
Abstract
As miniature, three-dimensional emulates of individual human organs generated in vitro, organoids are increasingly recognized as complex, humanized models of development, disease, diagnostics, and drug discovery. Organoids exhibit organ-specific architecture, function, and multicellular composition, can be infinitely derived from pluripotent stem cells, and can be further directed toward organoids of the endocrine or exocrine pancreas. Pancreatic endocrine organoids are rapidly redefining diabetes therapies due to their ability to recapitulate glucose-responsive insulin secretion. Conversely, there is less focus on pancreatic exocrine organoids, which possess untapped potential for investigating disorders such as cancer and cystic fibrosis. This review first summarizes human pancreatic organogenesis to contextualize relevant differentiation pathways, then details protocols that guide human pluripotent stem cells through key developmental stages. Methods to enhance cellular maturation and establish higher-performing end products, as well as the therapeutic value of different pancreatic genres, are assessed. Furthermore, crucial gaps are identified, including limited insight into non-beta-endocrine cells, progenitor lineage bias, and off-target differentiation. By chronicling the advancements of all pancreatic organoid classes, the importance of creating more intricate constructs is underscored, which could lead to their broader application.
Impact Statement
This review consolidates foundational work and recent advances in the generation and engineering of pancreatic organoids from human pluripotent stem cells, providing a comprehensive analysis of both endocrine and exocrine cell lineage development. Current differentiation protocols and the associated engineering innovations that improve the maturity of stem cell derivatives are evaluated. The review underscores the growing potential of organoids in disease modeling, drug discovery, and regenerative medicine. Emphasizing the need to expand the functional scope of organoids beyond β-cell-centric models is crucial for future efforts to develop physiologically complex and more clinically relevant pancreatic tissue equivalents.
Introduction
Organoids are three-dimensional (3D) multicellular tissue equivalents recapitulating key structural and functional aspects of complex human organs in vitro. Significant advances have been noted in the human pluripotent stem cell (hPSC)-based generation and cultivation of organoids that hold great promise for modeling detailed organ function and pertinent disease states, screening of drug candidates in high-throughput, and creating tissue equivalents for transplantation.
Tissue modeling using in vitro monolayer cultures has notable drawbacks compared to native 3D tissues, including differences in gene expression,1,2 epigenetics, 3 and cell function. 4 Such shortcomings are symptomatic of fractional cell-to-cell and cell-to-matrix interactions in two-dimensional (2D) configurations that fail to represent complex tissue functions. 5 Characterization of full-organ or multiorgan responses has necessitated the use of animal models, whose accuracy in predicting human physiology is limited by species divergence. Organoids address these issues by more accurately mimicking native tissue architectures and species-specific physiology while offering unprecedented avenues for genetic manipulations and scale-up.
The term organoid was used early on to describe tissue explants and 3D clusters of cells (spheroids). 6 However, the success in establishing models of increasing complexity gives rise to a need for precise nomenclature to define these different systems. As an “organoid,” a tissue must be derived in vitro from stem cells, progenitors, and/or differentiated cells, contain multiple organ-specific cell types, self-assemble into configurations reminiscent of the native organ, and recapitulate one or more aspects of that organ. 6 Analogous to organs of a developing embryo, all cell types arise from stem cells, committing to one of the three germ layers (endoderm, mesoderm, or ectoderm). Thus, organoids are categorized according to their number of constituting germ layers. The simplest structure meriting the organoid distinction is epithelial organoids, which derive from a single germ layer and maintain self-renewal properties. 6 Structures constituting two or more germ layers, termed multitissue organoids, undergo intraorgan self-organization but tend to lose their ability to self-renew as they mature. 6 Lastly, while multiorgan organoids exhibit connectivity among organ domains, very few protocols have achieved this level of complexity. 6
