
Abstract
Blueberry fruits were located in full shade (complete darkness), partial shade (16% full sun) and light exposed (100% full sun) conditions. An inversely proportional relationship between the growth rate of the fruit and its relative water content was clear in fully shaded fruits, with an increase in the shaded fruits transpiration rate during ripening having been recorded. The pedicels of light-exposed fruits showed greater development of phloem tissue than those of fully shaded fruits. The structure of the stomata was modified in the fully shaded fruits, with a decrease in the size of the subsidiary cells. The epidermis of exposed and partially shaded fruits presented a profuse layer of epicuticular wax at maturity, which was almost absent in fully shaded fruits. Throughout blueberry fruit development, the deficiency of solar radiation induced anatomical changes that, in turn, affected water economy.
Introduction
Growth stages of blueberry fruit are associated with distinct biological processes. During these periods, water and carbon accumulate in the fruit, determining quality traits such as fruit size and sugar concentration. 1 Blueberry fruit growth exhibits a double-sigmoid pattern, which is mainly defined in late ripening cultivars. 2 During the first stage of fruit growth, Birkhold. 3 estimated a large contribution of carbon from fruit photosynthesis, through CO2 assimilation in the exocarp. The comparatively high surface area: volumen ratio and a relatively high stomatal density improve CO2 and water exchange. Hypodermal and parenchymal cells of blueberry fruit contained numerous chloroplasts,4,5 however the sepals remained until maturity, contributing with chlorophyll and the majority of fruit stomata. 6 During the second sigmoidal stage, fruit photosynthesis also play a relevant role in blueberry fruit growth, 7 likely through CO2 uptake by the sepals 6 and captured and re-fixed in the mesocarp, as was determined in other fleshy fruits,8–13 so the gas and water vapor exchange through the stomata has a minor role.
It can be considered that, in general terms, fruits are architecturally isolated, and they often have low transpiration and xylem transport rates when compared with the rest of the plant 14 In fleshy fruits, both xylem and phloem tissues participate in delivering water, depending upon the developmental stage of the fruit. 15 It has been determined that pedicel xylem hydraulic conductance in some crops decreases at later ripening stages.16–18 A developmental switch to phloem water delivery from predominantly xylem driven delivery reduces the direct hydraulic link of fruit water status to that of the plant. 19 In grape, Thomas et al. 20 observed that during normal fruit development a decrease in mesocarp turgor coincides with the onset of veraison. A shift of phloem unloading from symplasmic to apoplasmic pathway is involved in developmental onset of ripening in grape berry. 21 Constantinescu et al. 22 model predicted a recirculation of water from the fruit phloem to the fruit apoplast during the middle part of the day, a regulated uptake of sugar into the fruit, and the prevention of water loss due to xylem backflow. Apoplastic space increases in ripening blueberries, in this sense Konarska 4 observed loose packing of cells resulting in large intercellular spaces between groups of cells.
The accumulation of calcium and its distribution in the fruit is highly dependent on the water supply and interactions with the cell wall in the apoplast. Being an immobile element in the phloem, the accumulation of calcium inside the fruit depends mainly on the transpiration flow. The link between the transport of water and calcium is particularly visible when observing target organs with relatively low transpiration rates, such as those that generally occur in fruits. 14
Vaccinium spp. are hypostomatous plants because stomata were not observed on the adaxial side of leaves. The stomata density and stomata area varied considerably among different blueberry genotypes. Overall, northern highbush blueberry genotypes exhibited more stomata than southern highbush blueberry genotypes. 23 Northern highbush blueberry exhibits higher leaf photosynthetic rates than that of southern highbush blueberry, but both blueberry types exhibit lower photosynthetic rates than those of other fruit crops. 24
The stomata present in the epidermis of the fruit, when functional, exchange water vapor with the environment. Fruit transpiration rate and fruit water content are influenced by the stage of fruit development, the evapotranspiratory demand and the effect of light intensity on stomatal opening. Functional stomata open in response to increasing light intensity whilst closure is brought about by reductions in light intensity. 25
Leaves adapted to different irradiances are found within the canopy. 26 In response to changes in light availability, shade leaves increased the specific leaf area (projected leaf area per unit leaf dry mass), had lower leaf thickness, less volume of the palisade parenchyma and spongy parenchyma, less stomatal frequency and stomatal area per unit leaf area, and had reduced xylem conduit number in petioles than sun leaves.27–32
Assuming that the fruit carpels are modified leaves, 33 the deficiency of solar radiation could induce anatomical changes that, in turn, would have implications for the physiology of the fruit. There are few references linking irradiance deficiency with anatomical changes in fleshy fruits. In kiwi (Actinidia chinensis var. deliciosa), shading negatively affected the differentiation of the vascular system, in particular the xylary component, 34 with implications for transpiration flow. In nectarine (Prunus persica var. nucipersica), shading negatively affected the structure of the accompanying phloem cells. 35 Such anatomical modification would influence the phloem discharge and would result in a low accumulation of soluble solids in the parenchymatic cells of the fruit.
It is known that light exposure is strongly associated with fruit temperature.36–38 The methodology used in the present work was aimed at assessing the direct effect of light, independent of the effect of temperature, on the anatomical features of blueberry fruits. It was hypothesized that local shading would induce anatomical changes in the highbush blueberry fruit that modify the transpiratory rate and the water economy of the fruit.
Methods and materials
Experimental site and fruit treatments
The trial was carried out in a commercial orchard of highbush blueberry (Vaccinium corymbosum) cvs. Brigitta (northern highbush blueberry) and OzarkBlue (southern highbush blueberry) planted in Sierra de los Padres (37°55’48’'S; 57°48’15’’W), South-East Pampas region, Argentina.
