
Abstract
Recent advancements in Parkinson’s disease (PD) research have both enriched our pathophysiological understanding and challenged conventional therapeutic dogmas. The emerging application of ectodermal mesenchymal stem cells (EMSCs) derived from the cranial neural crest for neuronal regeneration represents a paradigm-shifting therapeutic modality, diverging fundamentally from traditional dopamine-replacement strategies. However, the fundamental mechanisms responsible for their remarkable neurorestorative potential in PD pathophysiology are still not fully understood. This comprehensive review synthesizes current evidence on the pleiotropic therapeutic capacities of EMSCs, focusing on their ectoderm-derived molecular signatures. Central to this review are developmental insights into nasal mucosa-derived EMSCs, particularly their Nestin+ identity, elevated connexin43, niche-specific paracrine activity, and robust dopaminergic differentiation capacity, to guide therapeutic translation for PD. Through systematic interrogation of nasal mucosa-derived EMSC physiology, we aim to establish an evidence-based platform for developing targeted neuroregenerative therapies.
Impact Statement
While ectodermal mesenchymal stem cells (EMSCs) hold therapeutic potential for neurological disorders, current mechanistic comprehension remains incomplete. By elucidating their unique ectoderm-derived molecular characteristic, we demonstrate how EMSCs inherently circumvent the mechanistic limitations of conventional monoaminergic restoration approaches. This synthesis provides a critical framework for advancing translational strategies that bridge stem cell biology with clinically actionable Parkinson’s disease therapies, potentially through the integration of tissue-engineered delivery systems optimized for neural microenvironmental improvement in the future.
Introduction
The continuous advancement of biomedical technologies has driven a progressive extension of human longevity, fundamentally reshaping global demographic patterns. 1 Paradoxically, as people live longer, Parkinson’s disease (PD) is becoming much more common. This age-related neurodegenerative disease damages brain cells, severely impacting patients’ independence and quality of life.2,3 It was characterized by a chronic and progressive nature, imposing substantial burdens on individuals, caregivers, and aging societies. 4 The resultant lack of dopamine in the basal ganglia gives rise to a movement disorder marked by typical Parkinsonian motor symptoms and numerous nonmotor symptoms, with some preceding the motor dysfunction by more than a decade. 5 Currently, conventional treatments primarily rely on levodopa and symptomatic antibody therapy, yet these approaches do not provide a lasting solution to the underlying issue.
The field of stem cell therapy for PD has become a forefront area of medical research. Among the diverse stem cell types under investigation, ectodermal mesenchymal stem cells (EMSCs) have garnered considerable attention due to their distinctive biological origin and strategic location within neurogenic niche. EMSCs can be sourced from various tissues, but this discussion primarily focuses on those derived from the nasal mucosa. As the only tissue readily accessible externally, EMSCs can be easily obtained through biopsy from the reinnervated nasal mucosa. They were initially obtained from cultured rat nasal ensheathing cells and confirmed to be mesenchymal stem cells (MSCs) due to their clonogenic potential and their ability to differentiate into osteogenic, adipogenic, and chondrogenic lineages, as well as the expression of MSCs markers. 6 As research progresses, the ectodermal features of EMSCs are increasingly unveiling unexpected advantages, particularly in neural restoration.7–9 Although EMSC research primarily focuses on spinal cord injury, its significant reparative potential suggests promise for PD treatment.10–13 This promise has progressively generated new hypotheses for PD therapy.14,15 This review explores the therapeutic promise of nasal mucosa-derived EMSCs in PD, examining their developmental origin, key characteristics, and evolving translational strategies. We aim to elucidate the mechanisms underlying their potential to transform PD treatment and offer novel therapeutic approaches for this debilitating disorder.
Neural Crest Origin of EMSCs
Neural crest (NC) resided in the neural plate border constitutes a distinctive structure during embryonic development. 16 It was defined as a population of embryonic progenitor cells that form on the dorsal-most aspect of the neural tube during neurulation (Fig. 1A). 17 Following the invagination and separation of the neural tube, these transient cells undergo an epithelial–mesenchymal transition (EMT) to fulfill NC fate and populate out extensively along the anterior–posterior (AP) axis exhibiting their extraordinary multipotency in forming neurons, glia, muscle, cartilage, melanocytes, adrenal cells, and so on (Fig. 1C). 18 Depending on where the NC is located in the AP axis of the body, four subpopulation concepts named cranial, vagal, trunk, and sacral were later listed (Fig. 1B). 19 In 1983, Northcutt and Gans introduced the ‘new head’ hypothesis, suggesting that the emergence of the vertebrate lineage coincided with the appearance of the NC.20,21 Subsequently, more researchers have been coming to corroborate that cranial NC primarily gives rise to mesenchymal populations responsible for generating a craniofacial skeleton and a new sensory system.22–24 The event of trunk migration along the AP axis in thin streams appears successively and eventually promotes the maturation of the heart, thymic tissue, thyroid, and enteric canal.25–27 This evidence underscores the NC’s critical role in embryonic differentiation. Its developmental trajectory further suggests a resident EMSC population in the cranial region.