Human PSC-derived pancreatic endocrine organoids (hPSC-PENDOs), which display qualities of human islets, are well-profiled, and their production is becoming increasingly streamlined (Fig. 1). This affords in-depth modeling and the potential to ameliorate core disease phenotypes such as type 1 diabetes, 7 pancreatic cancer,8,9 and pancreatic cystic fibrosis (PCF).10,11 While significant efforts in the field have focused on generating islet cell progeny from hPSCs, few reports document the transition to pancreatic acinar and ductal cells. 10 These exocrine cell subtypes are heavily implicated in pancreatic diseases and thus make for an intriguing target for organoid modeling. Still, they have not received the same research prioritization as pancreatic endocrine cells. Additional challenges arise from the instability of acinar cells in culture, which further impedes the development of protocols for their 3D adaptation and downstream integration into organoid platforms. 12
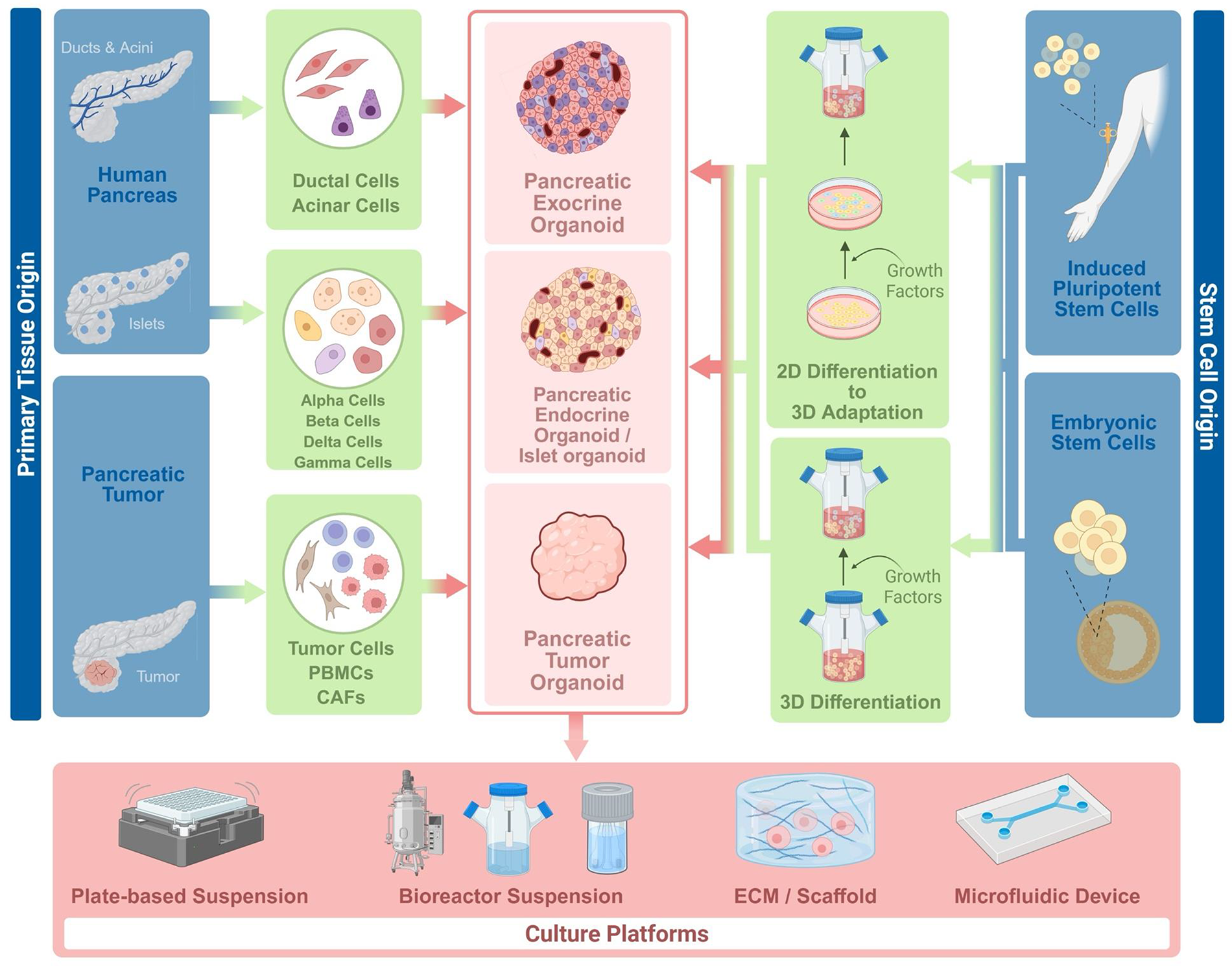
Methods to generate and maintain different categories of pancreatic organoids. The term “growth factors” represents the hormones, small molecules, and growth factors added to cell culture media to direct differentiation. *Not mentioned in culture platforms: air liquid interface systems. PBMCs, peripheral blood mononuclear cells; CAFs, cancer-associated fibroblasts; ECM, extracellular matrix. Created with BioRender.
Commitment to endoderm and formation of pancreatic progenitors (PPs) preludes the separation of endocrine and exocrine cell fates. Therefore, existing protocols for hPSC-PENDOs already document a substantial portion of the exocrine pathway and have been used to inform strategies to produce hPSC-derived pancreatic exocrine organoids (hPSC-PEXOs) (Fig. 1).
This review discusses recent advances in the generation of pancreatic organoids by (i) summarizing pancreas formation and organizing methods for the formation of (ii) endocrine organoids with islet-like features, and (iii) exocrine organoids featuring acinar and ductal cells. Prevailing protocols for PSC-PENDOs and PSC-PEXOs, as well as recent discoveries and integration with engineering approaches, are assessed for each organoid type.
Embryonic Pancreas Development and Functional Aspects of Pancreatic Cells
Pancreatic morphogenesis
Traditional in vitro organoid assembly aims to mimic facets of development, with the ideal end product being a miniaturized version of the organ of interest. The pancreas develops from the endodermal lining of the foregut during embryogenesis. Around the fourth week of gestation, the dorsal and ventral buds form on opposite sides of the developing duodenum and eventually fuse into the central pancreatic duct. The body, tail, and a portion of the head of the pancreas emerge from the dorsal bud, while the ventral bud forms the uncinate process and the remaining part of the head.13,14 Progenitor cells within these regions possess the multipotent capability to become acinar or bipotent trunk progenitors. The latter diverge at the next stage to give rise to either ductal or endocrine cells. 15
Functional aspects of pancreatic cells
Endocrine and exocrine cells work in tandem to perform major pancreatic functions. While the five secretory islet cell types of the endocrine compartment are tasked primarily with sustaining glycemic balance, the exocrine pancreas contains acinar and ductal cells that secrete enzymes for nutrient absorption during digestion.16,17
Major advances have been noted over the past decade in stem cell differentiation toward pancreatic endocrine cell fates due to the central role of insulin-producing beta cells (β-cells) in blood glucose homeostasis and relevant pathologies such as diabetes. Such advances will lead to the reproducible generation of mass quantities of therapeutically beneficial endocrine cells through differentiation protocols reenacting aspects of pancreas formation. 7 A critical quality attribute of hPSC-derived β-cells is their capacity to sense blood glucose levels and secrete appropriate amounts of insulin. These β-cells constitute approximately 70% of human islets,18–20 whereas glucagon-secreting alpha cells (α-cells) 21 are the second most common cell type at 20%. While insulin inhibits glucagon secretion and lowers blood glucose by stimulating glucose uptake by muscle, liver, and adipose cells, α-cells release glucagon to trigger glycogenolysis and gluconeogenesis in the liver, reversing hypoglycemia. 21 These cells also produce acetylcholine to sensitize β-cells to hyperglycemic environments and glutamate to uphold a feedback loop that halts glucagon release in response to hypoglycemia. 21 The functional interactions between the α-cells and β-cells highlight the importance of multicellular models.