Soil was a Typic Argiudoll, with pH 6.6, organic matter 6.2%, extractable potassium 425 mg kg−1, and phosphorus 15 mg Kg−1 (P-Bray) at the upper horizon. Previously to plantation, soil was amended with pine bark and rice hulls. Plants were set on raised beds, with a black plastic mulch and pine needles around the bushes during the first three years, later the plastic mulch was replaced with wheat straw. The orchard alley had a sod cover. Bushes were fertigated during spring with ammonium sulphate (AgroSuma S.R.L, Buenos Aires, Argentina). Drip irrigation was applied daily in the summer according to tensiometers placed at 20 and 30 cm depth. Soil moisture tension was kept within between 15–25 kPa. To correct alkalinity (pH = 7.8) of irrigation water, sulfuric acid was added to drop pH of water to 5.0–5.5. Commercial production started at the 3rd year after planting. Thereafter, plants were pruned annually during winter in order to achieve an open center. At the time of the experiments (2021–2022) plants were 6th – 7th years old. In 2021, the area had an oceanic climate typical of the temperate south-east Pampas region, with warm and humid summers, reaching an average maximum temperature of 27°C and an average minimum temperature of 14°C in January (cool nights). The climatic conditions in 2022 were characterized by a hot and dry summer. Maximum temperatures exceeding 30°C were frequently recorded during January 2022.
Fruits were exposed to different light exposures regardless of the rest of the plant. Three treatments were evaluated:
Local full shade treatment. A fruit cluster-shield developed by Godoy et al.
7
was used over the whole cycle of fruit development, i.e., from fruit set to harvest (total fruit shading). The device consisted of a double-layer cover, the inner of aluminium foil, and the outer of polypropylene film. The last one was used to counteract the effect of shading on fruit temperature. Temperature measurements were taken using thermocouple sensors placed next to the sampled bunches, connected to a data logger (Cavadevices, Buenos Aires, Argentina). Regression analysis was performed to check if the air temperatures inside the shield and the air temperatures next to exposed fruit were similar throughout the fruit development period. A close relationship (R2 = 0.93) was found between both treatments. Local partial shade treatment. An 80% shading net (high-density polyethylene monofilament, AgriNet, Mendoza, Argentina) covering the fruit cluster during the whole cycle of fruit development (partial fruit shading). Due to the folding to attach it to the bunch peduncle, the net intercepted on average 84 ± 1% of the photosynthetic flux corresponding to the control (16% full sun). Data were obtained using quantum radiation sensors coupled to a LiCor Li-1000 data logger (Lincoln, NE, USA). Control treatment. Sun-exposed clusters located at the outer and middle areas of the bush was randomly selected. A completely randomized design with six replications (bushes) per treatment was used.
Statistical analysis of the obtained data was performed using ANOVA followed by Tukey's multiple comparisons test.
Optical microscope images of fruit pedicels
Pedicels were dehydrated in a graded etanol-xylene series, embedding in paraffin and transversally sectioned with a Minot microtome. After deparaffinization in xilene, the pedicel sections were transferred to a glass slide, and were stained with safranin and fast green. The images observed under an optical binocular microscope (Olympus SZX12) at 40 x were captured by a digital camera (Olympus America Inc., OLY-200 Color Video Cámara w/ U-PMTVC) and processed using the Image J program.
Optical microscope images of fruit epidermal imprints
Fruits were superficially cleaned with a cotton cloth. Subsequently, a layer of transparent solvent-based varnish (ethyl acetate) was applied with a brush. Once dry, the imprint was separated from the fruit using transparent adhesive tape, which was placed on a slide. The images observed under a binocular optical microscope (Olympus SZX12) at 40X were captured by a digital camera (Olympus America Inc., OLY-200 Color Video Cámara w/ U-PMTVC).
SEM images of fruit epidermis
Fruits were dehydrated in a graded ethanol-xylene series, embedding in paraffin, and tangentially sectioned with a Minot microtome. After deparaffinization in xylene, the fruit sections were fixed on aluminium stubs by using double-sided adhesive conducting carbon tape, and were sputter coated with approximately 4 nm gold-palladium in a Denton Desk II sputter coating unit (Denton Vacuum, NJ, USA). Observations were carried out on a Jeol JSM 6460 - LV scanning electron microscope using a secondary electron detector with an accelerating voltage of 15 kV at high vacuum (HV) mode.
Fruit relative water content
The relative water content (RWC) of the fruits of each cluster was calculated according to the following equation:

where:
RWC: relative water content; W: fruit fresh weight of each cluster; Wsat: fruit saturated weight. In order to achieve maximum hydration of the fruits, the clusters, once separated from the plant, were immediately placed in numbered plastic tubes filled with distilled water, ensuring that the peduncles were submerged. The plastic tubes containing the clusters, 3 for each treatment, were placed in a rack. The fruits of each cluster, after 24 h of hydration in the dark (until constant weight of the cluster), were weighted in a precision balance accurate to 0.01 g. DW: fruit dry weight. Fruits of each cluster were weighted after having remained in an oven at 60°C until constant weight.
Three clusters / treatment per variety and per day were measured weekly.
Estimation of the fruit transpiration efficiency
An adaptation of the potometer method was developed for estimation of fruit transpiration rate. The clusters, once separated from the plant, were immediately placed in numbered plastic tubes filled with distilled water, ensuring that the peduncles were submerged. The plastic tubes containing the clusters, 6 for each treatment, were placed in a rack. The clusters, after 24 h of hydration in the dark, were taken to a culture chamber, previously leveling the tubes, exposing them to a PPFD of 140 µmol µmoles m−2 s−1, a 16 / 8 h of photoperiod (day/night) and 24 ± 0.5°C for 24 h. The difference in volume of water in each tube was measured. Fruits were separated from the rachis, the diameter of each fruit was measured, its surface was estimated assuming a spherical shape, and the total fruit surface was calculated. The mean transpiration rate was estimated as the volume of water consumed per cm2 of fruit surface by day.
