
Abstract
Periodontal ligament (PDL) is a thin connective tissue that connects the tooth to the bony socket and plays a crucial role in the regeneration and maintenance of homeostasis of periodontal tissues by supplying stem/progenitor cells. Induced pluripotent stem cells (iPSCs) are highly anticipated in regenerative medicine because of their differentiation potential into a wide variety of cell types. In this study, we investigated the effects of humoral factors on iPSC differentiation by culturing iPSCs in the presence of PDL cell-derived culture supernatants. Changes in gene expression were analyzed using quantitative real-time PCR, reverse–transcription PCR, and RNA sequencing. The marker protein expression on the cell surface was assessed using flow cytometry. Periodontal regeneration was verified by microcomputed tomography and histomorphological observation in a periodontal defect model using male F344/NJcl-rnu/rnu rats. When iPSCs were cultured in the PDL culture supernatant, some cells formed clumps, and spindle-shaped cells grew out from them. Upon passaging, spindle cells increased further, and by the fifth passage, these cells occupied the entire culture. These cells (iPS-PDLs) expressed genes such as periostin and Asporin/PLAP1, and their comprehensive gene expression patterns resembled those of PDL cells. iPS-PDL cells exhibited a cell surface antigen profile of CD90+, CD73+, CD105+, CD44+, CD29+, CD14−, CD34−, CD45−, and CD19− and differentiation potential into osteoblasts, adipocytes, and chondrocytes. Transplantation of iPS-PDLs into rat periodontal defects increased the height of newly formed bone and enhanced periodontal tissue regeneration after 4 weeks. Our results showed that iPSCs differentiated into cells with properties similar to those of PDL cells in the presence of humoral factors of cultured PDL cells. Additionally, the transplantation of iPS-PDL cells into periodontal defects induces periodontal tissue regeneration. These findings provide valuable insights for developing novel periodontal regenerative therapies using iPSCs.
Impact Statement
Periodontal disease remains a major cause of tooth loss, with current treatments offering limited regenerative potential. This study presents a novel approach to differentiate induced pluripotent stem (iPS) cells into periodontal ligament-like cells (iPS-PDL cells) using PDL-derived humoral factors. iPS-PDL cells exhibit key regenerative properties, including osteogenic and chondrogenic differentiation potential and the ability to enhance periodontal tissue regeneration in vivo. This research represents the first demonstration of iPS cell-derived PDL-like cells contributing to tissue regeneration, highlighting their potential as a cell-based therapeutic strategy for periodontal diseases and marking a significant advancement in dental regenerative medicine.
Keywords
Introduction
Periodontal disease is a chronic inflammatory condition of the periodontal tissues caused by plaque bacteria. 1 Prolonged inflammatory response leads to tissue destruction.2,3 Current regenerative therapies using growth factors and bioactive agents are clinically applied; however, they are limited in their applicability and the amount of regenerated tissue, highlighting the need for new approaches to periodontal tissue regeneration.4,5
The periodontal ligament (PDL) is a soft tissue between the tooth and alveolar bone. The PDL plays important roles in maintaining homeostasis, facilitating wound healing, and regenerating periodontal tissues by supplying stem cells.6–8 In the wound healing process of periodontal tissues, it has long been accepted that the type of wound healing largely depends on the initial cell type that migrates to the wound site.6,8 Specifically, root resorption occurs when gingival connective tissue-derived cells migrate first, long junctional epithelium attachment forms when epithelial cells migrate, ankylosis develops when bone-derived cells migrate, and proper periodontal regeneration (cementum-PDL-bone structure) occurs when PDL-derived cells are present. This concept suggests that periodontal regeneration can be facilitated only when PDL cells are induced at the wound site. Clinically, this has been demonstrated using a physical barrier membrane to prevent the rapid migration of epithelial cells, thereby promoting PDL cell proliferation and migration. 9 Based on this principle, guided tissue regeneration (GTR) was developed in the 1980s as the first regenerative periodontal therapy. 10 Furthermore, transplantation of in vitro-cultured PDL cells has been explored as a more direct approach, with clinical studies confirming periodontal regeneration through cultured PDL cell transplantation.11–14 This method holds promise as a novel regenerative therapy, as conventional approaches like GTR are limited in the extent of tissue regeneration. However, the culture of PDL cells requires tooth extraction, posing challenges for securing an adequate cell source for transplantation.