Aside from β- and α-cells, islets contain δ-cells releasing somatostatin, γ-cells secreting pancreatic polypeptide, and ε-cells generating ghrelin. Somatostatin-producing δ-cells comprise almost 10% of the human islet. Their hormone is an inhibitor of glucagon and insulin release,20–22 upholding homeostatic nutrient control under normal conditions. 23 The γ-cells represent approximately 5% of the human islet and are primarily associated with gastrointestinal activity.20,24 There is no clear anatomical segregation of the various endocrine cell types within the human islet. Instead, the cytoarchitecture denotes a random patterning of cells along blood vessels configured in 50–250 µm spherical clusters.20,21 Despite their importance, the islets of Langerhans only comprise about 2% of the adult human pancreatic mass. 23
Conversely, acinar and ductal cells work synergistically in the exocrine compartment of the pancreas to facilitate nutrient digestion. The distinguishing properties of these cells are acquired during pancreas morphogenesis and their localization in the pancreas “tip” or the “trunk.” Cells in the “tip” of the branching epithelium adopt an acinar fate, whereas those in the “trunk” turn into ductal or endocrine cells. 13 Groups of acinar cells, termed acini, and ductal cell structures dominate the exocrine compartment, making up 98% of organ weight. 14 Acinar cells aid in nutrient digestion through the secretion of enzymes in the pancreatic juice, catalyzing the breakdown of proteins, carbohydrates, and lipids. A network of ductal cells modifies and channels the pancreatic juice to the small intestine.13–15
Pancreatic Endocrine Organoids
PENDOs differentiation pathway
While β-cell supplementation can be achieved through intraportal vein infusion of cadaveric islets, the limited availability, chronic therapeutic cell loss, need for immunosuppression, and inferior quality of these cellular products hamper the wide-scale application of this approach.25,26 In vitro generation of human islets for transplantation is thus a primary driving force behind advances in 3D pancreatic endocrine differentiation.25,27,28 Besides serving as a source of islet cells, pancreatic organoids can serve as biomimetic materials for studying relevant biological questions, including mechanisms contributing to diabetes during islet development. 19 Such models exhibit superior cytoarchitectural homology to native human islets over traditional rat and mouse islet models, given the interspecies differences in intra-islet cell distribution and organization.20,21
To this end, hPSC-PENDOs are amenable to genetic engineering, allowing for scalable disease modeling and drug screening. 18 Protocols for hPSC-PENDO generation capitalize on developmental biology to manipulate specific signaling pathways responsible for driving cell fate along relevant lineages.29,30 The expression of stage-specific markers facilitates the evaluation of the progress and quality of differentiation. Stem cells are coaxed toward definitive endoderm (DE) by induction of nodal and canonical Wnt signaling,31,32 by Activin A and Wnt-activating ligands or small molecules7,15,33 (Fig. 2). DE cells subsequently enter the foregut endoderm/posterior foregut/gut tube endoderm (GTE) stage in response to an increase in fibroblast growth factor (FGF)7,33,34,36 and Wnt signaling33,36 (Fig. 2). Simultaneous suppression of bone morphogenic protein (BMP) (e.g., by LDN193189 [LDN]) and sonic hedgehog (Shh) signaling (e.g., by Sant17,34,36) prevents off-target induction into liver and lung tissues37,38 (Fig. 2). Vitamin C is often added at this stage to preserve cell viability and suppress premature Neurogenin 3 (NEUROG3) expression, which could otherwise prompt untimely endocrine specification reducing the efficiency of directed specification. 34 Once in the GTE stage, cells can transition into PPs capable of becoming either exocrine or endocrine cells. Additionally, fibroblast growth factor 7 (FGF7; also referred to as keratinocyte growth factor or KGF), BMP-inhibiting LDN, retinoic acid (RA), and a protein kinase C (PKC) activator such as phorbol 12,13-dibutyrate (PdBU) have been applied to convert GTE cells to PP.7,34,39
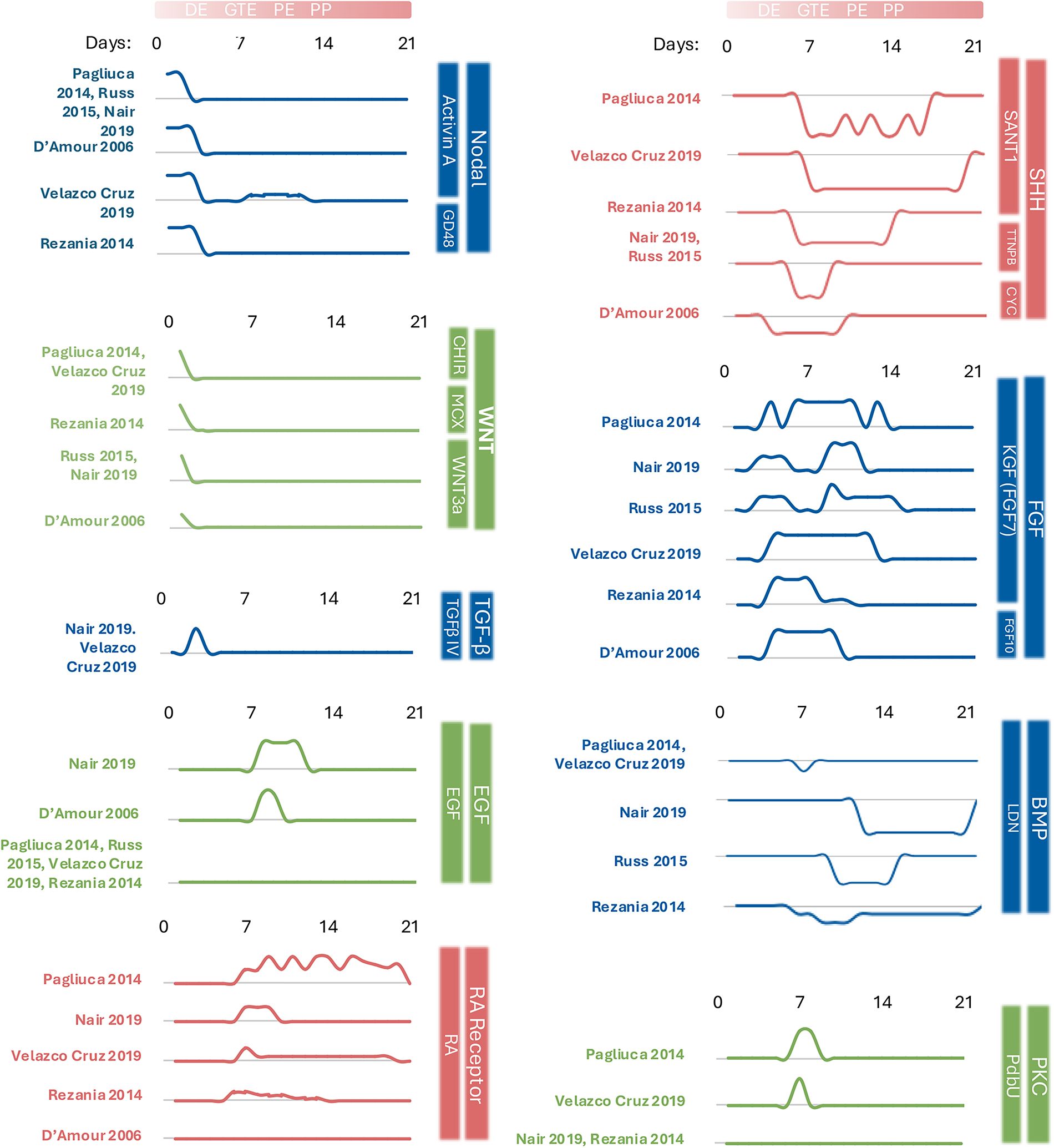