Determination of calcium content
Samples of 30 ripe blueberry fruits corresponding to each treatment were dehydrated in an oven (65–70°C), weighed and calcined in a muffle at 550°C. They were subsequently digested with concentrated nitric/perchloric acid (1/5, v/v) (Suprapure, Merck, Darmstadt, Germany), previously ultra-purified (Berghof Distillacid BSB-939-IR, GmbH Germany), with a microwave digestion system (Mars-5, CEM Corporation, USA). The analysis was carried out with an inductively coupled plasma optical emission spectrometer (ICP-OES) equipped with a cross-flow nebulizer (ICPE-9000, Shimadzu Corporation, Kyoto, Japan). A Milli Q-M device was used to produce high purity water used in calibration.
Results and discussion
Changes in fruit anatomy due to cluster shading
Vascularization of the pedicel
The xylem and phloem (excluding the sclerenchyma fibers layer) were identified and the corresponding surfaces were determined in each histological section (Figure 1). No significant differences were found (p = 0.688) between the surface of the pedicellar xylem of shaded (0.034 mm2) and exposed (0.033 mm2) fruits. On the other hand, the pedicels of exposed fruits showed a phloem surface (0.025 mm2) that was significantly higher (p = 0.035) than that of shaded fruits (0.021 mm2).
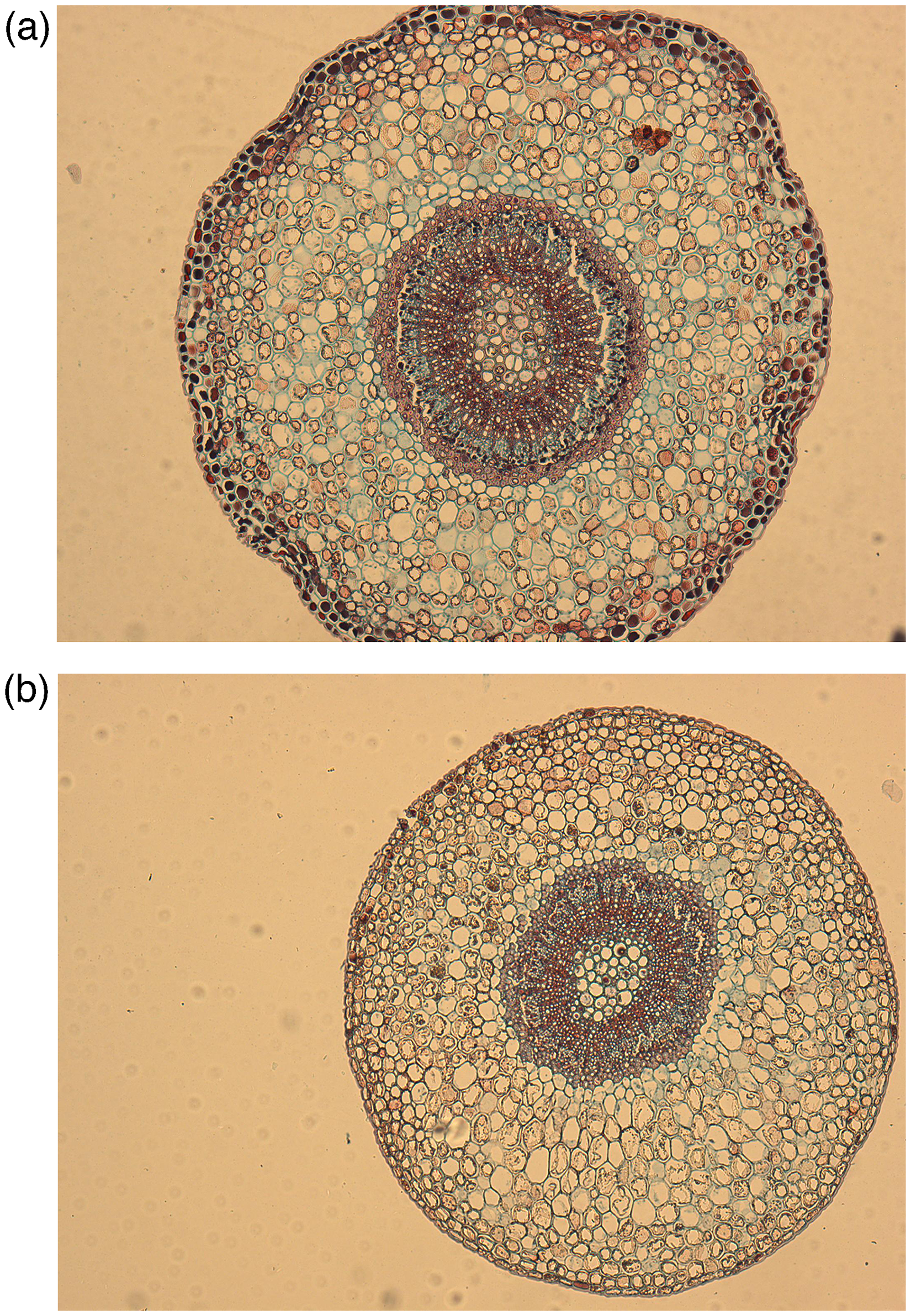
Pedicel cross-section corresponding to ripe blueberry fruits that were developed under (a) sunlight exposure and (b) total shading. References: epidermis (e), cortical parenchyma (cp), sclerenchyma fibers (sf), secondary xylem (sx), medullary parenchyma (mp), secondary phloem (sp).
Pedicels of exposed fruits presented a greater amount of phloem tissue than those of shaded fruits, with no differences being observed in the xylem tissue. Generally, the hydraulic conductance of the pedicel xylem decreases in the late stages of fruit development,16–18 coinciding with a modification of the pathway of hydraulic flow, which goes from xylem transport to fundamentally phloem transport. 19 This change is due to the increase in fruit demand, and is related to a change towards a phloemmatic discharge of solutes 15 and water 22 in the fruit apoplast. 21 Recently, in cranberry (Vaccinium macrocarpon) Rojas-Barros et al. 39 found a gradual reduction in pedicel xylem radius, both proportionally and absolutely, due to an increase in the phloem radius and pith during fruit development.