Induced pluripotent stem cells (iPSCs) are reprogrammed cells that gain pluripotency through gene introduction. Takahashi and Yamanaka generated iPSCs from skin fibroblasts through the introduction of Oct3/4, Sox2, Klf4, and c-Myc. 15 iPSCs can theoretically differentiate into almost any type of cells in the body, which makes them promising candidates for regenerative medicine. iPSC-derived cells have been used to treat a wide variety of diseases.16–19 The applications of iPSCs in regenerative medicine are anticipated to increase in the coming years.
Efforts to induce iPSC differentiation into promising cell types for cell-based treatment involve mimicking developmental processes in vitro using small molecules and growth factors or culturing iPSCs with target cells or in conditioned media (CM) derived from these cells. Chu et al. demonstrated that iPSCs differentiated into cardiomyocytes when cocultured with cardiac cells. 20 Similarly, Kitazawa et al. demonstrated that iPSCs cultured in dorsal root ganglion-CM differentiated into motor and sensory neurons. 21 Although coculture or CM-based differentiation of iPSCs is considered less precise due to the presence of unknown factors responsible for differentiation, it offers a simpler approach, as it does not require meticulous optimization of factors, concentrations, or timing. To date, it remains unclear whether iPSCs can be induced to differentiate into PDL-like cells using CM derived from PDL cells (PDL-CM), and whether these differentiated cells possess the capacity to regenerate periodontal tissues.
This study aimed to investigate changes in iPSCs cultured in PDL-CM, identify the cell types into which iPSCs differentiate, and evaluate whether these differentiated cells can contribute to periodontal regeneration, thus exploring the potential of iPSCs in periodontal regenerative therapies.
Materials and Methods
Cell culture
iPSCs (HPS0381 line) were obtained from the RIKEN BioResource Center (Tsukuba, Japan) and cultured using StemFit AK02N (Ajinomoto, Tokyo, Japan). Periodontal ligament fibroblasts (PDL cells) from Lonza (Basel, Switzerland) were cultured in α-modified minimal essential medium (αMEM; Thermo Fisher, Waltham, MA) supplemented with 15% fetal bovine serum (FBS). After 3 days, CM from PDL cells was collected, centrifuged, and filtered twice (0.22 µm pore; Kurabo, Osaka, Japan). PDL-CM was stored at −20°C until use, and 15% FBS was added for use in iPSC differentiation.
Induction of iPS-PDL differentiation
PDL-CM was applied to iPSCs, and the medium was replaced daily with a fresh PDL-CM. After 4 weeks, cells were passaged to 10 cm culture dishes with type 1collagen coating (AGC Techno Glass, Shizuoka, Japan). Cells were subsequently subcultured at a 1:2 ratio every 3–5 days using αMEM containing 15% FBS.
Flow cytometry analysis
Cells were detached and fixed with 4% paraformaldehyde solution (PFA) in PBS. The cells were blocked with 5× Blocking One (Nacalai Tesque). Afterward, 2 × 105 cells were reacted at 4°C for 30 min with various antibodies diluted in ×20 Blocking One. The following antibodies were used along with their isotype controls: CD19, CD105, CD29, CD44, CD73, CD45, TRA-1-81, TRA-1-60R, SSEA4 (BioLegend), CD90, CD14 (Abcam), and CD34 (BD Biosciences). Antigen expression levels were determined using FACSVerse flow cytometer (BD Biosciences) and FlowJo software (BD Biosciences).
Reverse–transcription PCR
Total RNA was extracted using NucleoSpin RNA (Macherey Nagel, Düren, Germany). Complementary DNA was constructed from 0.5 to 1 µg of total RNA using ReverTra Ace® qPCR RT Master Mix (Toyobo, Osaka, Japan). PCR reactions were performed using Quick Taq® HS DyeMix. The PCR products were separated by agarose gel electrophoresis and visualized using a gel imaging system (ChemiDoc XRS Plus; Bio-Rad, Hercules, CA). Sequences of used primers are listed in Supplementary Table S1, and original gel images are shown in Supplementary Figs. S5, S6, S7 and S8.