Temporal dynamics of exogenous modulators are used to regulate key developmental signaling pathways during the directed differentiation of human pluripotent stem cells toward pancreatic progenitors. Line plots represent the presence and relative concentration of each factor, normalized to the highest reported value of that factor, across differentiation days (0–21) as described in published protocols (Pagliuca 2014, 7 Rezania 2014, 34 Russ 2015, 33 Nair 2019, 27 Velazco Cruz 2019, 35 D’Amour 2006 32 ). Each modulator is grouped by the developmental signaling pathway it targets (e.g., nodal, WNT, SHH, FGF, RA receptor). The gradient line at the top denotes the approximate timing of each developmental milestone (e.g., DE, GTE, PE, PP), although it should be noted that some protocols have different timing and nomenclature for these stages. This figure highlights both shared and divergent timing strategies across studies. CHIR, CHIR99021; EGF, epidermal growth factor; KGF, keratinocyte growth factor, also known as fibroblast growth factor 7; LDN, lDN-193189; PdBU, Phorbol 12,13 dibutyrate; RA, retinoic acid; SANT1, Sonic hedgehog pathway antagonist 1; TGFβ-IV, transforming growth factor beta isoform IV; Wnt3a, Wingless type MMTV integration site family, member 3A.
PP cells express PDX1, and subsequent induction of NKX6.1 promotes specification to endocrine and ductal cells but not acinar cells.33,40 The initiation of endocrine commitment is predominantly suggested by the expression of NEUROG3, which silences the Notch pathway.29,41 Shh suppression is maintained with Sant1, and Alk5i is introduced 7 to inhibit the TGF-β receptor Alk5 and increase expression of MAFA, a specific β-cell marker. 30 Factors such as thyroid hormone T3 (also increases MAFA expression), XXI, Betacellulin, and Heparin are sometimes incorporated, 7 although their presence and concentrations vary considerably among protocols. Differentiation toward final islet cell types thereafter involves a reduced set of factors, including thyroid hormone T3 and RA.7,30 Dosage and exposure duration of these factors must be tailored to each hPSC line.
State-of-the-art protocols
The signaling pathways detailed above were codified by several keystone publications focusing primarily on β-cell generation from hPSCs.7,27,32,33,35,42 In early works, differentiations typically yielded mixtures of polyhormonal cells which, in some cases, reverse hyperglycemia upon transplantation in diabetic rodents.31,32,43 Detailed time-wise differentiation from PSC to definite PPs 44 and scalable PP maintenance in suspension45,46 have been reported, leading to the production of endocrine derivatives capable of ameliorating diabetes upon transplantation in mice.7,33,34 Further transcriptional characterizations 42 and technical refinements27,35,44 outlined the developmental roadmap from PSC to PP for the reproducible generation of glucose-responsive β-cells.
In most studies, the PSC specification to pancreatic cells skews toward glucose-responsive β-cells.7,27,32,34,35,42 These involve adding soluble factors to amplify the expression of the hallmark transcription factors PDX1 and NKX6.1, underlying insulin-producing phenotypes and enhanced β-cell proliferation, respectively.7,47 The fine-tuning of media additives in differentiation protocols has partially addressed the presence of immature phenotypes. Temporal optimization of TGF-β signaling, specifically through the use of Alk5i after PP formation, equips PSC-derived β-cells with a biphasic insulin secretion pattern (∼150 μIU/μg within the first 10 min after a glucose ramp from 2 mM to 20 mM, followed by a 30–40 min second-phase plateau of ∼50 μIU/μg). 35 While such changes improve cell performance, the resulting β-cells exhibit an immature phenotype with low glucose sensitivity and minimal insulin production compared to human islets, most likely due to the exclusion of islet architecture, other islet cells, and associated interactions.19,27,35,48 Islet organoids featuring all endocrine cell types will be useful for precision disease modeling and diabetes therapies. Recent efforts to consolidate foundational protocols and identify key pathways for addressing the generation of immature islet cells are described below.