In the case of blueberries exposed to sunlight, whose tissues photosynthesize even in the second stage of growth, the greater synthesis of carbohydrates would cause an increased water supply to the fruit, 22 which would be accompanied by a greater participation of pedicellar phloem transport and a concomitant increase in the proportion of phloem tissue.
Stomata
Stomatal density in the calycinal zone, where stomata are gathered in mature fruits, did not differ significantly (p = 0.766) between fully shaded fruits (36.9 ± 1.1 stomata/mm2) and exposed fruits (38.8 ± 1 stomata/mm2). It is likely that when shading was imposed the total number of stomata was almost defined. Cano-Medrano and Darnell 40 found that cell division in blueberry (V. ashei) fruits ceased early in their development. Yang et al. 41 determined that stomatal density was greater at the beginning of fruit development. In developed blueberry fruit, stomata are scarce and concentrated primarily near the calyx. 5 In blueberry, fruit stomatal conductance is 3-fold lower during later stages compared to that at earlier stages of fruit development (petal fall). 42
The surface of the stomatal guard cells (250 ± 2 µm2 shaded fruits; 256 ± 4 µm2 exposed fruits) did not differ significantly between treatments (p = 0.696), however, the surface of the subsidiary stomatal cells (Figure 2) in the shaded fruits was significantly lower (p = 0.005) than in the exposed fruits (712 ± 8 µm2 vs. 890 ± 13 µm2).
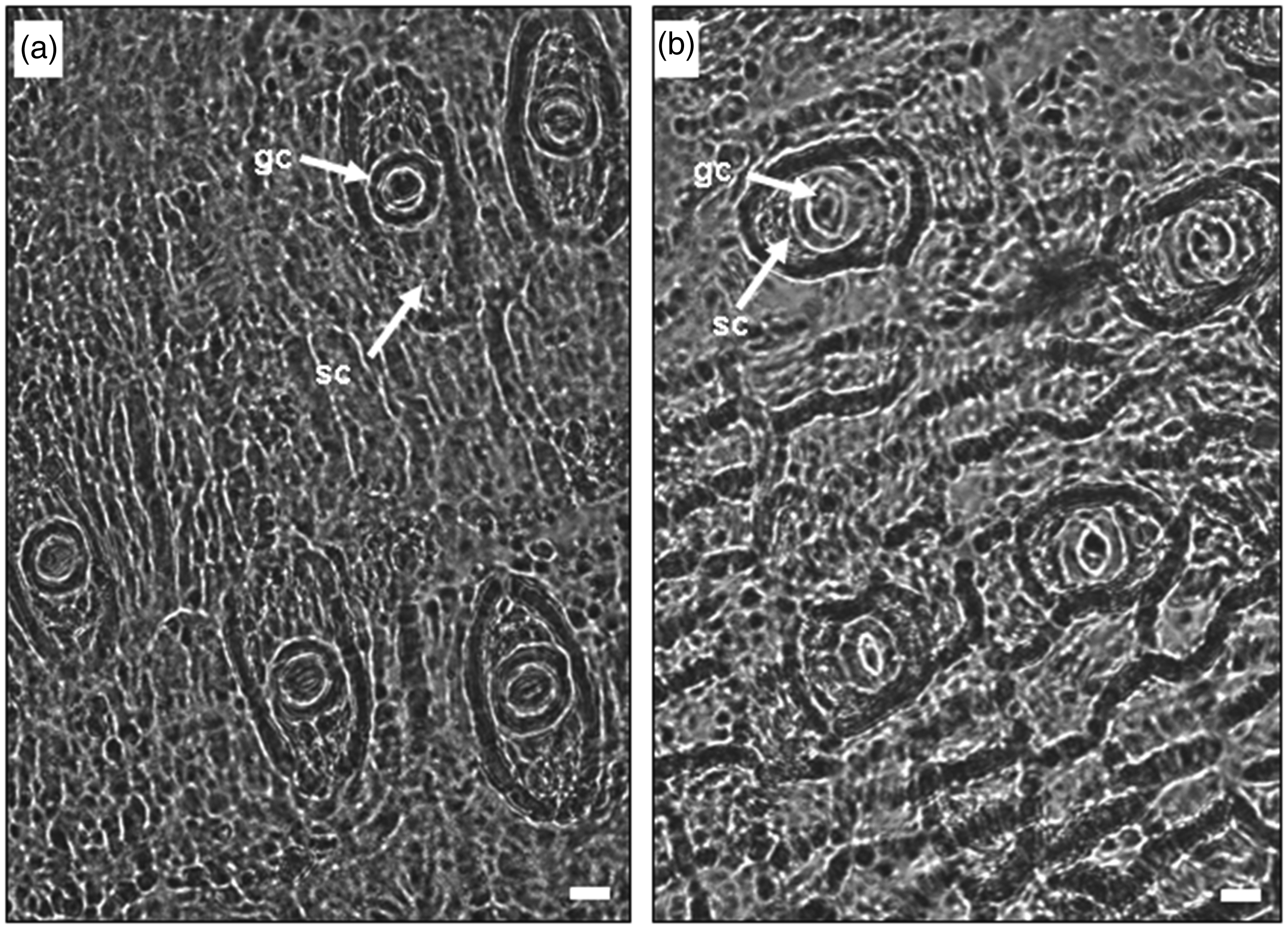
Epidermal imprints of ripe ‘Ozarkblue’ blueberry fruits (400x) that were developed under (a) sunlight exposure and (b) total shading. References: guard cell (gc), subsidiary cell (sc). Bar: 10 µm.
Paracytic stomata, also called rubiaceous, are characteristic of the tribe Vaccinieae, to which the genus Vaccinium belongs. 43 These stomata have two subsidiary cells parallel to the long axis of the guard cells (Figure 3). In general, the morphological arrangement of subsidiary cells in angiosperms affects the mechanical properties of stomata. 44

Image of epidermis of ripe blueberry ‘Brigitta’ fruit subjected to total shading obtained by scanning electron microscopy (600 X). The paracytic stomata are observed with open occlusive cells and the corresponding outer subsidiary cells.