Quantitative real-time PCR
Quantitative real-time PCR was performed using THUNDERBIRD® Next SYBR™ qPCR Mix (Toyobo) and the StepOnePlus Real-Time PCR system (Thermo Fisher). Gene expression levels were calculated using the ΔΔCT method with GAPDH as an internal control. Sequences of used primers are listed in Supplementary Table S1.
Induction of differentiation into osteoblasts, chondrocytes, and adipocytes
Cells were cultured for 3 weeks in αMEM supplemented with 50 µg/mL
Rat periodontal tissue defect model and cell transplantation
This study protocol was approved prior to the experiment by the Osaka Dental University Institutional Animal Care and Use Committee (approval number #23-04012) and the DNA Recombination Research Committee (approval number #73). All procedures adhered to the ethical guidelines of the Science Council of Japan and ARRIVE guidelines (Supplementary Data). The animal experiments were conducted at the Osaka Dental University Animal Facility between June 2023 and March 2024. The rats were housed two per cage, and cages were kept at a constant temperature and humidity. Immunodeficient rats were used to prevent immune rejection of transplanted human iPS-derived cells. Sixteen male immunodeficient rats (F344/NJcl-rnu/rnu, 8–10 weeks old) (Shimizu Laboratory Supplies, Kyoto, Japan) were anesthetized via intraperitoneal injection of medetomidine (Zenoaq, Fukushima, Japan), midazolam (Astellas Pharma Inc., Tokyo, Japan), and butorphanol (Meiji Seika Pharma Co., Ltd., Tokyo, Japan). A periodontal defect was created at the mesial root of the maxillary first molar in rats, following previous reports, as a well-established model of periodontal tissue defects.22,23 An incision of approximately 3 mm was made in the gingiva at the mesial side of the maxillary first molar, and a mucoperiosteal flap was elevated. A standardized alveolar bone defect measuring 1 mm buccolingually and 2.5 mm in depth was then created at the mesial root surface of the maxillary first molar by removing the alveolar bone, cementum, and dentin under a microscope using a dental bur. One defect was created per animal, minimizing gingival incisions and avoiding damage to surrounding tissues. iPS-PDL cells were cultured for 3 days in αMEM supplemented with 50 µg/mL
RNA-sequencing analysis
Total RNA was extracted from iPSCs, three donor-derived PDL cells, and three independently differentiated iPS-PDL cell lines (#1–3) using NucleoSpin RNA. The RNA samples were processed and sent to Aelieff (Tokyo, Japan) for RNA sequencing. Sequencing was performed on NovaSeq 6000 system (Illumina) with 4 Gb of data per sample, using 150 bp paired-end sequencing. Sequence data were analyzed using principal component analysis (PCA), hierarchical clustering, differential expression analysis, and pathway enrichment.
Karyotyping by G-band method
The iPS-PDLs (#1–3) were cultured to 30% confluency in T25 flasks and transported to Nihon Gene Research Laboratories Inc. (Sendai, Japan). G-band pattern photographs and karyotypes were obtained.
Statistical analysis
All experiments were repeated more than three times to confirm the reproducibility. The representative results were presented in figures. All data are expressed as mean ± standard deviation and were analyzed using GraphPad Prism with Student’s t-test. Statistical significance level was set at p < 0.05.
Results
Differentiation of iPSCs into iPS-PDL cells
iPS cells were maintained in single-cell feeder-free conditions (Supplementary Fig. S1). Figure 1A illustrates the method to induce iPSC differentiation with the PDL-CM. It was observed that various growth factors are contained in PDL-CM, including hepatocyte growth factor (HGF) and vascular endothelial growth factor (VEGF) (Supplementary Fig. S2). The culture medium of iPSCs was changed to PDL-CM and changed daily until day 28, which was followed by five passages. As shown in Figure 1B, iPSCs cultured in PDL-CM initially exhibited a high nucleus-to-cytoplasm ratio. Cell clusters began to form within several days to 2 weeks (Fig. 1C). Between days 14 and 28, spindle-shaped cells began to proliferate around the clusters (Fig. 1D). After passaging of cells, two cell types, spindle-shaped cells and cells with a large cytoplasm, were observed (Fig. 1E). With further passaging, spindle-shaped cells became dominant, and by approximately the fifth passage, nearly all the cells were spindle-shaped (Fig. 1F) and were designated as iPS-PDL cells.