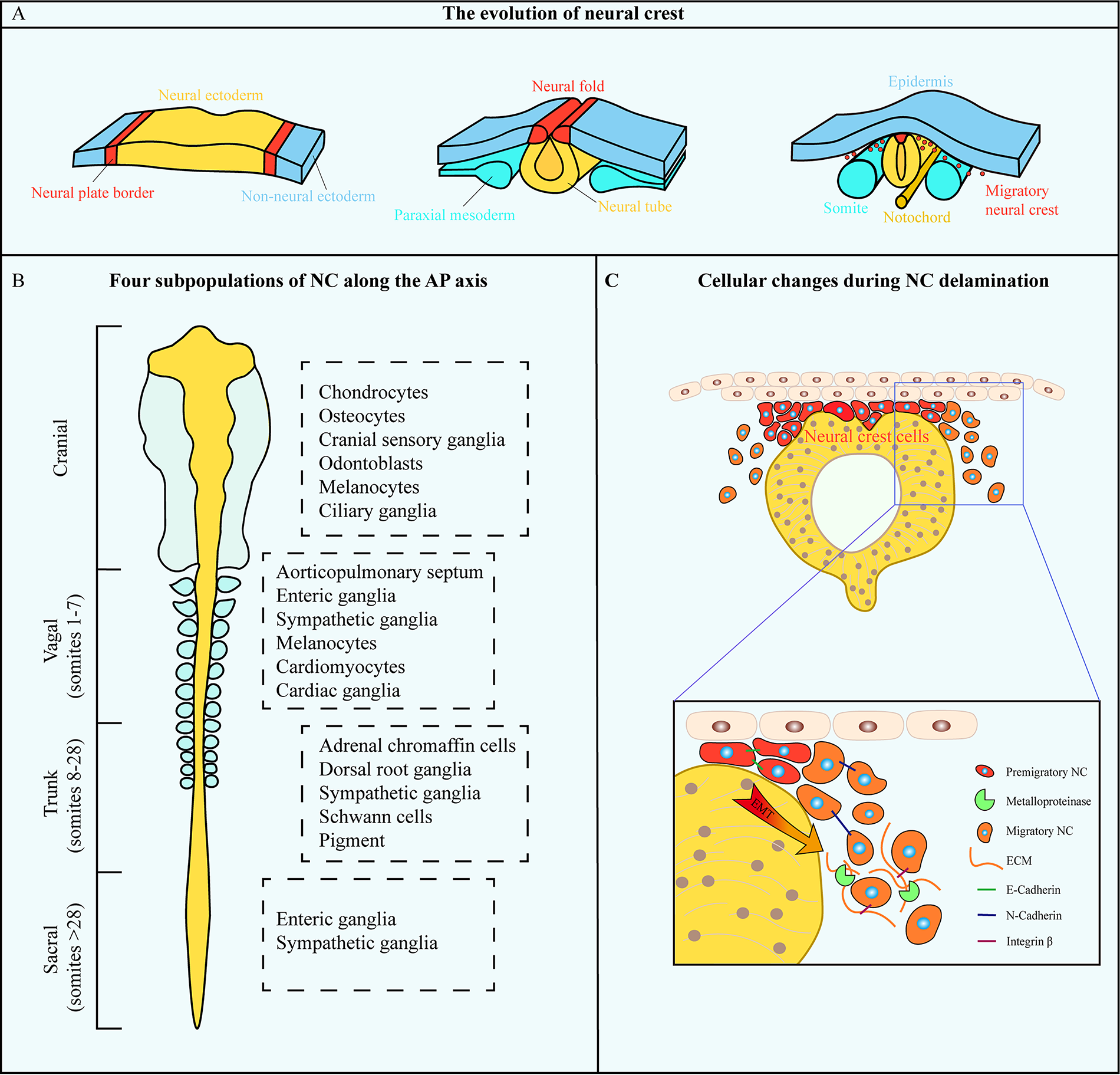
Schematic representation of neural crest development and differentiation.
Source Site of EMSCs
The developmental ontogeny of the NC implies the existence of a specialized ectomesenchymal progenitor population within craniofacial territories. These NC-derived cells undergo EMT to constitute the developmental origin of MSCs. Next, we discuss the predominant tissue sources of EMSCs identified in contemporary studies (Fig. 2).
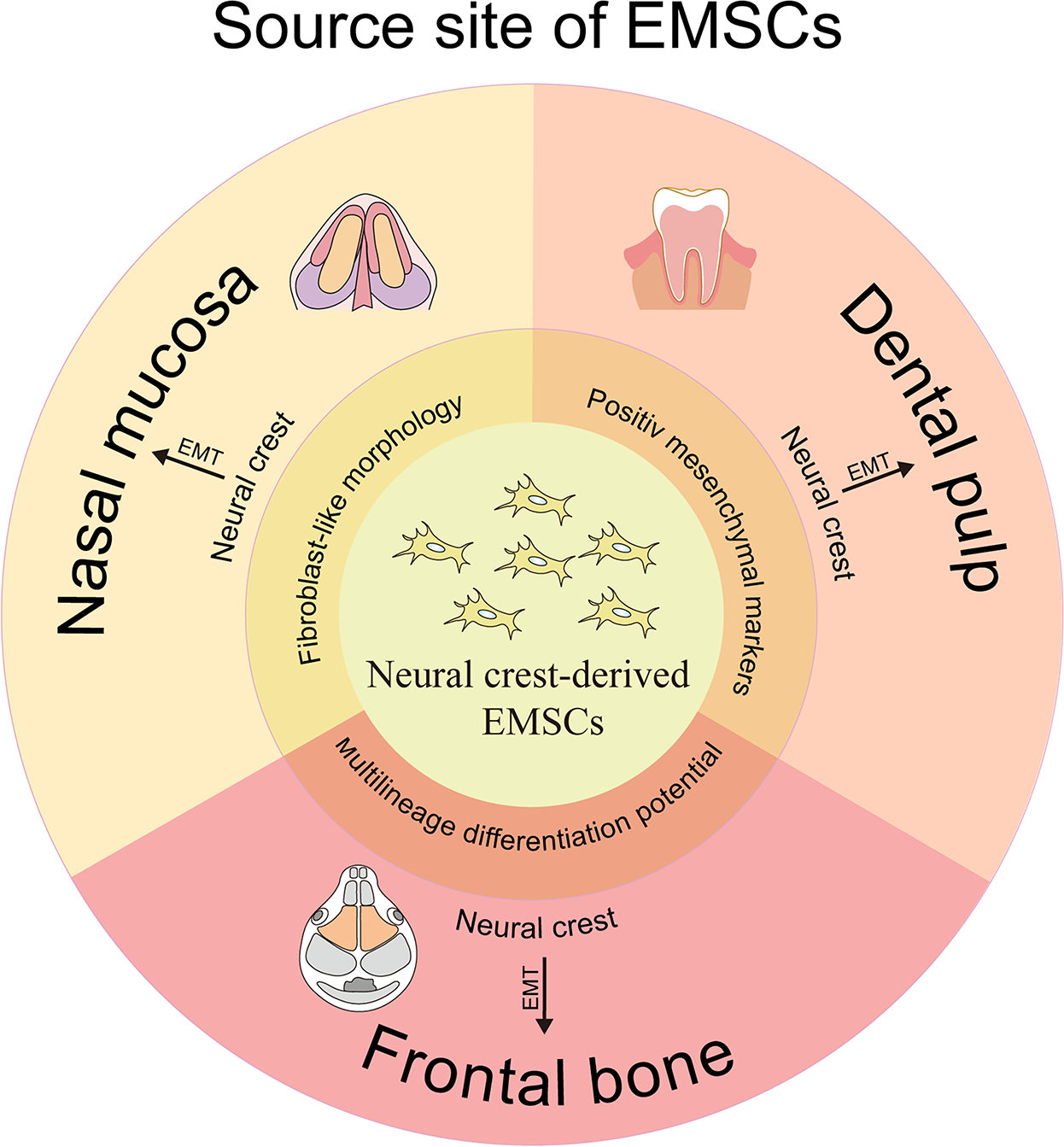
Different source sites of EMSCs originating from the cranial neural crest. EMSC, ectodermal mesenchymal stem cell.