In species with paracytic stomata, Franks and Farquhar 45 showed that lateral movement of subsidiary cells favored stomatal opening, probably due to changes in osmotic potential. 44 In their natural habitat, blueberries grow as understory plants in low to moderate light environments and diffuse light. 46
Therefore, it would be expected that blueberries have developed mechanisms to respond quickly to abrupt changes in radiation availability, caused, for example, by shading by their own leaves or those of adjacent plants. However, McAusland et al. 47 observed that blueberries had relatively lower stomatal responses than other species analyzed.
The smaller dimensions of the subsidiary cells in conditions of extreme light deficiency may be the result of an adaptation to the environment, preventing excessive water loss through the stomata, in a situation in which photosynthetic capacity is negatively affected. 25
Stomata were wide-open at the early green stage of blueberry fruit development and had a slight deposit of wax along the guard cells. As the berries expanded during the initial period of rapid growth, most of the stomata remained near the distal segment of the berries, and by the late green stage, almost none was found in the middle and proximal segments. The majority of these stomata were completely covered with wax when the berries began to change color and ripen. 41
Fruits exposed to sunlight and those partially shaded began to cover their epidermis with a profuse layer of epicuticular wax, which even covered the stomata completely at maturity (Figures 4 and 5).

Image of epidermis of ripe blueberry ‘Brigitta’ fruit that developed exposed to solar radiation, obtained by scanning electron microscopy (1000 X). A profuse deposition of epicuticular waxes is observed.

Image of epidermis of ripe blueberry ‘Ozark Blue’ fruit that was subjected to partial shading throughout its development, obtained by scanning electron microscopy (500 X). The limits of the epidermal cells are not distinguished as they are covered by epicuticular waxes.
Yang et al. 41 found that most of the stomata were completely covered with wax at veraison and that stomatal conductance declined as the fruit developed. However, in the present work fruits developed under darkness had stomata that were not covered with wax (Figures 6 and 7), which suggests the possibility of greater stomatal conductance.

Image of epidermis of ripe blueberry ‘Brigitta’ fruit that was subjected to total shading throughout its development, obtained by scanning electron microscopy (1800 X). The partial deposition of epicuticular waxes is observed, allowing the outline of the epidermal cells to be appreciated.

Image of epidermis of ripe blueberry ‘Ozark Blue’ fruit that was subjected to total shading throughout its development, obtained by scanning electron microscopy (2000 X).
Deposition of epicuticular waxes
Fruits that developed exposed to sunlight showed a profuse deposition of epicuticular waxes on the epidermis (Figure 4), so that the limits of the epidermal cells cannot be clearly distinguished. The fruits that developed under a partial shading condition (80%) also showed a conspicuous waxy layer on the epidermis (Figure 5), indicating that the widespread shading situations do not represent a limitation to the synthesis of epicuticular waxes.
On the other hand, in situations of total shading during fruit development, the deposition of epicuticular waxes was negatively affected, and accordingly, open stomata and defined boundaries of the epidermal cells could be seen (Figure 6).
When comparing fruits from different varieties, Brigitta vs Ozarkblue (Figures 6 and 7), which were developed under complete shading conditions, different patterns of wax deposition are evident among them, which probably have a genetic basis.
The cuticle is the first barrier to light radiation. The increase in thickness of the cuticular wax layer in response to high irradiation has been observed in many species. The spectral quality of light also affects wax biosynthesis, with UV radiation having been shown to increase the quantity and composition of waxes. In fleshy fruits, cuticular waxes play a predominant role in minimizing water loss through a surface often lacking stomata. In the epidermis of fruits, cuticular waxes are composed mainly of triterpenoids and blueberries also contain high levels of β-diketones. Cuticular waxes occur as “intracuticular wax”, of an amorphous nature, which is integrated into a cutin matrix and connected to the polysaccharides of the walls of the underlying epidermal cells, and also as “epicuticular wax”, which can be crystallized as different micro-morphologies. 48
Both the fruits that developed being exposed to sunlight and those that developed under a condition of partial shade presented a conspicuous layer of epicuticular waxes at ripening. These findings differ from those obtained by Yang et al., 41 who determined that blueberries exposed to full sun had thicker wax and cuticle layers than partially-shaded fruits.
Wax deposition was substantially reduced under low light levels, allowing the boundaries of the epidermal cells to be clearly observed. Consequently, only a significantly low solar radiation would affect the synthesis of epicuticular waxes.
Different patterns of wax deposition could be seen in the varieties evaluated, revealing a clear genetic component in the synthesis of epicuticular waxes. In Ericaceae, Dashbaldan et al. 49 determined that the predominant waxes in ripe fruits were triterpenoids, ursolic acid in Vaccinium vitis-idaea (lingonberry) and oleanolic acid in V. myrtillus (bilberry). In previous works, no apparent differences in the morphology of the waxes between different blueberry cultivars had been detected.5,50 The cv. Brigitta presented crystalloids in the form of thick tubules, longitudinally joined to each other. Chu et al. 50 determined that the cuticular wax of ripe blueberry (Vaccinium spp.) fruits is composed of triterpenoids and β-diketones. Triterpenoids are the most abundant wax group in blueberry waxes, and β-diketones are the second one. 51 The characteristic morphology of epicuticular waxes is usually determined by the predominant constituent. 52 In ‘Brigitta’ fruits, Moggia et al.53,54 identified different triterpenoid compounds, mainly lupeol, a secondary alcohol. Secondary alcohols crystallize in the form of tubules. 52 On the other hand, in the cv. Ozarkblue wax crystalloids were arranged as clustered branched rods, a conformation that β-diketone waxes may adopt. 55
Relative water content of fruits
The relative water content (RWC) of the fruits (Figure 8) varied throughout the season, presenting a similar pattern for the three treatments, although more defined in the case of the control treatment, corresponding to the exposed fruits.