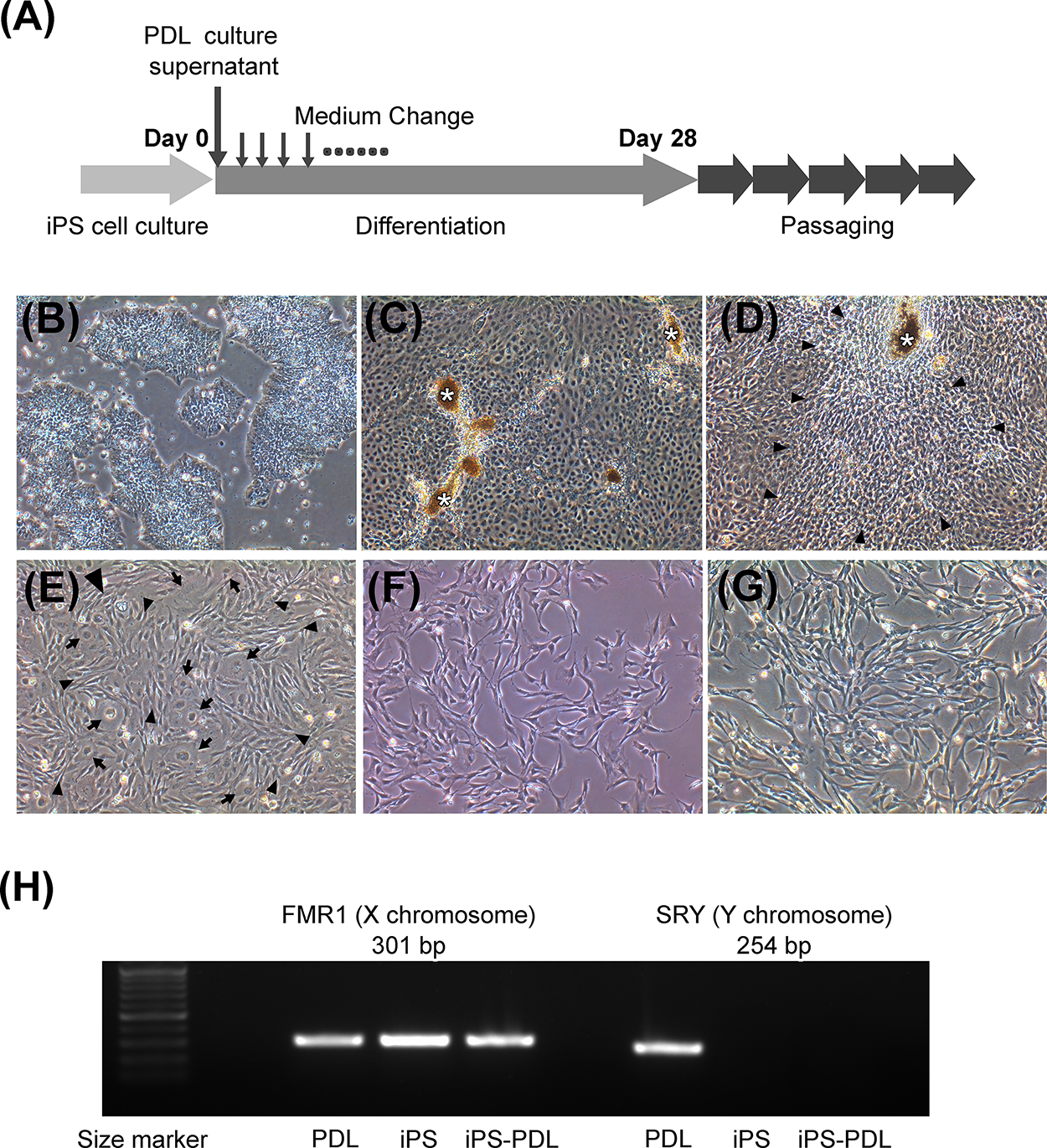
Induction of differentiation of iPSCs into PDL-like cells (iPS-PDL).