EMSCs could be obtained from the dental pulp. In the initial stages of embryonic development, the dental pulp forms as cranial NC stem cells migrate to the first and second branchial arches. 28 This connective tissue is enveloped by odontoblasts and progenitor/stem cells. 29 In 2000, Gronthos et al. isolated MSC-like populations known as dental pulp stem cells (DPSCs). 30 They revealed that DPSCs share several characteristics with MSCs, including a fibroblast-like morphology, strong adhesion, clonogenic capability, and high proliferation. Subsequent studies led to the identification of three additional types of dental-MSCs-like populations from exfoliated deciduous teeth, known as SHED, 31 periodontal ligaments, 32 and apical papilla. 33
Frontal bone constitutes another EMSC source. One published hypothesis, spanning over three decades, proposes that the NC is responsible for forming the skull and its related connective tissues. In contrast, placodes are involved in developing sensory organs.34,35 Thus, researchers sought to isolate MSCs derived from the ectoderm of the frontal and parietal bones. 36 The ectoderm cell populations originating from the frontal bone (FbMSCs) displayed a fibroblast-like morphology and tested positive for mesenchymal markers CD29 and CD105, while showing negativity for markers associated with hematopoietic and endothelial cell lineages, namely CD31 and CD45. Concurrently, FbMSCs exhibited high levels of the ectodermal-specific transcription factor Tfap2β expression, signifying their ectodermal identity.
In recent years, we have also successfully isolated EMSCs from the lamina propria of the nasal mucosa.37,38 These cells coexpress canonical NC (Sox1, Sall4, Snail, Sox10) and MSC markers (S100β, CD133, Nestin, vimentin, CD44). When cultured in neural basal medium (serum-free Dulbecco's Modified Eagle's Medium [DMEM] supplemented with 2% B27, 20 ng/mL epidermal growth factor [EGF], and 20 ng/mL basic fibroblast growth factor [bFGF]), EMSCs demonstrated robust proliferation and the ability to form spherical clusters. Furthermore, the EMSCs demonstrated multilineage differentiation potential, such as osteogenic, adipogenic, and neurogenic differentiation.39,40 The findings indicated that the nasal mucosa represents a reliable source of EMSCs.
Nasal Mucosa: A Privileged Source of EMSCs
Although EMSCs can be isolated from diverse anatomical sources, research efforts increasingly prioritize nasal mucosa-derived populations. Given the scant comparative evidence delineating source-specific characteristics and therapeutic potential, we mainly discuss the superior therapeutic potential of nasal mucosa-derived EMSCs through the dual lenses of developmental biology and clinical translational potential.
Anatomical advantages of nasal mucosa EMSCs
The vertebrate nasal system, derived embryonically from both NC and olfactory placode lineages, comprises the olfactory epithelium (OE), olfactory bulb, and nasal cortex.34,41–43 Within the OE, olfactory sensory neurons (OSNs) mediate chemosensory transduction but remain highly vulnerable to environmental damage due to their location in the nasal cavity. Crucially, olfactory function is preserved by adult neurogenesis driven by resident basal stem cells: globose basal cells (GBCs) and horizontal basal cells (HBCs) (Fig. 3).37,44–51 However, this robust regenerative capacity extends beyond neuroepithelium stem cells (GBCs, HBCs) alone, with relatively poor proliferative rates and restricted differentiation potential.
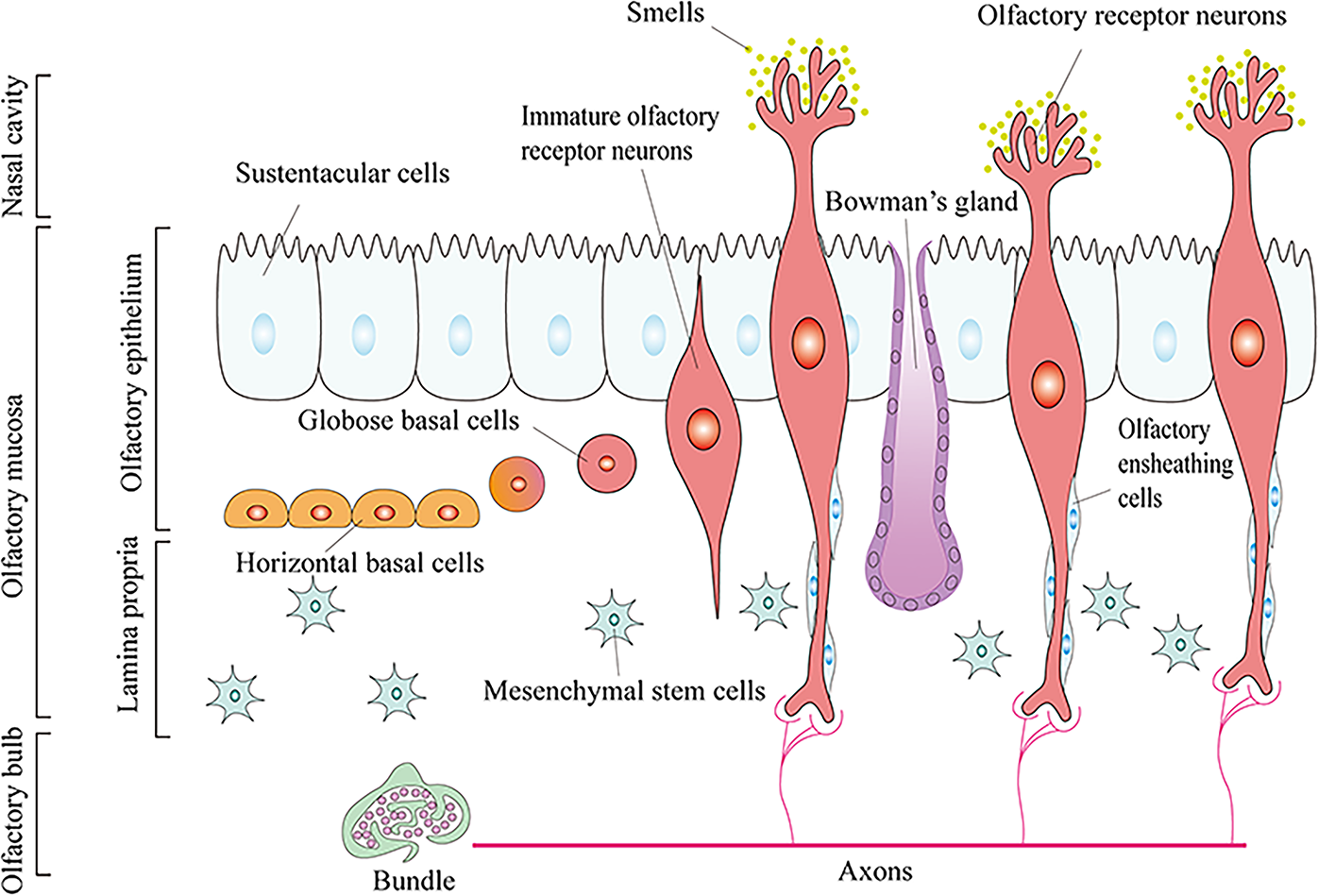
Composition of the olfactory system.