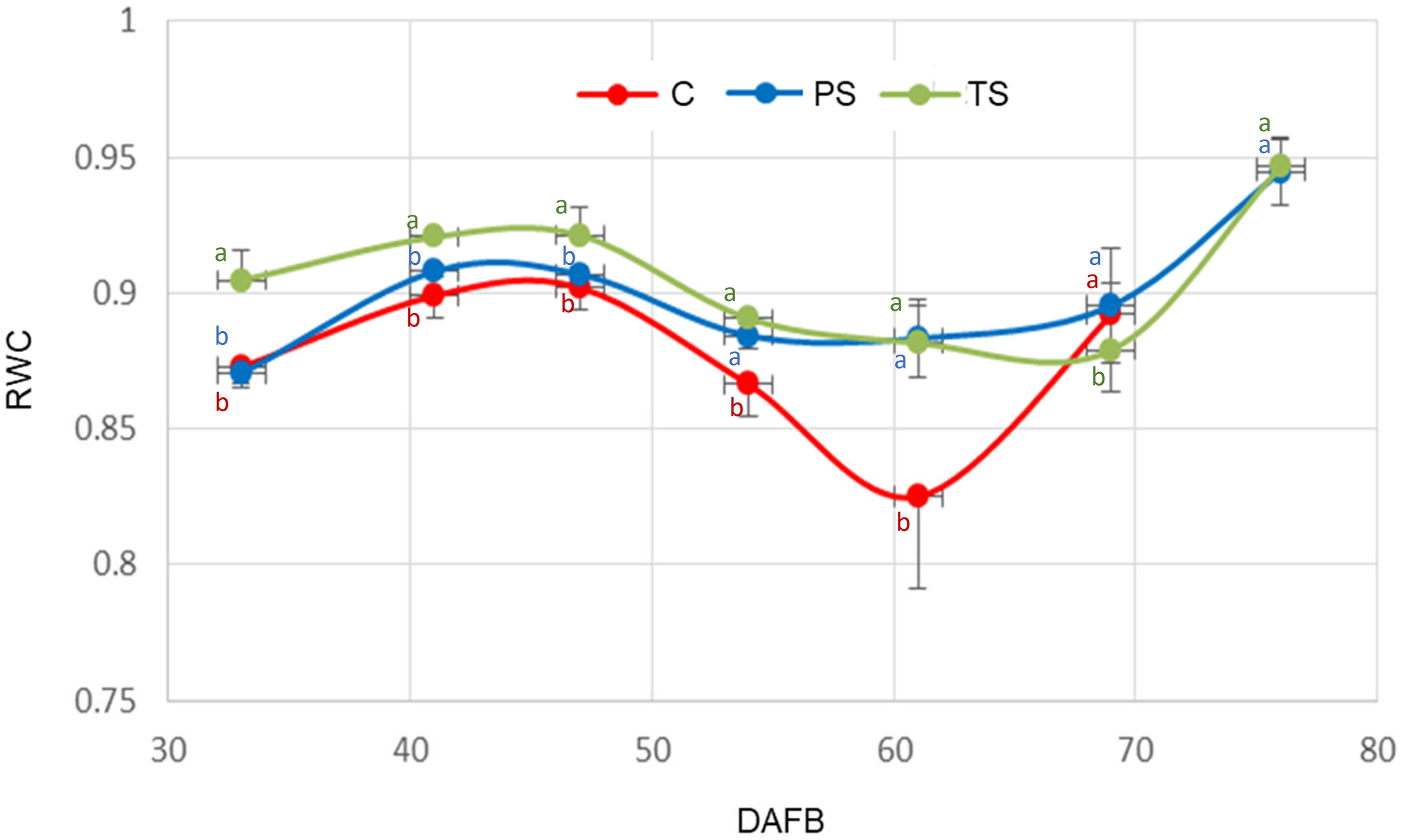
Variation in the relative water content (RWC) of ‘Ozarkblue’ blueberry fruits subjected to different shading treatments during its development. C: control, PS: partial shading, TS: full shading, DAFB: days after full bloom. Bars indicate SE. Means with the same letter are not significantly different from each other (P > 0.05 ANOVA followed by Tukey's test).
RWC was related to the absolute growth rate (AGR) (Figure 9). It is observed that the highest values of RWC correspond to the lowest values of AGR and vice versa, for all treatments. However, the relationship between RWC and AGR was statistically significant (pr < 0.05) only in the total shading treatment. During the second phase of fruit growth (Figure 9), a more pronounced decrease in RWC (Figure 8) was observed in the control, probably due to the loss of water caused by the high atmospheric demand in summer 2022, together with changes in cuticular conductance.
