possible Mechanisms Associated With the Relationship Between Intestinal Microbiota and PD. The ability of intestinal flora to degrade cellulose to synthesize SCFAs may reduce, leading to increased production of pro-inflammatory factors and ultimately inducing neuroinflammation. The gut microbiota may contribute to increasing intestinal permeability via decreasing the epithelial mucin proteins, which results in translocation of endotoxins and neurotoxins across the intestinal wall. In addition, endotoxins may induce production of pro-inflammatory cytokines, which eventually lead to neuroinflammation. Intestinal microbiota may reduce H2S production, H2 production, and increase tryptophan degradation. Gut microbiota may lead to the impairment of lipid metabolism. These mechanisms may lead to impaired CNS function, such as alterations in cognition, behavior and stress responses, finally causing or aggravating PD.
Experimental Studies
As suggested by animal studies, gut microbial dysbiosis can be detected in various animal models of PD. Most of these works indicate that, compared with normal controls, PD patients display changes in the relative abundances of potentially “harmful” and “beneficial” bacteria (Table 1). Research on germ-free (GF) mice first suggests that gut microbiota is related to PD. Sampson et al. (2016) found that specific pathogen-free (SPF)-Thy1-αSyn (alpha-synuclein-overexpressing [ASO]) animals suffered from progressively declined motor function, while such phenomenon was markedly postponed among the Germ-free (GF)-ASO animals. Besides, the reduced abundances of order Clostridiales and phylum Firmicutes, as well as the elevated abundances of orders Enterobacteriales and Turicibacterales and phylum Proteobacteria are recently detected in fecal samples collected from mice with PD induced by 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP)(Sun et al., 2018).
Changes in Microbiota Composition Associated With PD in Animals.

In line with the research, rotenone-induced PD mice also display fecal microbial dysbiosis, which is featured by the significantly changed microbial compositions (particularly the elevated Firmicutes-to-Bacteroidetes ratio) and the decreased bacterial diversity. Additionally, the rotenone-induced motor and GI impairments are suggested to markedly relate to the alterations of fecal microbial composition (Yang et al., 2017). Further, as discovered from Sprague-Dawley (SD) rats exposed to rotenone treatment the relative abundances of Lactobacillus, Bifidobacterium, Sutterella and Turicibacter increased in colon and small intestine; by contrast, those of Oscillospira, Prevotella and S24-7 declined (Johnson et al., 2018). According to histological analysis, rats exposed to rotenone treatment show thickened colonic mucosa as well as goblet cell hyperplasia, and this is possibly an adaptive response to GI microbial changes. In addition, Bhattarai et al. (2021) demonstrated that chronic rotenone administration by oral gavage resulted in the loss of tyrosine hydroxylase (TH) neurons in both GF and conventionally raised (CR) mice, the decrease in motor strength and coordination was observed only in CR mice. Chronic rotenone treatment did not disrupt intestinal permeability in GF mice, but it resulted in a significant change in gut microbiota composition and an increase in intestinal permeability in CR mice. These results suggested the potential role of gut microbiota in regulating barrier dysfunction and motor deficits in PD.
Collectively, these findings obtained from diverse animal samples reveal the role of gut microbial dysbiosis in the pathogenic mechanism of PD. But, a lot of research is still needed to determine the specific strains of bacteria that can treat PD and the specific strains that can cause PD. Meanwhile, further study need to emphasize more on the metabolomic aspect and its role in pathophysiology and the nutritional aspect.
Clinical Studies
Increasing evidence has suggested that gut microbial dysbiosis is also found in PD patients (Table 2). Meanwhile, gut microbiota plays a certain role in the pathophysiology of PD. It is previously suggested that, bacterial overgrowth in the small intestine is prevalent among PD cases, which is related to poor motor function (Tan et al., 2014). Hopfner et al. (2017) found that the Beta-diversity was significantly different between PD cases and controls. Meanwhile, Lactobacillaceae, Barnesiellaceae and Enterococcacea markedly increased in PD cases compared with those in controls. In addition, Keshavarzian et al. (2015) found that compared with control subjects, the fecal and mucosal microorganisms showed significant difference among PD cases. In the feces of PD patients, the relative abundances of “anti-inflammatory” butyrate-producing bacteria from genera Blautia, Coprococcus, and Roseburia significantly decreased compared with those in controls. Typically, the relative abundance of genus Faecalibacterium markedly decreased in mucosal samples among PD cases compared with those in controls. However, the abundance of “proinflammatory” Proteobacteria of genus Ralstonia significantly increased compared with that in controls. In addition, as revealed by predictive metagenomics, many metabolic genes were markedly down-regulated in fecal microbiota among PD cases relative to normal controls, while genes related to lipopolysaccharide (LPS) biosynthesis and the type III bacterial secretion systems were evidently up-regulated among PD cases relative to normal subjects. Based on the above findings, proinflammatory dysbiosis possibly existed among PD cases, which might promote the inflammation-mediated α-Syn misfolding and PD progression.