To confirm that PDL cell contamination in iPSC culture did not occur through PDL-CM, we used a CM from male-derived PDL to differentiate female iPSCs and analyzed FMR1 (X chromosome) and SRY (Y chromosome) expression levels. The results showed that the iPS-PDL cells expressed FMR1(+) and SRY(−), confirming that they differentiated directly from the iPSCs (Fig. 1G).
Marker gene expression in iPS-PDL cells
We examined the expression of iPS- and PDL-specific genes in iPS-PDL cells. Analysis of the three iPS-PDL cell lines (#1–3) showed no expression of the pluripotency markers Oct4A and Nanog. In contrast, PDL cell-associated genes, including CD73, CD90, TN-C, Periostin, Asporin/PLAP1, COL1A1, CEMP1, aSMA, fibronectin (FN), COL3A1, and Decorin, were expressed in iPS-PDL cells (Fig. 2A and Supplementary Fig. S3). The gene expression profiles of iPS-PDL cells were similar to those of PDL cells but distinct from those of iPSCs. For protein levels, iPS-PDL were positive for PDL-related maker proteins including aSMA and periostin (Supplementary Fig. S4). Temporal analysis revealed that Oct4A expression was lost within 1 week, whereas CD73, Periostin, and COL1A1 expression increased within 1–2 weeks, and Asporin/PLAP1 emerged in the third week (Fig. 2B). These findings show that iPSCs lose pluripotency markers within 7 days and express PDL-associated genes after 3 weeks in the presence of PDL-CM.
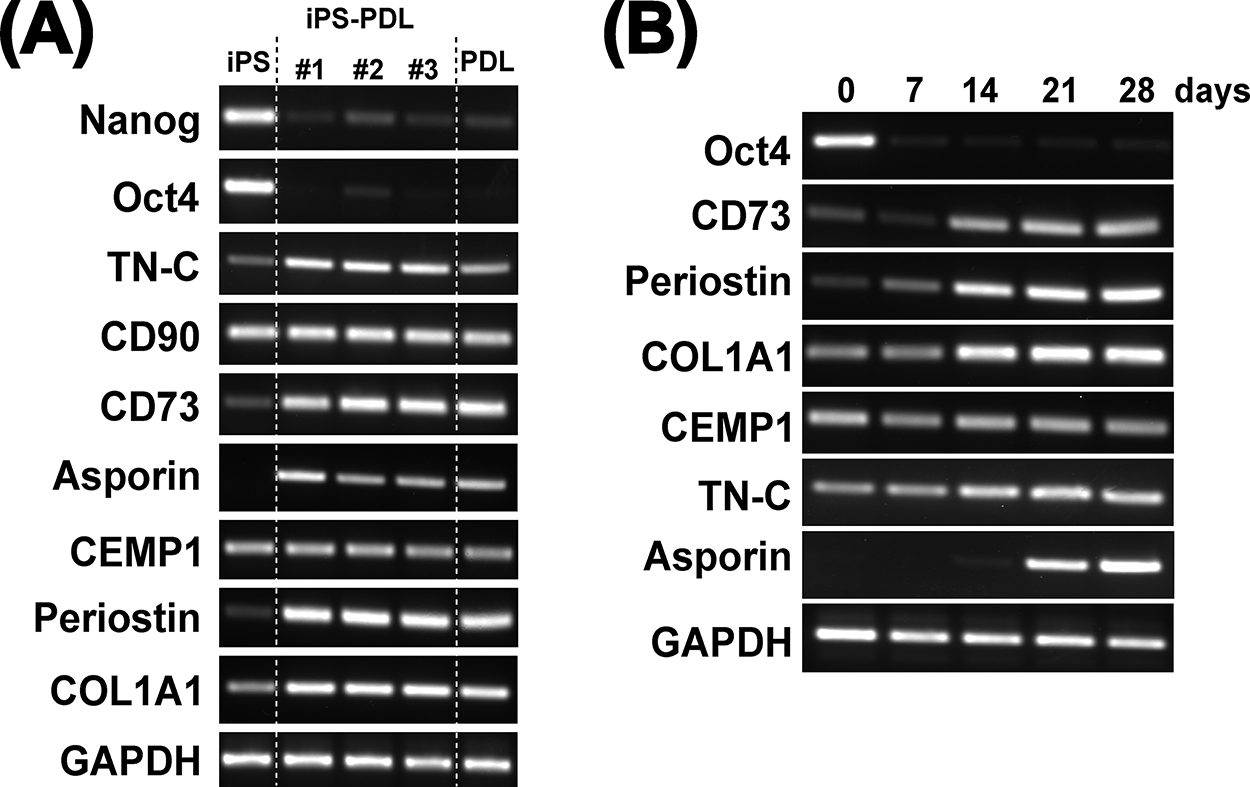
Gene expression in iPS-PDL cells.