Recent additions
In recent years, emphasis has been placed on the maturation of hPSC-derived islet cells.27,28,49 Maturation refers to the adaptive genetic and environmental programming that specializes a cell for specific tasks, such as glucose-dependent glucagon secretion by α-cells. Barsby 2022 50 compiled leading techniques,7,27,35,42 creating a single protocol that is further optimized by a cell aggregation step, driven by the hypothesis that the absence of cell clustering hinders cell maturation. This is aligned with the observation that endocrine cells initially scatter across the epithelium before aggregating into islet precursors and attaining functional maturity during pancreatogenesis. 50 To this end, PSC differentiation to posterior foregut may take place in adherent culture followed by aggregation (e.g., in microwells) for PP cells and a final adaptation to 3D suspension for formation of immature islets with ∼80% PDX1+/NKX6.1+ cells. Cellular diversity is enriched by a 6-week maturation period with the addition of an aurora kinase inhibitor and omission of Alk5i. This raises monohormonal glucagon-producing cells from 5% to 40–50% and maintains insulin- and somatostatin-secreting populations at 40% and 4%, respectively. These hPSC-PENDOs display comparable cytoarchitectural organization to primary adult islets, with coalescing INS+ and GCG+ cells, and β-like cells containing dense-core insulin granules. Accordingly, the insulin content of these organoids surges fourfold during 3 weeks of maturation, as illustrated by glucose-stimulated insulin secretion (GSIS). 28
Maturation is also influenced by the activity of signaling cascades, including noncanonical Wnt, as suggested by transcriptional analysis of hPSC-PENDOs cocultured with fibroblast and endothelial cells. In WNT4-treated organoids, genes associated with mitochondrial function are upregulated, revealing increased oxidative metabolism and GSIS response and maintenance of glucose homeostasis in humanized diabetic mice for over 6 weeks. These findings point to noncanonical Wnt as contributing to the final maturation step of hPSC-PENDO, specifically when pancreatic tissue develops amid endothelial lining and vasculature. 25
Optimizing the PENDOs microenvironment
While attention has largely centered on soluble factors for directed differentiation, extracellular matrix (ECM) remodeling is also a determinant of cell specification and organ formation. Consequently, scaffold design is becoming increasingly relevant in organoid cultivation, providing a way for structural fine-tuning of the microenvironment. Encapsulation, for example, can be leveraged to expose cells to additional biological cues and mechanical stimuli that are absent in the culture media.51,52 This combination is proven effective in supporting physiologically relevant proportions, spatial distribution, viability, and the maturity of cell types within islet organoids. Hydrogel-based scaffolds protect from external shear and allow the diffusion of oxygen and nutrients.53,54 Organoids encapsulated by alginate hydrogels during later stages of differentiation toward PSC-PENDOs contain higher proportions of active secretory cells, 55 possibly due to confinement-induced integrin signaling.55,56 Collagen-based scaffolds bolster the viability and maturity of PSC-PENDOs.51,57 When PSC-PENDOs are cultured in Matrigel-collagen composites, they exhibit elevated stage-specific markers of DE (SOX17) and PE (PDX1, NEUROG3), as well as 5- and 4.3-fold increases of β-cell maturity markers INS and GLUT2, respectively. 51
Platforms specifically for organoid production attempt to merge the mechanical cues and migration aptitude of different scaffolding materials to precisely control organoid size, heterogeneity, architecture, and functional maturation. A contender in this space is Amikagel, a hydrogel comprised of mechanically stiff amikacin hydrate and nonadhesive polyethyleneglycol diglycidyl (PEGDE), facilitating organoid formation. PSCs seeded at different stages in pancreatic formation (PSC, DE, and PP) on Amikagels of various PEGDE/amikacin monomer ratios were evaluated on their rate of aggregation and islet cell maturity. The PEGDE/Amikacin ratio 3:1 supported spontaneous aggregation at a controllable size determined by initial cell seeding density, likely due to the presence of hydroxyl groups. When further coaxed to mature islet phenotypes, qPCR and flow cytometry showed a 17,000-fold increase in INS1 gene expression, marking highly mature β-cells as opposed to Matrigel substratum control, and a 7,000-fold increase in PDX1 gene expression. The use of Amikagel allowed for precise control over cell migration, including the integration of endothelial cells to introduce internal vasculature. Although Amikagel primed spheroids composed of PSC-derived PPs for islet-specific maturation, integration of the polymer into differentiation was stage-sensitive, requiring the cell surface changes that occur during differentiation from DE to PP (likely that of increasing E-cadherin and NCAM genes).54,55
The inclusion of various ECM types in the environment of a hydrogel scaffold is the subject of PSC-to-islet specification studies. Protein-based51,58 or ECM-derived matrices 59 remain concentrated within the capsule, supplying a continuously accessible source of nutrients and biological factors that sustain high viability, reduce hypoxic stress, and improve the differentiation outcome. 60 Such scaffolds protect against external stressors, offer basic mechanical cues, 56 and improve cell viability and differentiation outcomes,51,56 but do not offer the whole gamut of developmental cues necessary for PSC differentiation and maturation. However, ECMs made of decellularized pancreas have been applied independently as coatings on cell culture plastics or in conjunction with 3D scaffolds to create an islet-specific biomimetic microenvironment.57,59,61 Cells were cultured as monolayers on a matrix derived from decellularized rat pancreas (dpECM) and Matrigel or Matrigel-only coated surface until the PP state, before being transferred to ultra-low attachment plates to induce aggregation for further transition to endocrine cells. The islet-like clusters derived on dpECM-Matrigel substrate exhibit cell distribution and organization similar to native islets. The β-cell MAFA and α-cell MAFB are detected by immunocytochemistry in the nuclei of PSC-islets grown on dpECM. In a dpECM-Matrigel substrate ratio of 2:1, over 10% of the cell population is glucagon+/MAFB+, about 30% is C-peptide+/MAFA+, and about 5% is SST+ and pancreatic polypeptide−, as determined by quantitative immunofluorescence. The ratio of insulin secreted at 20 mM and 2 mM of glucose is almost two-fold higher for PSC-islets from dpECM-Matrigel cultures compared to those from Matrigel-only cultures and comparable to the response by native human islets. These findings point to additional biochemical signals supplied by pancreatic ECM to support differentiation, maturation, and improved cellular diversity. 59
Integration of pancreatic dpECM into islet-specific 3D bioprinted niches recreates aspects of the spatial architecture and perivascular ECM, improving the structural organization and functional maturation of hPSC-PENDOs. 61 Successes with dpECM implementation have led to increased efforts to characterize tissue-specific cues for further optimization of the PENDO protocol. Proteomic analysis of decellularized rat and porcine pancreas identified Collagen Type 2 (COL2) as an enhancer of PDX1 and NKX6.1 expression via modulation of Wnt/β-catenin signaling during differentiation from hPSCs. The growth of PSC-PENDOs on a Matrigel substrate enriched with COL2 from dpECM accordingly elevates their insulin and glucagon stimulation indices (ratio of insulin secretion from organoids at 20 mM to 2 mM glucose, shown as mean ± SD) to 5.4 ± 2.7 and 2.7 ± 1.3, respectively, indicating more functionally mature, glucose-responsive PENDOs. 57
Although not traditionally classified as scaffolds, microfluidic platforms are used to modulate the microenvironment of cultured cells through the perfusion of media in microsize channels, delivering oxygen, nutrients, or other factors to cells or organoids while removing waste.62,63 Microfluidics were employed in PSC-PENDO differentiation, with polydimethylsiloxane (PDMS) microwells supporting the perfusion of media, embryoid body formation, and maturation of heterogeneous islets. 64 Based on imaging using the cell-permeable Ca2+ indicator dye Fluo-4 AM, perfused organoids exhibited higher glucose sensitivity that augmented Ca2+ oscillations. Static organoid culture controls secreted significantly less insulin (average of <3 µU/mL) than organoids on chips (>3 µU/mL). Correspondingly, relative mRNA expression of insulin and glucagon was over 40 times that of static-cultured organoids, suggesting improved cellular maturity attributed to the dynamic culture conditions. 64 Microfluidic chips are also considered high-throughput tools for quality control of islet functionality before transplantation. They can be used to detect and report subtle variations in islet response when paired with a suitable functional assay. 65 Thus, microfluidic chip technologies hold promise as tools for assessing the functional quality of PSC-islets.