EMSCs, the third type of stem cell identified in the nasal mucosa, reside beneath the neuroepithelium and exhibit dual neural/mesenchymal properties. 8 Evidence indicates that they originate from Sox1+ neuroepithelium (not mesoderm), representing the earliest embryonic trunk MSCs. 52 Their unique association involves neuroepithelium-lamina propria signaling essential for olfactory development. 53 Meanwhile, their role is crucial for niche biology within this tissue. 54 Scholars propose EMSCs can migrate across compartments postinjury, traverse the basement membrane, differentiate into neurons, and replenish damaged OE. 8 This provides a plausible mechanism for olfactory function recovery. Furthermore, located in the basal layer beneath neuroepithelial stem cells, EMSCs may promote the neural differentiation and regeneration of neighboring basal stem cells through paracrine signaling. 55 While current research on EMSCs in olfactory regeneration remains limited, their multifaceted roles position them as a promising strategy for neural regeneration therapies.
Translational advantages of nasal mucosa EMSCs
The clinical translation of MSCs, sourced from tissues such as fat, bone marrow, or umbilical cord, faces significant hurdles primarily rooted in tissue acquisition. Harvesting adipose tissue or bone marrow requires invasive surgical procedures, posing patient risk and discomfort. While umbilical cord tissue avoids surgery, its utilization introduces substantial logistical and economic burdens related to long-term cryopreservation and associated costs. Nasal mucosa EMSCs present a promising alternative, enabling rapid tissue collection via a simple, minimally invasive biopsy performed directly on the patient. Critically, this biopsy preserves olfactory function intact. 56 The resected tissue undergoes regeneration due to the inherent regenerative niche of the nasal mucosa. By eliminating the need for complex surgery and costly long-term storage, EMSCs present distinct advantages in patient accessibility, reduced procedural burden, and overall cost-effectiveness, positioning them as a compelling solution to key translational challenges in MSC therapies.
EMSCs isolated via the tissue adherent method exhibit a flat, fibroblast-like morphology. 57 The antigenic profile and differentiation properties of EMSCs and bone marrow MSCs (BMSCs) are similar. 58 MicroRNA fingerprinting further revealed that EMSCs are 64% homologous to classical BMSCs. 59 Interestingly, these cells demonstrate significantly higher proliferative capacity than BMSCs in vitro, evidenced by a nearly threefold increase in population doubling time at week 2 postplating. Furthermore, EMSCs maintain long-term self-renewal capability in culture (>15 weeks), contrasting sharply with BMSCs, which fail under these conditions. 8 This dramatic difference in proliferative potential is mainly attributed to dissimilarities in telomere regulation. 60
Nasal Mucosa EMSCs: Translating Developmental Advantages into PD Therapeutics
Parkinson’s disease
The global rise in life expectancy has precipitated a surge in age-related neurodegenerative disorders, with PD exhibiting particularly alarming growth rates. 61 Characterized by progressive movement impairment and mainly occurring in the elderly, PD has become a major neurological challenge of the 21st century. 62 Epidemiological data reveal that the global population living with PD has more than doubled over recent decades, exceeding 6 million individuals, and projections indicate sustained acceleration of this trend. 63 The Global Burden of Disease study reported that PD was the fastest-growing neurological disorder between 1990 and 2016. 64 The latest estimates indicate that in 2019, PD contributed to 5.8 million disability-adjusted life-years, marking an 81% increase since the year 2000. 65 This escalating prevalence and disease burden trajectory has generated profound global public health concern.
PD is characterized by the progressive degeneration of dopaminergic neurons in the substantia nigra pars compacta (SNpc), leading to both motor and nonmotor symptoms.66,67 The hallmark motor impairments collectively define the clinical diagnosis, including bradykinesia, resting tremor, muscle rigidity, and postural instability.68,69 Concurrently, nonmotor symptoms frequently manifest, including nasal dysfunction,70,71 pain and dysautonomia,72,73 gastrointestinal dysfunction,74,75 and cognitive and neurobehavioral abnormalities.76,77 Motor and nonmotor circuit disturbances in PD: which happens first? The protein dynamic changes may be the pivotal clue to resolving this problem. 78 Cellular proteostasis mediated through a vast network of biochemical pathways was closely bound to neurodegenerative diseases. 79 When the equilibrium of proteostasis was disrupted, it led to the pathological consequence of protein aggregation. 80 Different pathological protein consequences, including synucleinopathies, amyloidopathies, tauopathies, contributed to the classification of neurodegenerative disorders. PD is molecularly classified as a synucleinopathy due to the essential histopathological hallmark: the aggregation of α-synuclein (a 14-kDa neuronal protein highly expressed in the brain) into intraneuronal Lewy bodies and Lewy neurites. 81 Pathological α-synuclein is thought to initially spread from the peripheral to the central nervous system (CNS), first causing nonmotor symptoms. It subsequently affects the mesencephalon, accounting for the cardinal motor features, and finally progresses to the neocortex as dementia develops. 82 Dopaminergic neurons in the dense part of the substantia nigra transport dopamine transmitters to the striatum (including the putamen and caudate nucleus) via the substantia nigra-striatum pathway and are involved in motor regulation of the basal ganglia. The loss of these dopaminergic neurons, caused by α-synuclein accumulation, leads to a significant decrease in dopamine levels in the striatum. This key biological event ultimately disrupts neuronal circuitry, underpinning the motor dysfunction (Fig. 4).