Changes in Microbiota Composition Associated With PD in Humans.

In line with these results, Hasegawa et al. (2015) showed that the sum of fecal bacterial counts markedly decreased in PD patients relative to controls. Compared with normal controls, the abundance of Lactobacillus remarkably increased, whereas those of Prevotella, Bacteroides fragilis, Clostridium coccoides, together with bacteria that produced hydrogen significantly declined in fecal samples from PD cases. Additionally, the course of disease was predicted using the linear regression model, which revealed that the Lactobacillus gasseri subgroup and the C. coccoides group were significantly positively and negatively correlated with coefficients, separately. Meanwhile, the serum levels of lipopolysaccharide (LPS)-binding proteins markedly reduced in PD patients compared with controls. All in all, these data suggest that gut microbiota dysbiosis may result in reduced mucin production but elevated intestinal permeability, thus affecting the development and progression of PD. As suggested in a clinical study, the Prevotellaceae level decreased by 77.6% in fecal samples from PD cases in relative to normal controls (Scheperjans et al., 2015). It is interesting that, the Prevotella-produced hydrogen sulfide protects the dopaminergic neurons in mouse and rat models with PD (Hu et al., 2010; Kida et al., 2011; Cakmak, 2015). In addition, the abundance of Enterobacteriaceae was significantly higher among cases developing postural instability and gait difficulty (PIGD) than those with having tremor dominant (TD). In the meantime, the Enterobacteriaceae level showed positive correlation with PIGD severity. To sum up, the above findings indicated the relationship of at least one change in certain bacteria with PD pathology or symptoms (Scheperjans et al., 2015).
It is also discovered in another work that, the abundances of Bacteroidetes, Prevotellaceae, Lactobacillaceae, Enterococcaceae and Faecalibacterium prausnitzii evidently decrease in the fecal samples from PD cases in comparison with normal subjects, while those of Enterobacteriaceae and Bifidobacterium dramatically increase in feces from PD cases relative to normal subjects (Unger et al., 2016). At the same time, the fecal contents of short-chain fatty acids (SCFAs) in feces markedly decreased among PD cases compared with normal controls. Taken together, the above findings indicate that gut microbiota dysbiosis possibly results in reduced SCFAs contents, thereby inducing changes in ENS and facilitating GI dysmotility in the context of PD (Unger et al., 2016). It is also suggested that, the levels of unclassified Firmicutes and Verrucomicrobiaceae (Akkermansia muciniphila) apparently increase, while those of Erysipelotrichaceae (Eubacterium biforme) and Prevotellaceae (Prevotella copri) evidently decrease among PD samples relative to normal subjects (Bedarf et al., 2017). Furthermore, the microbiota metabolism that involves the β-glucuronate degrading pathways significantly decreases, while the microbiota metabolism involving the tryptophan degrading pathways significantly increases in PD (Bedarf et al., 2017). Altogether, these results suggest that gut microbiota possibly has a certain effect on PD occurrence through destroying the gut barrier function while aggravating intestinal inflammation (Bedarf et al., 2017).