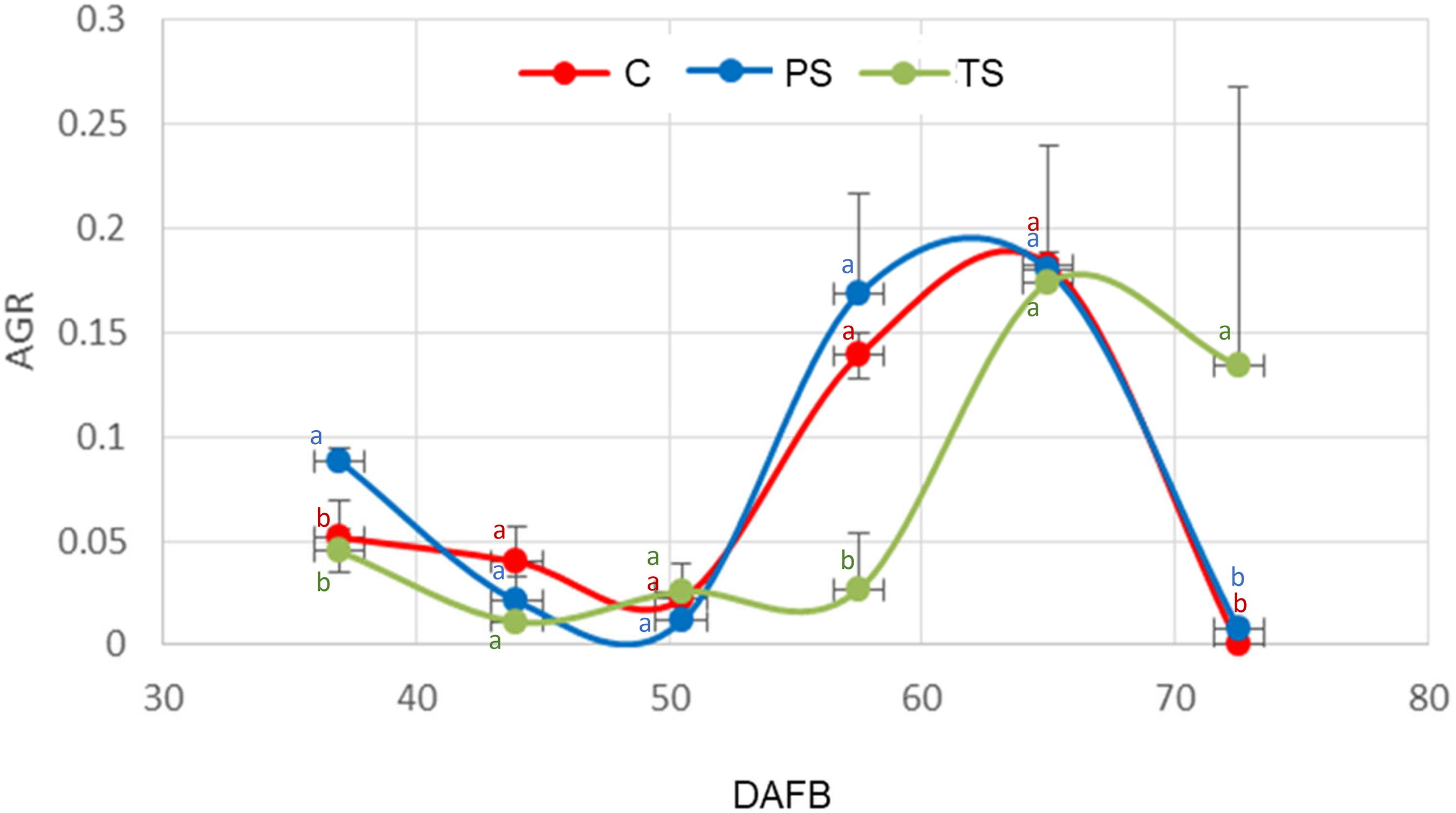
Absolute growth rate of ‘Ozarkblue’ blueberry fruits subjected to different shading treatments during its development. C: control, PS: partial shading, TS: full shading. Bars indicate SE. Means with the same letter are not significantly different from each other (P > 0.05 ANOVA followed by Tukey's test).
During the rapid growth of the fruit, cracks generated in the epidermis, when the limits of the cuticle elasticity are exceeded,56,57 would be the reason for the increase in the conductance, according to the model of Gibert et al.. 58 Konarska 5 observed in blueberry fruits (V. corymbosum) during stage III of the double sigmoid curve, corresponding to the rapid growth phase, the presence of cracks in the pedicellar region of the epidermis.
Fruit transpiration efficiency
The transpiration efficiency of isolated fruits (Figure 10), measured under similar conditions of lighting and evaporative demand, was significantly different between fruits with total shade vs. fruits with partial shade and vs. the control, for the last two dates evaluated (p < 0.05).
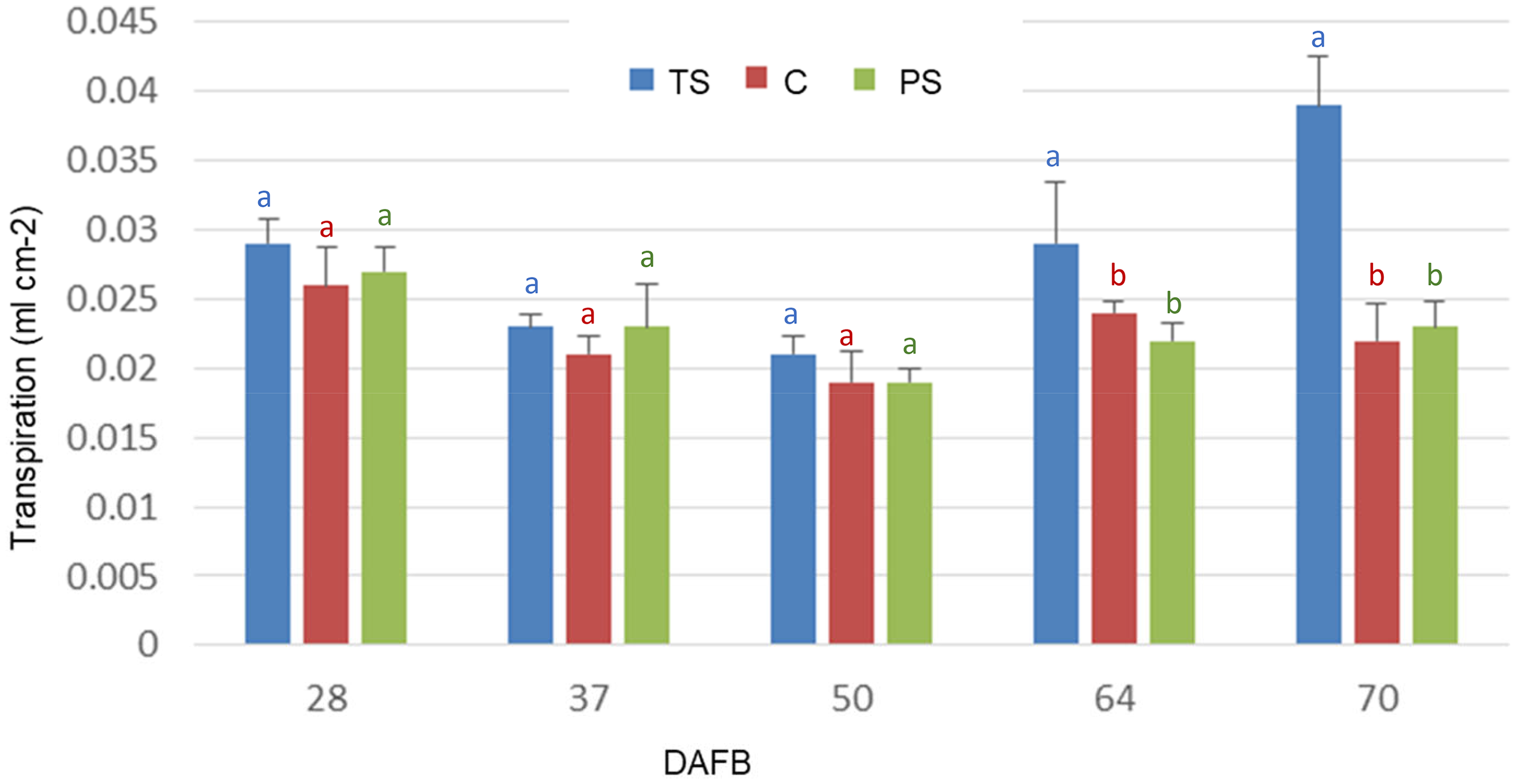
Transpiration of blueberry fruits cv. Brigitta during its development, corresponding to total shading (TS), partial shading (PS) and exposed (C). Bars indicate SE. Means with the same letter are not significantly different from each other (P > 0.05 ANOVA followed by Tukey's test).