Surface marker expression in iPS-PDL cells
We evaluated the cell surface marker expression in iPS-PDLs using flow cytometry. As shown in Figure 3, iPS-PDL cells did not express pluripotency markers (TRA-1-60R, TRA-1-81, and SSEA4) and hematopoietic markers (CD14, CD34, and CD45), but expressed mesenchymal stem cell (MSC) markers (CD73, CD90, and CD105). Cell surface marker expression profile of iPS-PDL was similar to that of primary PDL cells and distinct from that of iPSCs.
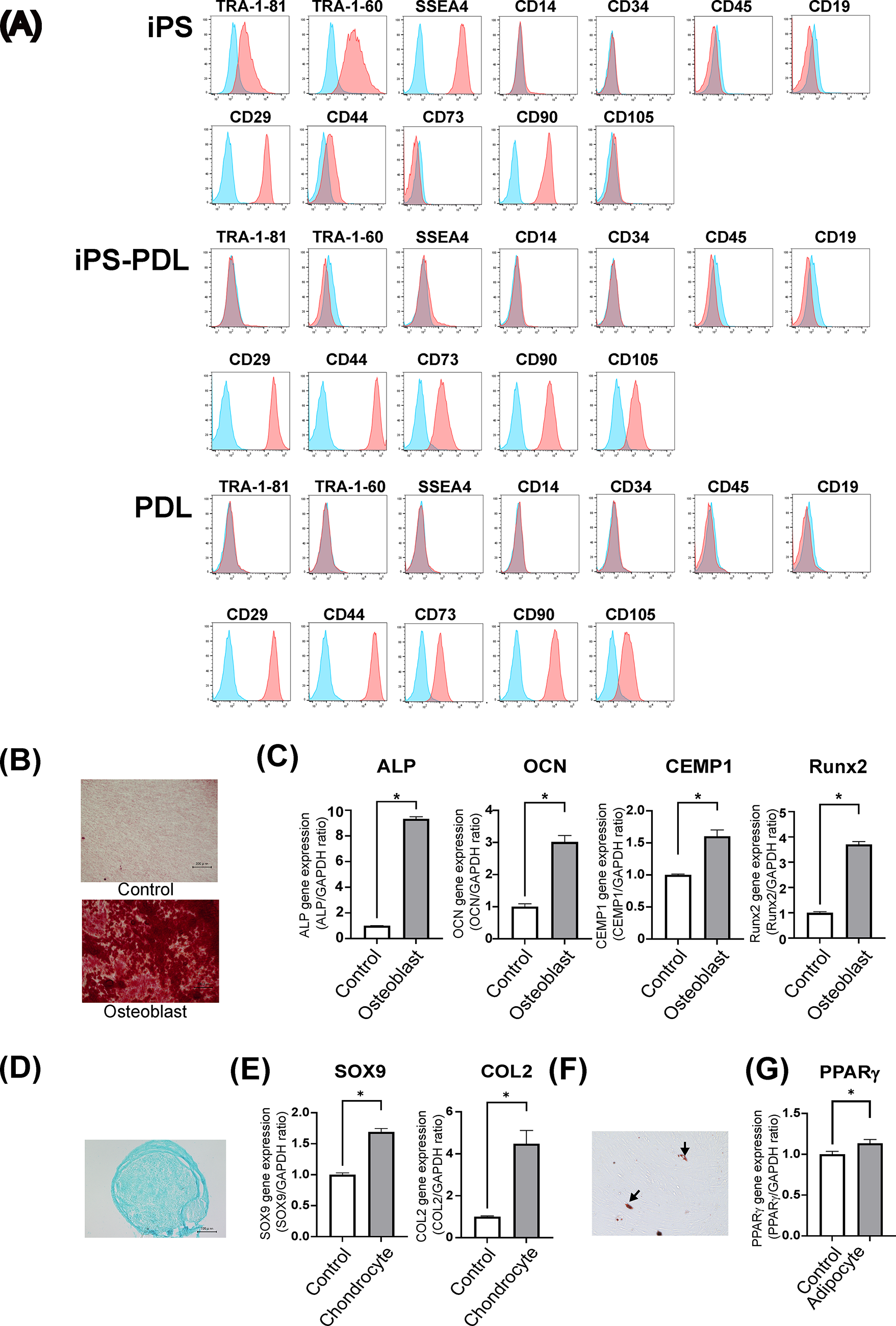
Cell surface marker expression and differentiation potential of iPS-PDL cells. Flow cytometric analysis of cell surface marker expression in iPSCs
Differentiation potential into osteoblasts, adipocytes, and chondrocytes
Given the MSC-like phenotype of iPS-PDLs in flow cytometry, we evaluated their differentiation potential into osteoblasts, chondrocytes, and adipocytes. After osteogenic induction, iPS-PDL cells showed significant matrix mineralization, as confirmed by Alizarin Red staining (Fig. 3A). The osteoblast markers, including ALP, Osteocalcin (OCN), CEMP1, and Runx2, were significantly upregulated (Fig. 3B). Under chondrogenic differentiation condition, pellet culture formed cell aggregates, which was positive for an Alcian Blue, indicative of cartilage (Fig. 