As suggested in more clinical studies, fecal samples from PD show high abundances of Akkermansia, Bifidobacterium, Ruminococcaceae, and Lactobacillus, whereas decreased abundance of Lachnospiraceae. Additionally, functional predictions reveal the increased activity of the degrading xenobiotics pathways in PD (Hill-Burns et al., 2017). Also, compared with normal subjects, PD samples display reduced cellulose-decomposing bacterial species of genera Blautia, Ruminococcus and Faecalibacterium, whereas the pathogenic bacteria of genera Streptococcus, Escherichia-Shigella, Enterococcus and Proteus evidently increase among PD cases in comparison with normal subjects. Additionally, the course and severity of PD show negative correlation with cellulose decomposing bacteria, whereas positive correlation with pathogenic bacteria. These findings indicate that, the gut microbial structural alterations among PD patients possibly decrease SCFAs secretion, but increase the production of neutrotoxin and endotoxin. Such alterations may be related to PD pathological development (W. Li et al., 2017). It is reported in one work that, the abundances of genera Aquabacterium, Clostridium IV, Anaerotruncus, Sphingomonas, Holdemania, Butyricicoccus and Clostridium XVIII increase, but those of genera Sediminibacterium and Lactobacillus decrease among PD cases in comparison with normal subjects. Additionally, genera Clostridium XlVb and Butyricicoccus are related to the non-motor symptom (NMS) cognitive impairment (Qian et al., 2018). Particularly, changes in gut microbiota exert a vital part among PD cases. In line with the research, by characterizing the gut microbial compositions of 63 PD patients, 63 healthy spouses (HS) and 74 healthy people (HP), Zhang et al. (2020) found that microbial compositions were significantly different among PD, HS and HP samples in terms of alpha/beta diversity. Meanwhile, microbial compositions were also different in Hoehn & Yahr stage and disease duration. Eight inflammation-associated microbial genera (Parabacteroides, Akkermansia, Coprococcus, Bilophila, Collinsella, Methanobrevibacter, Eggerthella, Adlercreutzia) shared a continuously increasing trend with the increases in Hoehn & Yahr stage and disease duration, indicating characteristic bacteria associated with deterioration in PD. Additionally, seven bacterial markers were identified for accurately differentiating PD patients from the controls. Recently, a meta-analysis finds that these PD-related gut microbiota dysbiosis may lead to the impairment of SCFAs producing process, lipid metabolism, immunoregulatory function, and intestinal permeability, which contribute to the pathogenesis of PD (Shen et al., 2021). Vascellari et al. (2020) analyzed the gut microbial composition of 64 Italian PD patients and 51 controls. Their results discovered that the most significant changes in the PD group highlighted a reduction in bacterial taxa, which were linked to anti-inflammatory/neuroprotective effects, particularly in the Lachnospiraceae family and key members, such as Butyrivibrio, Pseudobutyrivibrio, Coprococcus, and Blautia. In addition, lipids (linoleic acid, oleic acid, succinic acid, and sebacic acid), vitamins (pantothenic acid and nicotinic acid), amino acids (isoleucine, leucine, phenylalanine, glutamic acid, and pyroglutamic acid) and other organic compounds (cadaverine, ethanolamine, and hydroxy propionic acid) changed in the fecal metabolites. Most modified metabolites were strongly correlated with the abundance of members belonging to the Lachnospiraceae family.
All in all, these observations shed more lights on gut microbial dysbiosis among PD cases, which may be used to guide disease interventions through gut microbiota manipulation. However, how does the gut microbiota change during the course of PD remains unclear. Moreover, it remains unclear if different microbial compositions are related to different phenotypes of PD. Further research is needed to determine which bacteria and metabolites can serve as the biomarkers for accurately differentiating PD patients from the controls. In addition, PD-associated metagenomics studies should consider the family environmental factor.
Future Prospects of FMT Therapy for PD
The FMT therapy refers to the injection of fecal suspension obtained from one healthy subject to the GI tract of another subject for the treatment of a certain disorder. FMT is a direct and superior treatment for PD through the manipulation of human gut microbiota. The FMT therapy has been applied for more than 50 years. Typically, Ge Hong in China first described to use feces as a treatment for various disorders, like diarrhea, in the 14th century (Zhang et al., 2012). In 1958, Eiseman et al. used fecal enemas to treat pseudomembranous colitis (possibly induced by Clostridium difficile infection [CDI]), which was the beginning of introducing FMT in the mainstream medicine (Eiseman et al., 1958). Since then, FMT has become increasingly popular because it is effective and easy to use to treat CDI. In recent years, FMT is increasingly investigated. It plays a certain role in a number of disorders, including irritable bowel syndrome, inflammatory bowel disease (IBD), anorexia nervosa, metabolic diseases, multiple sclerosis, autoimmune diseases, neuropsychiatric disorders, cancers, and cardiovascular diseases (CVDs)(Di et al., 2013; Smits et al., 2013; Vrieze et al., 2013; Kang et al., 2016, Kang et al., 2017; Kang and Cai, 2017b, 2018a, 2018b, 2018c, 2019a). Similarly, FMT is thus used as a treatment for PD. Some recent studies have evaluated the feasibility of changing gut microbiome for treating PD (Table 3). The candidate mechanisms of FMT underlying these situations are manipulation of gut microbial composition, gut barrier fortification, pathogen suppression, and immunomodulation (Figure 2). For instance, Sun and coworkers (Sun et al., 2018) carried out research on FMT, where fecal microbiota from the normal C57BL/6J donors was transplanted into the PD C57BL/6J recipients. FMT therapy mitigated gut microbial dysbiosis mainly by increasing the abundances of Firmicutes and Clostridiales, while reducing those of Proteobacteria, Turicibacterales and Enterobacteriales. Meanwhile, FMT decreased the production of fecal SCFAs, mitigated physical dysfunction, and boosted the levels of striatal serotonin (5-HT) and dopamine (DA) in PD mice. Furthermore, FMT decreased the activation of astrocytes and microglia in substantia nigra, and down-regulated the components in the Toll-like receptor 4 (TLR4)/TNF-α signal transduction pathway in the brain and gut. The above findngs revealed the effect of FMT on protecting PD mice through inhibiting neuroinflammation while decreasing the TLR4/TNF-α signal transduction. In line with the observation, Sampson et al. (2016) found that specific pathogen-free (SPF)-Thy1-αSyn (alpha-synuclein-overexpressing [ASO]) animals exhibited progressively declined motor function, while this markedly postponed among the Germ-free (GF)-ASO animals. It is remarkable that, colonizing the mice with αSyn-over-expression with microbiota obtained from PD cases aggravates physical dysfunction in comparison with microbial transplants obtained from the normal subjects. The above results suggest that gut microbiota modulates the mouse motion disorders and the changes in human microbiome may serve as a risk factor for PD.
Changes in Microbiota Composition Associated With PD and FMT Therapeutic Strategies.

NO refers to no test or no research.

Possible Mechanisms Underlying the Relationship Between Fecal Microbiota Transplantation (FMT) and PD Are as Follows: FMT May Inhibit Neuroinflammation by Decreasing the TLR4/TNF-a Signal Transduction. FMT may decrease astrocytes and microglia activation by reducing production of SCFAs, and ultimately inhibits neuroinflammation. FMT may boost serotonin (5-HT) and dopamine (DA) levels.
Concluding Remarks and Perspectives
However, FMT therapy is a new and under-explored therapy for PD. The available scientific information in this field is lacking. Therefore, more efforts should be made to better understand the role of FMT in PD. First of all, a causal relation between alteration of gut microbiota and PD need to be proven. Application of fecal transplantation in PD disease may be regarded as an interesting research tool to identify the causally involved intestinal microbial species and/or non-microbial components as initiators of both gastrointestinal and systemic autoimmune reactions. Therefore, we should devote more effort to studying FMT therapy to PD. Secondly, it is worthwhile to find out how FMT re-establishes a balanced intestinal microbiota and results in impressive cure rates in patients with PD disease. Thirdly, more randomized controlled trials (RCTs) with reasonable design should be carried out to determine whether FMT is safe and effective on PD. Additional high quality data (e.g. longitudinal study) are also needed to analyze the potential adverse effects. Furthermore, it is important to analyze the fecal and small intestinal microbial compositions prior to and following FMT. The above studies shed more lights on the mechanism of FMT and help to discover the microbes together with the metabolites related to PD pathogenesis. Fourthly, the effects of microbial metabolites on PD should be further investigated. Particularly, it should be further investigated about whether SCFAs protect or promote PD. Last but not least, FMT is a simple treatment, which only requires a short operation time. Compared with repeated conventional treatments, the hospitalization cost of FMT is low. Thus, the standardization of FMT protocols is urgently established. For this reason, in addition to conventional approaches, FMT is promising as an alternative therapy for PD diseases in the future.
Footnotes
Author Contributions
Yongbo Kang collected and wrote most of the experiments with help from Xing Kang, Hongfang Zhang, Qingqing Liu, Hao Yang and Weiping Fan.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by Science Research Start-up Fund for Doctor of Shanxi Medical University (Grant No.XD1807), Science Research Start-up Fund for Doctor of Shanxi Province (Grant No.SD1807), Scientific and Technological Innovation Programs of Higher Education Institutions in Shanxi (Grant No. 2019L0425) and Shanxi Province Science Foundation for Youths (Grant No. 201901D211314).