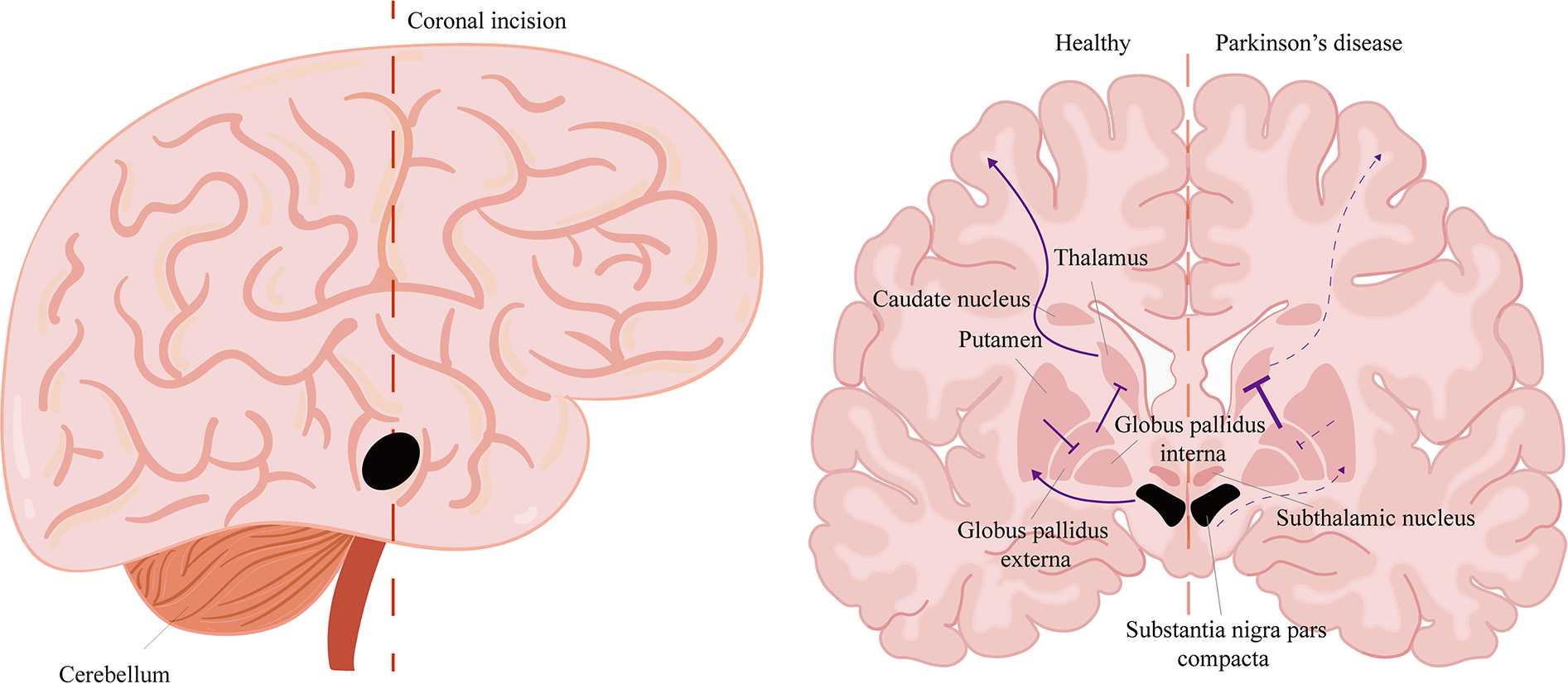
Schematic of the neuronal circuits in PD. For simplicity, the pathways are shown directly, with solid lines indicating active pathways and dashed lines indicating inactive pathways. Normally, this circuit facilitates movement, but in PD, the output is reduced. The midbrain substantia nigra pars compacta (SNc) provide dopaminergic input to the putamen, which excites the direct pathway. The putamen inhibits the globus pallidus interna, which then suppresses the thalamus. The thalamus projects excitatory input to the motor cortex. In PD, degeneration of the SNc leads to a decreased signal in the putamen. Subsequently, the globus pallidus externa enhances the inhibition of the thalamocortical projection, thereby limiting movement. PD, Parkinson’s disease.
In summary, PD is defined by two core pathological features: the selective loss of dopaminergic neurons in the substantia nigra and the widespread accumulation of α-synuclein aggregates. A deeper understanding of these molecular and cellular mechanisms is essential for developing innovative treatments that can halt or reverse disease progression, ultimately alleviating the growing global burden of PD.
Current therapeutic paradigms for PD
Conventional pharmacological/surgical interventions
The management of PD has historically relied on pharmacotherapy. George Cotzias’ pioneering introduction of high-dose L-3,4-dihydroxyphenylalanine (L-DOPA) in the 1960s revolutionized PD therapy,83,84 establishing a pharmacological foundation that persists today as the gold-standard symptomatic treatment.
85
Despite decades of searching and exploring,
Recent advances in PD therapeutics have seen the development of pathogenesis-targeted strategies. 90 Central to this is α-synuclein, a potentially toxic protein in cellular processes driving PD-associated neurodegeneration. 91 Direct application of α-synuclein monoclonal antibodies has minimized the accumulation and spread of aggregated α-synuclein, demonstrating therapeutic potential.92,93 Similarly, certain polyphenols can counteract α-synuclein aggregates through detoxification. 94 The adjacent dihydroxyphenyl groups conferred the ability to inhibit α-synuclein fibrillation upon the flavonoids. 95 The evidence also suggested that the essential process for inhibiting α-synuclein fibrillation is the covalent binding of oxidized flavonoid species to α-synuclein. Intriguingly, the full therapeutic efficacy of flavonoids in PD extends beyond current mechanistic understanding. Beyond α-synuclein pathology, neuroinflammation mediated by inflammasome activation critically contributes to PD pathogenesis. Baicalein, a prominent bioactive flavone compound derived from Scutellaria baicalensis, also suppresses neuroinflammation. 96 Baicalein mitigates neuroinflammation via the NLRP3/caspase-1/GSDMD pathway, highlighting its distinctive anti-inflammatory properties and therapeutic relevance in PD. 97 Furthermore, compelling evidence demonstrates Baicalein’s neuroprotective effects and role in maintaining neurotransmitter homeostasis.98,99 While these polyphenolic compounds effectively attenuate α-synuclein-mediated pathology, their application may be insufficient for addressing the multifactorial pathophysiology of moderate-to-severe PD, particularly in cases involving extensive dopaminergic neuron loss and compromised neural circuitry.