Pancreatic Exocrine Organoids
PEXOs differentiation pathway
Pathologies linked to extraislet pancreatic cells, such as pancreatic ductal adenocarcinoma (PDAC), underscore the need for models of the exocrine pancreas. However, research on PEXO generation is lagging that of endocrine counterparts due to the strong impetus from diabetes-focused studies and the intrinsic challenges of sustaining acinar cell viability and identity ex vivo. Adaptation of primary acinar cells to 3D culture 12 is challenging. The cells also retain increased plasticity that makes them prone to differentiation, mainly into ductal cells. 66 This trait is exacerbated in vitro in both monolayer 67 and suspension cultures. 68 Exocrine cells prepared from human pancreases (≈63.1% acinar and ≈34.6% ductal) and cultured for 1 week on adherent 24-well plates showed uniform loss of acinar biomarkers, including amylase and chymotrypsin, while ductal cell markers were preserved. Genetic tracing using a Cre-lox-based system was used to exclusively label acinar cells with enhanced green fluorescent protein (EGFP) and trace their fate over time, confirming the adoption of a ductal cell phenotype by acinar cells. 67 This was also assessed in acinar spheroids using a green fluorescent-conjugated lectin to label and track acinar cells over time. This demonstrated dedifferentiation into cells expressing embryonic progenitor markers (e.g., CD142) and pancreatic progenitor markers (PDX1, SOX9), signatures that are atypical of mature acinar cells. 68 While PSC-derived acinar cells are challenging to maintain their enzyme-secreting functionality and remain fully differentiated in vitro, 69 they are generally more robust and have been successfully expanded.10,70,71 The adaptation of methods originally discovered in mouse PSCs—specifically the combination of inducing factors FGF7, GLP-1, and RA—has shown promise toward generating functional acinar cells. When implemented into a four-stage protocol for human embryoid bodies, stable expression of mature acinar genes for amylase and elastase 1 was identified through reverse transcription (RT)-PCR analysis, and digestive enzyme proteins amylase, elastase, and chymotrypsin were confirmed by immunostaining. Functional evaluation showed the prolongation of α-amylase release between ≈ 40 and 50 U/L across 8 days, with a decrease to ≈ 25 U/L on day 15. These data were consistent with electron micrograph images showing secretory granules, indicating the presence of mature acinar cells. 71
The PP state represents a fork in the developmental roadmap where cross-antagonistic expression of NKX6.1 and PTF1A is observed. NKX6.1 expression becomes exclusive to ductal cells and endocrine progeny of the trunk domain of the developing pancreas, while PTF1A is linked to the emergence of acinar cells at the “tip” domain.10,40,70,72 Similar gradients occur with GATA4 and SOX9, with GATA4 becoming increasingly restricted to the acinar cell compartment, while SOX9 commits to trunk domain progenitors with a slight delay behind NKX6.1.8,29 Unique to the ductal cell lineage is the continued activation of the Notch signaling pathway, the deactivation of which contributes to the specification to the acinar cell fate or the divergence of endocrine precursors from the bipotent trunk progenitor.8,13 Early addition of nicotinamide, FGF10, and epidermal growth factor (EGF) promotes ductal specification, 8 while factors promoting the conversion to acinar cells have not been thoroughly investigated.
Early efforts to grow organoids comprising exocrine cells were motivated by a need to model pancreatic cancer pathogenesis. While traditional in vitro cancer models have utilized patient-harvested tumor cells, PSCs were used to derive aggregates of PPs harboring cancer-related mutations. Although useful for studying phenotypic changes between wild-type and cancerous clones in early stages, PSC-derived cells were not coaxed past the PP stage. 70 These aggregates were termed organoids, but similar to previous tumor models, they did not possess the cellular diversity, cytoarchitectural, and organ-specific functional attributes exhibited by bona fide organoids.