The model developed by Gibert et al. 58 for peaches reports a pronounced decrease in the water vapor conductance of the fruit surface during the early stage of fruit development.
The number of stomata is determined at anthesis or even before anthesis, and remains constant during fruit development.59,5 With the expansion of the fruit, the stomata become diluted on its surface, leading to a decrease in stomatal density,10,60 and being located in the calycinal region of the berries.5,41
The decrease in stomatal density would explain the increase in the RWC (Figure 8) and the decrease in the transpiration (Figure 10) during the first phase of fruit growth (Figure 9). The increase in the transpiration (Figure 10) in shaded fruits recorded in the last dates is possibly associated with the scarcity of epicuticular waxes on the fruit surface (Figures 6 and 7).
According to Konarska, 4 the wax layer that covers the fruits increases in thickness during ripening, affecting the transpiration rate, and preventing dehydration of the fruit. 61 The progressive coating of the epidermis with epicuticular waxes, which limits the exchange of water vapor through the stomata, is reflected in the decrease in fruit transpiration.
Taking into account a greater stomatal density in the initial stages of fruit growth, it is expected that gas exchange will have greater preponderance at the beginning of development.
Calcium content
Analyses indicated that fruits exposed to full solar radiation and shaded fruits showed similar (p > 0.05) calcium content (Table 1).
Ca content (ppm dry weight) in the tissues of ripe blueberry cv. Brigitta subjected to different shading treatments.

TS: total shading, PS: partial shading.
Means with the same letter are not significantly different from each other (P > 0.05 ANOVA followed by Tukey's test).
Kalcsits et al. 62 found there was a significant relationship between apple tree position and fruit Ca concentration, where higher Ca concentrations were determined in the upper part of the canopy. The mechanisms underlying these variations were unclear; however, they may be a result of within-tree variation in plant transpiration. In a study carried out on kiwifruit, Montanaro et al. 63 evaluated the influence of exposing branches to different light intensities on transpiration and water flow to the fruit, determining that fruits exposed to the sun accumulated more Ca than shaded fruits.
However, in such experiments the direct effect of exposing the fruits to sunlight was not considered, and a possible additional effect of temperature was not taken into account, which normally increases when tissues absorb solar radiation.
In most fruits, majority of fruit Ca2+ is translocated into the organ within a few weeks after anthesis. 42 Montanaro et al. 64 determined that around 80% of total apricot fruit Ca content was gained within the first 4 weeks after fruit-set. In the same way, 80% of the fruit Ca2+ accumulates during early melon fruit development. 65 Winkler et al. 66 found that Ca import into developing sweet cherry was positively related to changes in fruit transpiration. Fruit transpiration had a maximum rate in very young fruits, from then on transpiration rapidly decreased. Ca accumulation in the cranberry fruit exhibited a sigmoidal curve. Cranberry fruit Ca accumulation was mostly explained by fruit transpiration, which exhibited a similar sigmoidal pattern. 39 It was found that in highbush blueberry stomatal conductance was high at petal fall and rapidly declined as fruit developed. 41 According to this finding, since shading was imposed from fruit set, it may have been somewhat late for detecting significant accumulation of Ca in highbush blueberries.
Conclusions
Differences were found in some anatomical characteristics of the fruits that would be influencing water economy.
The pedicels of exposed fruits presented greater development of phloem tissue than those of completely shaded fruits.
The epidermis of exposed and partially shaded fruits presented a profuse layer of epicuticular wax at maturity, which was almost absent in completely shaded fruits.
The structure of the stomata was modified in the fully shaded fruits, with a decrease in the size of the subsidiary cells, possibly related to greater stomatal control.
With respect to water economy, an inversely proportional relationship between fruit growth rate and fruit relative water content was clear in shaded fruits, with an increase in fruit transpiration during ripening.
Footnotes
Acknowledgments
To the memory of Dr Jorge Tognetti, for his support, enthusiasm, and enriching ideas.
The author acknowledge Lic. Mónica Oppedisano (Microscopy Laboratory—UNMdP) for the technical assistance with scanning electronic microscopy.
Ethical considerations
Ethical approval was not required.
Author contributions
Conceptualization, performance of work, interpretation of data and writing the article: C.G.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Universidad Nacional de Mar del Plata (Grant /projects AGR 482-535-596 UNMdP), Argentina.
Universidad Nacional de Mar del Plata, (grant number AGR 482-535-596 UNMdP).
Declaration of Conflicting Interests
The author declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data availability statement
The data that support the findings of this study are available from the corresponding author, [CG], upon reasonable request.