3C). The chondrogenic markers, SOX9 and COL2, were significantly upregulated (Fig. 3D). Adipogenic induction produced oil droplet-like structures stained with Oil Red O (Fig. 3E), but their number was limited, and the adipogenic marker PPARγ was slightly increased (Fig. 3F).
Comprehensive gene expression analysis of iPS, iPS-PDL, and PDL cells
We examined the global gene expression profiles of iPS, iPS-PDL, and PDL cells using RNA sequencing. In hierarchical clustering (Fig. 4A), iPS, iPS-PDLs, and PDL initially formed separate clusters within each group, and iPS-PDLs and PDL then formed clusters. Volcano plots highlighted the DEG between cell types (Fig. 4B). Venn diagrams (Fig. 4C) revealed that 10,876 and 10,900 genes were differentially expressed between iPS-PDL versus iPS and iPS-PDL versus PDL cells, respectively, whereas 3896 genes differed between iPS-PDL and PDL cells. PCA confirmed distinct clustering of the three cell types, with iPS-PDL and PDL cells plotted closer together than iPSCs (Fig. 4D). Reactome pathway enrichment analysis (Fig. 4E) identified the top 10 pathways with significant differences. Eight of the top 10 pathways were shared between iPS versus iPS-PDL and iPS versus PDL, but no common pathways were found between PDL versus iPS-PDL and other combinations.