Beyond pharmacological interventions, subthalamic burst firing represents another pathophysiological target. 100 Deep brain stimulation (DBS) surgically treats PD by implanting electrodes in targeted brain regions, with proven efficacy in patients.101,102 An electrical current could trigger a series of influences, including increased blood flow, hyperactive neurogenesis, and improved local release of neurotransmitters.103–106 Undeniably, DBS has substantially advanced our understanding of PD pathogenesis over the past two decades. However, the side effects, management of nonmotor symptoms, accurate lead placement through intraoperative testing, and selection of optimal stimulation parameters remain limitations in DBS treatment. 107 Providing convenient treatment poses a challenge due to its time-consuming and empirical nature, involving complex programming sessions in a specialized clinic.108,109
Cell-based regenerative strategies
The irreversible progression of PD stems primarily from the CNS’s limited regenerative capacity, rendering conventional therapies ineffective against neurodegeneration. This therapeutic impasse has driven a shift toward regenerative strategies, particularly cell-based interventions to restore dopaminergic circuitry. Following the clinical success of fetal midbrain dopamine tissue transplants, dopamine neuron replacement therapy has become a significant focus in PD treatment over recent decades. 110 Although this approach demonstrates efficacy, sourcing sufficient dopamine neurons remains the primary challenge. 111 Rapid advances in reprogramming technology offer a promising solution.112,113 By manipulating cell fate, researchers can develop PD cell therapies based on this approach. 114 Evidence confirms the feasibility of converting somatic cells into dopaminergic neurons,115–117 with these reprogrammed cells demonstrating significant therapeutic potential. 118 However, low conversion efficiency and poor survival of directly reprogrammed cells posttransplantation present significant hurdles for widespread clinical application.
Valued for their potent paracrine regenerative capacity, MSCs have become a leading focus in regenerative medicine. 119 This is particularly evident in PD, where MSC infusion reduces motor impairments such as uncoordinated limb movement.120,121 The regenerative microenvironment could be reshaped by the neurotrophic growth factors secreted by MSCs, including glial cell-derived neurotrophic factor, brain-derived neurotrophic factor, nerve growth factor, and vascular endothelial growth factor.122–124 Beyond promoting neurological preservation, repair, and neurogenesis, the MSC secretome functions as a potent immunomodulator. 125 It modulates three pivotal immune phases: antigen presentation, T-cell activation/proliferation, and effector responses. This immunomodulation holds significant clinical relevance, as neuroinflammation is a well-established driver of progressive neurodegeneration and neuronal loss in PD. 126 Recent research demonstrates that immunomodulatory MSCs protect dopaminergic neurons in PD models by suppressing NLRP3 inflammasome-mediated neuroinflammation. 127 Furthermore, emerging evidence indicates that MSCs promote α-synuclein clearance through M2 microglia polarization. 128 Specifically, MSC-derived interleukin-4 (IL-4) induces this protective microglial phenotype, subsequently mediating neuroprotection. These mechanisms position MSC-based therapy as a promising therapeutic candidate for PD.
Exploiting nasal mucosa EMSC-specific therapeutic traits
The efficacy of advanced regenerative cellular therapies hinges upon multifaceted determinants, with the selection of optimal seed cells constituting a pivotal determinant of therapeutic outcomes.129,130 EMSCs demonstrate unique niche-specific advantages among emerging candidates, positioning them as prime biological agents for PD intervention.
Nestin-positive EMSCs
As a type VI intermediate filament protein, Nestin is an established proliferation marker in diverse stem and progenitor cells.131–133 Significantly, its role as a specific NC stem cell marker underlies the niche-dependent functional properties of these cells, including multipotency, enhanced proliferative capacity, self-renewal, and regenerative potential.134,135 This association makes Nestin-positive stem cells highly valuable targets for tissue engineering strategies.
Our previous work demonstrated that nearly all EMSCs isolated from nasal mucosa strongly express Nestin. 38 Comparative analyses of Nestin expression revealed a striking divergence: nearly 100% of EMSCs tested positive, only ∼50% of BMSCs expressed this marker despite identical isolation protocols. 136 Achieving the desired cell number in clinical trials from EMSCs is significantly faster than from BMSCs. 137 As noted, some scholars attribute the superior proliferative capacity of EMSCs over BMSCs to telomere regulation. We propose instead that Nestin-positive status is the more likely underlying mechanism. Hedgehog (Hh) signaling pathway plays a central role in regulating the proliferation of adult stem cells. 138 Nestin expression is widely observed during stem cell expansion and tissue regeneration, where Hh pathway activation is frequently involved. Research confirms that Nestin enhances Hh signaling by targeting Gli3, with its presence leading to significantly prolonged and elevated pathway activation levels. 139 This indicates a greater proliferative capacity in Nestin-positive cells. Furthermore, Nestin knockdown across multiple cell types consistently demonstrated that reduced Nestin expression significantly impaired cellular proliferation.140–142 The rapid expansion capability of Nestin-positive EMSCs provides a critical advantage for PD treatment by ensuring adequate cell supply for transplantation.
The functional benefits of Nestin expression to EMSCs extend beyond this proliferative advantage. Some researchers have shown that the Nestin+ BMSC subpopulation, isolated from highly heterogeneous BMSCs, significantly enhanced myocardial infarction repair compared with the Nestin+ BMSC group. This effect is primarily attributed to the enhanced CXC chemokine ligand 12 (CXCL12) secretion by Nestin+ BMSCs. 143 Since Nestin is expressed in almost all EMSCs, we expected the results from other studies demonstrating their superior CXCL12 secretion relative to BMSCs.144,145 Based on these findings, we infer that Nestin expression positively correlates with CXCL12. CXCL12, a small immunomodulatory cytokine, is a potent chemoattractant for lymphocytes and monocytes.146,147 Beyond recruitment, CXCL12 further modulates monocyte differentiation toward an immunosuppressive program. 148 Critically, a recent study demonstrates that CXCL12 secreted by BMSCs directly drives macrophage polarization toward the M2 phenotype. 149 Resident CNS macrophages, termed microglia, can polarize into pro-inflammatory (M1) or anti-inflammatory (M2) phenotypes. In the progression of PD, sustained activation of microglia by α-synuclein, environmental toxins, or pathogens promotes polarization to the M1 phenotype. This sparks inflammatory cascades, driving progressive neurodegeneration. 150 Therapeutic approaches in PD achieve benefits by driving microglial polarization from the M1 to M2 phenotype and controlling neuroinflammation, curtailing dopaminergic neuron loss.151–153 Consequently, Nestin+ EMSCs could exhibit enhanced PD therapeutic effects through CXCL12-mediated microglial M2 polarization, augmenting immunomodulatory capacity. This mechanistic insight is consistent with evidence of EMSCs’ robust ability to induce M2 phenotypic switching in macrophages.154,155
Nestin-positive cells function as crucial regulators in the CNS postinjury. Cawsey et al. evaluated the changes and effects of these cells in vivo following CNS injury. Their results demonstrated Nestin-positive cell detection across three distinct spinal segments, indicating that Nestin reactivity is not merely a localized injury response. Upon activation, these cells may contribute to injury recovery through their neuroendocrine functions and neurogenic potential. 156 Notably, in fish cerebellar injury models, Nestin is reexpressed by neural stem cells within neurogenic niches and adjacent regions, exerting neurotrophic and neurogenic effects that further support neural repair mechanisms. 157 More interestingly, some scholars indicate that Nestin-positive neural progenitors mediate adult neurogenesis of nigral dopaminergic neurons in mice. 158 These insights position Nestin-positive EMSCs as a promising therapeutic strategy with inherent advantages for PD treatment.