Results on generating acinar and ductal cells from PSCs have been limited.8,10 Cells have been cultured as monolayers up to the PP stage and either embedded in Matrigel or transferred to ECM-free liquid suspension plates (Fig. 1). At this stage, exocrine cell determination is induced by adding nicotinamide and FGF2 to the basal media, yielding acinar and ductal cells, which rapidly assemble into structures with cyst-like acinar/ductal morphology and functional resemblance to the fetal human pancreas. The hPSC-derived ducts display microvilli and tight junctions, while secretory granules and lumen formation are noted in acinar progeny. Initial monolayer differentiation yielded 70% PDX1+/NKX6.1+ PPs, and subsequent adaptation to 3D culture produced PEXOs of relevant acinar/ductal composite architecture (34%±15% acinar and 61% ± 19% ductal cells) and expression of chymotrypsin C (exocrine), SOX9, and cytokeratin 19 (ductal). Acinar-exclusive biomarkers were not included, although acinar-like structures that were positive for amylases were observed. Upon orthotopic transplantation into immunodeficient mice, organoids formed pancreatic ducts and acinar tissue. 10
PEXOs disease modeling applications
A range of pancreas pathologies stands to benefit from the development of pancreatic organoid-based technologies. These pancreatic disorders implicate the endocrine (e.g., diabetes) or exocrine pancreas (e.g., chronic pancreatitis [CP], Alagille syndrome [ALGS], 73 Schwachman–Diamond syndrome [SDS], 74 and PCF 75 ). Pancreatic exocrine insufficiency, characterized by aberrant secretion or activity of digestive enzymes leading to maldigestion and nutritional deficiency, can also occur independently of the aforementioned diseases as a byproduct of extrapancreatic health conditions such as inflammatory bowel syndrome, celiac disease, HIV, surgery, and aging and lifestyle. 76
Research on PDAC, which is a highly aggressive cancer of 3% survivability, can benefit from organoid-based systems featuring differentiated ductal cells9,70 (Fig. 3). Challenges in identifying the cellular origins of pancreatic oncogenesis, as well as the lack of suitable disease models and early detection methods contribute to the poor survival outcomes of PDAC70,77 Organoid-like tumor models composed of biopsied tissue are regularly deployed to study PDAC. 78 However, exocrine organoids of PSC descent differ from traditional tumor models by putting greater emphasis on offering insight into disease progression in intermediate cell types. Starting from a stem cell state also offers a higher threshold for genetic modification and, consequently, greater control over the mutations being modeled to mimic specific genetic predispositions. Although it is generally accepted that PDAC can manifest in both acini and ducts, pathophysiology differs dramatically between cellular origin, making a major goal of PDAC research to trace precursor presentation.8,9,77,79 PSC-derived ductal cells have established complex ductal structures upon transplantation into murine models and act as vectors for carcinogenic mutations. To this end, PSC-derived ducts harboring PDAC-affiliated mutations in KRAS, a gene that relays signals for cell growth and division, develop lesions. 8 Moderate exocrine lineage specification was induced in progenitor organoids with a 3.5-fold increase in SOX9 expression and a decline of NEUROG3. Immunocytochemical analysis revealed the mutation TP53R175H driving SOX9 into the cytosol of exocrine cells. The aberrant localization of SOX9, which was later confirmed in primary PDAC samples, 70 suggests a potential prognostic indication of shifted gene expression during tumor development and malignancy. Of note, there are currently no hPSC-PEXOs of mature acinar and ductal cell lineages being used as models for PDAC.
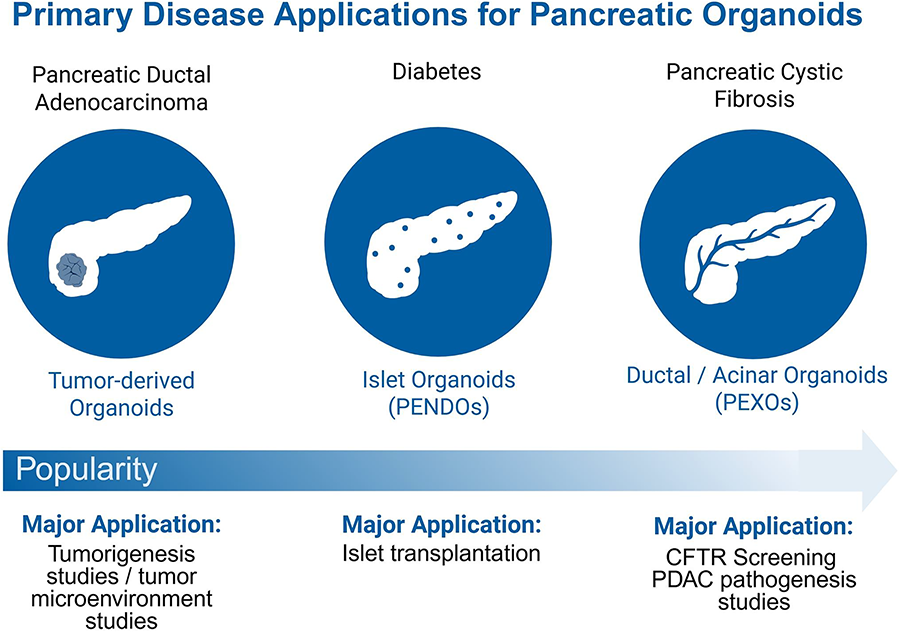
Primary disease applications of pancreatic organoid subtypes. The popularity of using pancreatic organoids as research tools varies depending on the disease target. A well-established application is the use of tumor-derived organoids to model PDAC and other pancreatic cancers, enabling studies of tumor progression. Islet organoids, specifically those derived from stem cells, have recently gained traction in the diabetes space as potential transplants to regulate insulin production autonomously. Ductal and acinar organoids represent the exocrine pancreas and support studies of genetic diseases such as cystic fibrosis, pancreatitis, as well as PDAC. Created with BioRender.