EMSCs exhibiting high Connexin43 levels
Connexin 43 (Cx43), the predominant connexin isoform, classically forms gap junctions (GJs) that enable direct intercellular transfer of ions, metabolites, and signaling molecules (<1 kDa).159,160 Its assembly begins with oligomerization of six monomers into connexons (hemichannels) in the endoplasmic reticulum, which subsequently traffic to the plasma membrane and dock with counterparts on neighboring cells at specialized sites to establish functional GJs. 161 Beyond this canonical role, proteomic analyses strengthened the biological involvement of Cx43 beyond gap junctional intercellular communication, including transcription, metabolism, autophagy, and ion channel trafficking.162,163 GJs enable NC cell migration through ubiquitous membrane localization and functional intercellular coupling.164,165 Specifically, N-cadherin (N-cad) and Cx43 coordinate NC motility by engaging the locomotory apparatus via p120 catenin-mediated signaling.166,167 Given their NC origin, EMSCs likely retain this cellular characteristic developmentally. Subsequent experimental validation in our study definitively demonstrated robust Cx43 expression in EMSCs, substantiating this critical phenotypic characteristic. 37
The normal brain’s most prevalent and active cells (neurons, astrocytes, and oligodendrocytes) abundantly express connexins.168,169 A selective pattern of connexin expression in the CNS contributes to organizing these cell populations into distinct functional networks. 170 Densely interconnected via GJs, these cells form functional networks that operate as integrated units rather than isolated entities. The networks exert a broad neuroprotective influence by distributing metabolic substrates and dynamically regulating extracellular ion gradients through spatial buffering mechanisms.171,172 However, others argue that GJ-formed networks can exacerbate disease progression by inducing bystander cell death. 173 This occurs as the continuous transmission of harmful substances facilitates the expansion of the damaged area via the “bystander effect.” 174 The dual nature means the current understanding of GJs is paradoxical, as they exhibit aggravating and alleviating effects. 175 It is reasonable to assume that bystander cooperation is protective in minor injury cases (which can be adaptively adjusted). However, the bystander effect manifests when injury severity surpasses a critical threshold. The transplantation of EMSCs exhibiting high Connexin43 expression levels confers unique advantages, notably elevating the impairment threshold by augmenting “bystander cooperation.”
Ferroptosis, implicated in PD-associated pathological cell death, is driven by aberrant iron accumulation in the brains of PD patients.176,177 This iron overload initiates the Fenton reaction during PD progression, generating destructive hydroxyl radicals that mediate cytotoxicity and ultimately induce ferroptosis.178,179 Critically, PD pathogenesis occurs when iron ion accumulation surpasses a critical threshold, 180 suggesting that modulating the ferroptosis threshold represents a viable therapeutic strategy. 181 In this context, transplantation of EMSCs with high connexin43 expression emerges as a promising intervention. These EMSCs establish GJs with neural cells, facilitating intercellular ion transfer and significantly elevating the ferroptosis induction threshold. By forming “rescue” channels with damaged dopaminergic neurons, EMSCs enable recipient neurons to tolerate elevated iron loads, enhance their antioxidant capacity, and reduce susceptibility to ferroptotic stimuli. Although direct experimental evidence for the involvement of GJs in ferroptosis remains limited, their capacity to mediate intercellular redistribution of drugs is well-established in tumor chemoresistance research. 182 This functional parallel suggests that GJs may similarly facilitate iron ion (Fe2+) transfer between adjacent cells during ferroptosis. Structurally, GJ channels (1–1.4 nm in width) enable passive diffusion of Fe2+ (ionic radius 76 pm) and small molecules (<1 kDa).183,184
Niche-specific paracrine characteristic of EMSCs
Nasal sensory neurons are exceptional within the mammalian nervous system due to their lifelong neurogenesis and capacity for neuronal replacement following injury. 185 The regeneration of the olfactory neuronal population is driven primarily by stem cells (basal cells) residing in the OE. 186 Crucially, EMSCs constitute essential contributors to the specialized niche, which sustains continuous neurogenesis and neuronal integration and facilitates the renewal and functional incorporation of new OSNs into the olfactory circuitry. 187
Functionally linked to their olfactory localization, EMSCs leverage paracrine signaling to support neuroepithelial regeneration. Zhang et al. confirmed enrichment of mitogenic factors such as EGF and bFGF in EMSC-conditioned medium.188,189 By stimulating neural precursor proliferation and differentiation, these secretions may maintain basal stem cell homeostasis in the neuroepithelium. 190 The significant roles of these neurotrophic factors in dopamine neuron viability are well-established. The neurotrophic activity of EGF on midbrain dopaminergic neurons is evidenced by its supplementation preventing degeneration 191 and intracerebroventricular administration of EGF improving dopaminergic neuronal survival/function. 192 At nanogram/milliliter concentrations, bFGF exhibits superior dopaminergic neurotrophic activity to EGF in fetal midbrain neurons. 193 Further studies indicated it supports dopaminergic neuron survival, growth in vitro, and neuroprotection.194,195
Sonic Hedgehog (SHH) contributes to CNS neurogenesis and neural patterning during development. 196 Its dysregulated signaling is closely linked to PD progression. 197 Exogenous SHH protein reduces behavioral deficits in rat models of PD. However, unlike typical pharmacological agents, higher doses of SHH did not enhance its therapeutic effects. 198 This therapeutic efficacy was further confirmed through lentivirus-mediated striatal delivery of sonic hedgehog. The observed benefit likely stems from SHH-mediated preservation of dopaminergic neurons, combined with induced sprouting of dopaminergic fibers and increased dopamine production. 199 Furthermore, studies using purmorphamine, a small-molecule SHH signaling agonist, validate the therapeutic role of SHH signaling in PD. 200 It is now widely accepted that SHH protects late-stage dopaminergic neurons. 201 Researchers propose that SHH exerts a protective effect early in PD progression by regulating inflammation through microglial activation, while later it reduces dopaminergic neuron loss. These evidences point to SHH administration as a promising novel approach for treating PD. 202 Our previous data demonstrated that EMSCs strongly express SHH, and SHH can also be detected in the conditioned medium of EMSCs. 203 On the contrary, coculture evidence shows EMSCs promote both OPC differentiation/maturation and osteogenic differentiation of MC3T3-E1 cells through SHH secretion.204,205 Notably, EMSC may offer therapeutic benefits for PD through SHH release.