Cystic fibrosis (CF) is a genetic disorder in which mutations on the cystic fibrosis transmembrane conductance regulator (CFTR) gene lead to a faulty chloride channel needed for fluid homeostasis in organ epithelia, causing a buildup of viscous mucus in the pulmonary and gastrointestinal tracts.10,11 Organoids of relevant organs can be programmed with mutation-specific defects to model aspects of CF and facilitate the development of therapeutics 11 (Fig. 3). Pancreatic exocrine differentiation has been employed to develop a scalable disease model for CF. Keratinocytes from the hair follicles of CF patients were reprogrammed into PSCs, which were then subjected to pancreatic exocrine differentiation. A fluorescent dye quenched by the presence of chloride elucidated functional differences in chloride content within the organoid lumen, where CFTR impairment caused a reduction in chloride levels compared to the normal chloride influx of wild-type organoids. Gene expression profiling confirmed that resulting organoids natively retained the disease-specific genotypes, which was attributed to the loss of CFTR function. This allowed authors to pursue a novel screening methodology to determine the treatment eligibility of CF patients. 10
The exocrine pancreas is implicated in several orphan diseases such as SDS74,80 and ALGS73,81 where high variability in presentation and a small demographic make diagnosis and treatment challenging. Acinar and ductal cells differentiated from the reprogrammed PSCs of patients with SDS—a disease characterized by bone marrow failure, pancreatic insufficiency, and skeletal abnormalities—formed organized tube-like structures between acinar colonies in monolayer cultures. In contrast to transgene-corrected cells, disease variants exhibited progressive loss of exocrine tissue, aberrant morphology, and increased protease activity that could be partially reversed upon treatment with a panel of common protease inhibitors. 80 PSC-derived 3D models for ALGS, a disease caused by mutations in the Notch signaling pathway, are currently exclusive to the liver 82 despite 40% of cases exhibiting pancreatic insufficiency. 73
Summary
The high therapeutic relevance of islet-like cells to potentially cure diabetes, a condition impacting over 10% of the global adult population, 83 has motivated large-scale generation of the insulin-producing β-cells that exist within the islet complex. In this pursuit, researchers drew from knowledge of pancreas development and prior work on PSC differentiation to identify a sequential orchestration of signaling pathways and factors that derive 3D islet-like organoids from PSCs. The pathway toward PSC-PENDOs began with procedures committing PSCs to endoderm,31,32,43 then to an endocrine and exocrine progenitor state44–46 before achieving functional islet (mainly β-) cells.7,27,34,35,42 Among all protocols, PP quality is hallmarked by strong coexpression of transcription factors PDX1 and NKX6.1. Functionality testing hinges on PENDOs' ability to respond to varying levels of glucose exposure with an appropriate balance of insulin, as tested through GSIS and commonly measured through ELISA. Similarly, the detection of glucagon and somatostatin is used to identify α-cell and δ-cell populations, respectively. Scaffold-based strategies are helpful for advancing spatial organization, viability, and maturation of PSC-PENDOs. Hydrogel encapsulation and ECM integration are the most popular scaffolding equipment due to ease of fabrication and customization- factors that consequently increase their scale-up potential. However, the reliance of these systems on Matrigel poses a significant challenge due to its xenogeneic origin and undefined composition, which renders it unsuitable for clinical use.
The stepwise trajectory from PSC to PEXO is significantly less researched than that of PSC-PENDOs. Still, protocols defining the PSC > DE > GTE > PP pathway have laid a groundwork where optimization of the PP-to-exocrine juncture can be established.10,11 PEXOs offer a modeling tool for studying diseases originating or implicated in the acinar/ductal network of the pancreas and provide a profound understanding of diseases such as SDS that start in early development. 80 The acinar cells' tendency to dedifferentiate ex vivo into progenitors and adopt new cell fates poses a challenge. 69 Although more difficult to culture than ductal-biased tumor models, PSC-PEXOs improve upon acinar cell sensitivity, have a more complex microenvironment, and offer more genetic control for recapitulating specific mutations, providing deeper insight into the diverse origins and development of oncogenic cellular matrices. Even for rare diseases, PSC-PEXOs provide a way to study direct genotype-phenotype relationships and identify areas for therapeutic intervention.
Outlook
While PSC-PENDOs are rapidly defining a new age in diabetes treatment, their window of application remains limited to β-cell-centric protocols that fulfill this demand, leaving their potential as diagnostic tools and more complex models of pancreatic disease relatively unexplored. Alpha cells and δ-cells are often treated as convenient byproducts of β-cell production, and few studies elaborate on their roles. Updated protocols are pushing for elements to enhance the maturation of cellular end products, advocating for the incorporation of islet diversity, endothelial cells, and proper vasculature to provide mature cells with an environment that closely mimics that of the native tissue.15,25,28,50 For example, mouse models suggest a critical role of endothelial cells in relaying signals in the embryo during endoderm induction, but few protocols have investigated ways to implement this in PSC-PENDO generation. 15 Efforts to mimic the mechanics of pancreas organogenesis have already revealed an increase in cellular maturation,27,28,50 calling for more studies to incorporate mechanotransduction in PSC-PENDOs culturing practices. More accurate recreations of embryonic development are key to revealing additional signals that can make PENDOs of improved maturity and greater functional utility. 25
Developing protocols for exocrine cell specification from originally endocrine cellfocused practices assumes that PP cells do not acquire commitment bias and are true progenitors that can commit to endocrine and exocrine lineage after identical molecular and growth factor additions. 40 Fewer protocols delineating the PP to PEXO trajectory leave possibilities of PP commitment bias uninvestigated. It should also be noted that the yield of heterogeneous or immature cell products in PEXOs (and PENDOs), which increases the risk of spontaneous differentiation into potentially harmful off-target cell types after transplantation, 84 may hinder their use in an implantation setting. Addressing this concern will require boosting the differentiation efficiency 80 in conjunction with more stringent purification/negative selection methods to eliminate undesired cells. Extrapancreatic lineage differentiations can be prominent, and purifying cells of off-targets at the PP stage is a crucial optimization that has yet to be achieved. 10
To this end, the derivation of acinar cells with terminal functionality from PSCs must first be optimized. PEXO generation should follow the organizational principles of endocrine protocols (controlled signaling, staged differentiation, optimized microenvironment) but accounting for exocrine-specific biology. Adjacent strategies to curate organoids of collective pancreatic parenchyma can be examined for biomarkers used to define PPs capable of traversing acinar, ductal, and endocrine trajectories. A high level of glycoprotein-2 (GP2) in PPs has been linked to such multipotent capability. 85 Compound testing to produce and purify GP2-rich populations has optimized stage-specific media conditions for generating progenitors of a greater trilineage potential, which contribute to the formation of more functionally and morphologically relevant organoids. 86 This is promising for the extension of PEXOs to rare pathogenic niches where therapeutic research is limited by patient scarcity and poorly understood disease mechanisms.
Authors’ Contributions
S.K.: Conceptualization, visualization, writing—original draft, and writing—review and editing. E.S.T.: Writing—review and editing.
Footnotes
Funding Information
Funding support has been provided by the National Science Foundation (CBET-1951104, CBET-2015849, CBET-2326510) to E.S.T.
Disclosure Statement
The authors declare no competing interests.