It has been established that immunoregulatory factors, including TGF-β and IL-10, secreted by MSCs contribute to forming an immunosuppressive environment. 206 EMSCs possess a greater IL-10 and TGF-β production capacity than BMSCs. 207 Therapeutic targeting of the anti-inflammatory cytokine IL-10 is reported to directly regulate immune responses and reduce neurodegeneration in PD. 208 Some scholars have pointed out that IL-10 plays a multifaceted role in PD, enhancing microglial phagocytic activity, reshaping the local immune response, and ultimately promoting the survival of dopaminergic neurons. 209 Similarly, TGF-β is pivotal in maintaining immune homeostasis and preventing excessive inflammation. 210 Within the context of PD, these cytokines additionally mediate immune regulation by modulating astrocyte phenotypes. 211 The immune regulatory effects conferred by EMSC paracrine signaling thus constitute a significant mechanism for inflammation control during the early stages of PD.
Progressive dopaminergic neuron loss is critically exacerbated by persistent neuroinflammation. This interplay underscores the urgent need for therapies that simultaneously target neuroinflammation and protect dopamine-producing neurons. EMSCs hold great potential precisely because their innate paracrine machinery provides a unified therapeutic platform. The factors they secrete enable modulation of immune responses, counteracting neurotoxic inflammation, and direct support for neuronal health. This synergy within EMSC biology offers a fundamentally new approach to disrupting PD progression.
Inherent dopaminergic differentiation capacity of EMSCs
Substantial progress has been made in directing the differentiation of MSCs toward dopaminergic neuronal phenotypes through external interventions.212–214 Notably, research has established that the efficient induction of dopaminergic phenotypes in MSCs depends on key factors, particularly SHH, bFGF, and FGFs.215,216 Significantly, EMSCs exhibit robust endogenous expression of these factors (as discussed in the above chapter). This intrinsic expression profile positions EMSCs as a unique source for dopaminergic differentiation. Recent research confirms this superiority, demonstrating EMSCs’ enhanced capacity for dopaminergic neuron differentiation compared with umbilical cord-derived MSCs (UC-MSCs). 217 Owing to their shared neuroectodermal origin with the nervous system, EMSCs showed significantly higher expression levels both at the mRNA and protein levels of the neural marker MAP2 and dopaminergic markers (tyrosine hydroxylase, dopamine transporter, and PITX3) relative to UC-MSCs. In a rat model of 6-hydroxydopamine-induced PD, implantation of EMSCs into the substantia nigra pars compacta ameliorated Parkinsonian symptoms. 218 This behavioral recovery was attributed to the in vivo dopaminergic differentiation of the engrafted EMSCs.
While direct evidence for in vivo dopaminergic differentiation of EMSCs is scarce and transformation efficiency is undefined, current research implies that such differentiation likely operates synergistically with key EMSC therapeutic functions. Critical among these are neurotrophic support and immunomodulation, which jointly mitigate Parkinsonian pathology.
Conclusions and Future Directions
The anatomical accessibility of the nasal mucosa EMSC source enables minimally invasive harvesting through outpatient endoscopic procedures, significantly enhancing its clinical feasibility for autologous transplantation strategies. EMSCs from the nasal mucosa maintain stable expansion without senescence through P10, providing sufficient cell quantities for clinical translation with high therapeutic promise. Meanwhile, they could overcome critical limitations observed in conventional MSC therapies, including donor site morbidity and restricted neural differentiation capacity.
Seed cells are the cornerstone of tissue engineering. Nasal mucosa-derived EMSCs fulfill core therapeutic requirements through accessible sourcing and scalable expansion. Beyond these advantages, their clinical potential for PD treatment extends further. We integrate EMSCs’ biological signatures with developmental insights, including their embryological origin and nasal mucosa anatomy to advance novel PD therapeutics. Their ectoderm-derived features (Nestin+ phenotype, high connexin43, niche-specific paracrine signaling, and strong dopaminergic differentiation) may synergistically combat multiple PD pathological mechanisms. Future investigations should prioritize three key domains: First, mechanistic studies should be conducted employing single-cell RNA sequencing and spatial transcriptomics to elucidate the molecular choreography underlying EMSC-mediated nigrostriatal pathway reconstruction. Second, clinical translation pathways require optimization of cell delivery methods (intranasal or stereotactic delivery), dose escalation protocols, and longitudinal safety assessments in relevant primate models. Third, EMSCs should be combined with tissue engineering for PD treatment. The therapeutic efficacy can be significantly enhanced by integrating them with tissue engineering approaches. These scaffolds can provide a supportive microenvironment, promoting cell survival, differentiation, and functional integration into the host tissue. Combining EMSCs with tissue engineering makes it possible to create a more effective and sustainable treatment strategy for PD, addressing both cell replacement and the restoration of neural circuitry. The high expression of Cx43 in EMSCs likely plays a critical role in tissue engineering. GJs formed by Cx43 not only act as reservoirs but also facilitate drug delivery. Given the innate migratory capacity of MSCs to target damaged sites, these cells internalize sustained-release therapeutic agents for precise transport. Ultimately, drugs are delivered directly to adjacent neurons via GJ-mediated intercellular transfer.
Parallel efforts must establish standardized Good Manufacturing Practice protocols and rigorous quality control metrics for clinical-grade EMSC production. Addressing these challenges will determine whether EMSCs can transition from experimental models to transformative clinical applications in neurodegenerative medicine.
Authors’ Contributions
Z.W. wrote the article and designed figures. Q.Y. participated in the editing of the article. W.D. provided formulation or evolution of overarching research goals and aims. J.B. contributed to article revision. X.X. and J.Y. participated in the management and coordination responsibility for the research activity planning and execution. All authors read and approved the final article.
Footnotes
Acknowledgment
The authors acknowledge Professor Zhang Zhijian for his expertise and assistance regarding the theoretical foundations of germ layers.
Funding Information
This work was supported by the National Natural Science Foundation of China (82173785, 82373820, 81720108030, 81803475) and the National Key Research and Development Project of China (2018YFE0208600).
Data Availability
Data availability is not applicable to this article as no new data were created or analyzed in this study.
Disclosure Statement
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.