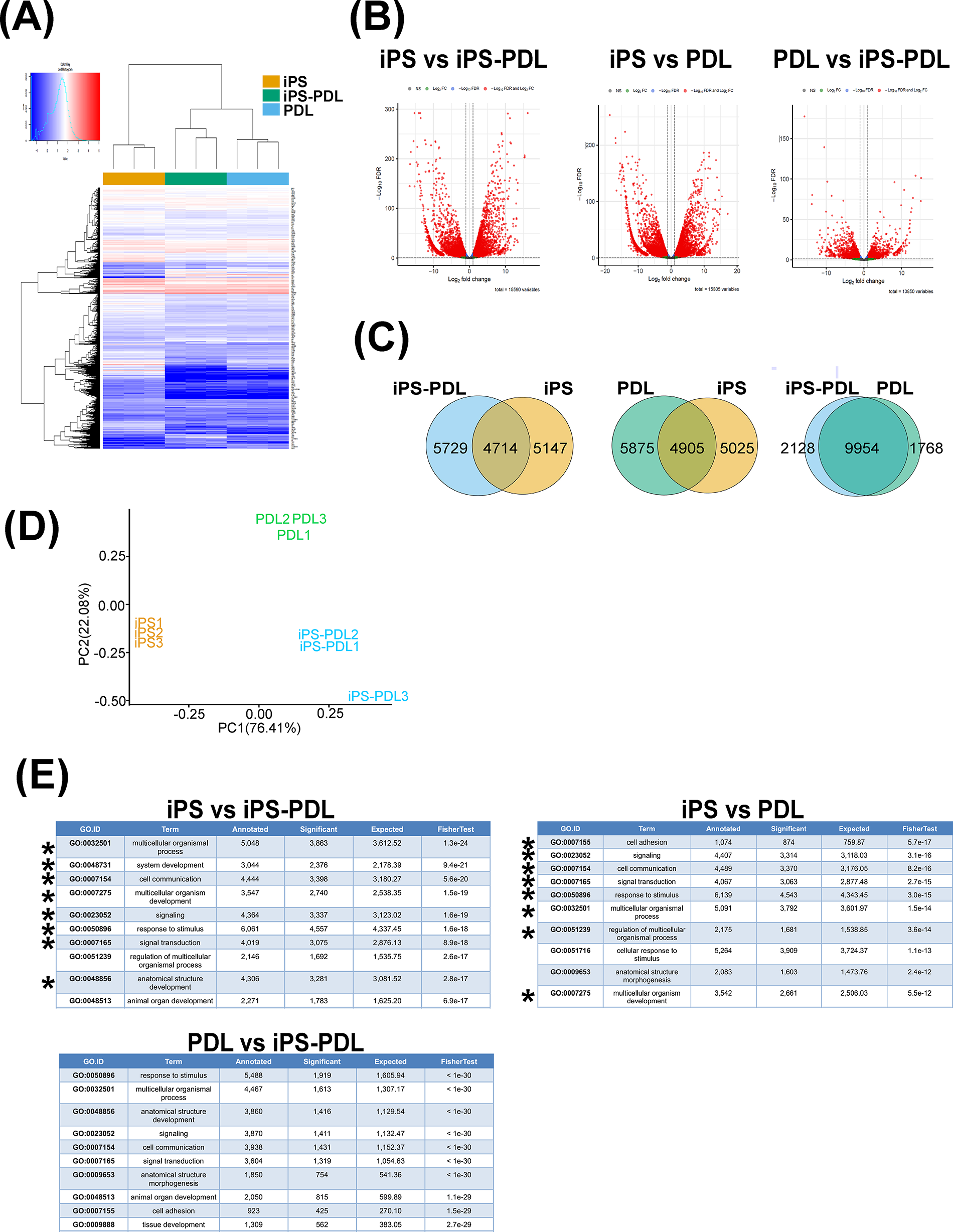
Global gene expression analysis of iPS, iPS-PDL, and PDL by RNA sequencing. RNA-sequencing analysis was performed on iPS, iPS-PDL, and PDL, and gene expression was compared. Results of hierarchical clustering
Transplantation of iPS-PDL cells into periodontal defect model in rats
iPS-PDL cells were transplanted in surgically created periodontal defects in rats to assess their potential for periodontal regeneration (Fig. 5A). Four weeks posttransplantation, micro-CT analysis showed significantly smaller exposed root area in the iPS-PDL group compared with the control group (effect size = 0.92, Fig. 5B and C). Histological analysis showed that the height of the new bone formation, which was the primary outcome of this experiment, was greater in the iPS-PDL group (effect size = 0.94, Fig. 5D and E). HE and azan staining confirmed the formation of new PDLs and alveolar bone in the iPS-PDL group, demonstrating that iPS-PDL transplantation enhanced periodontal regeneration (Fig. 5F).
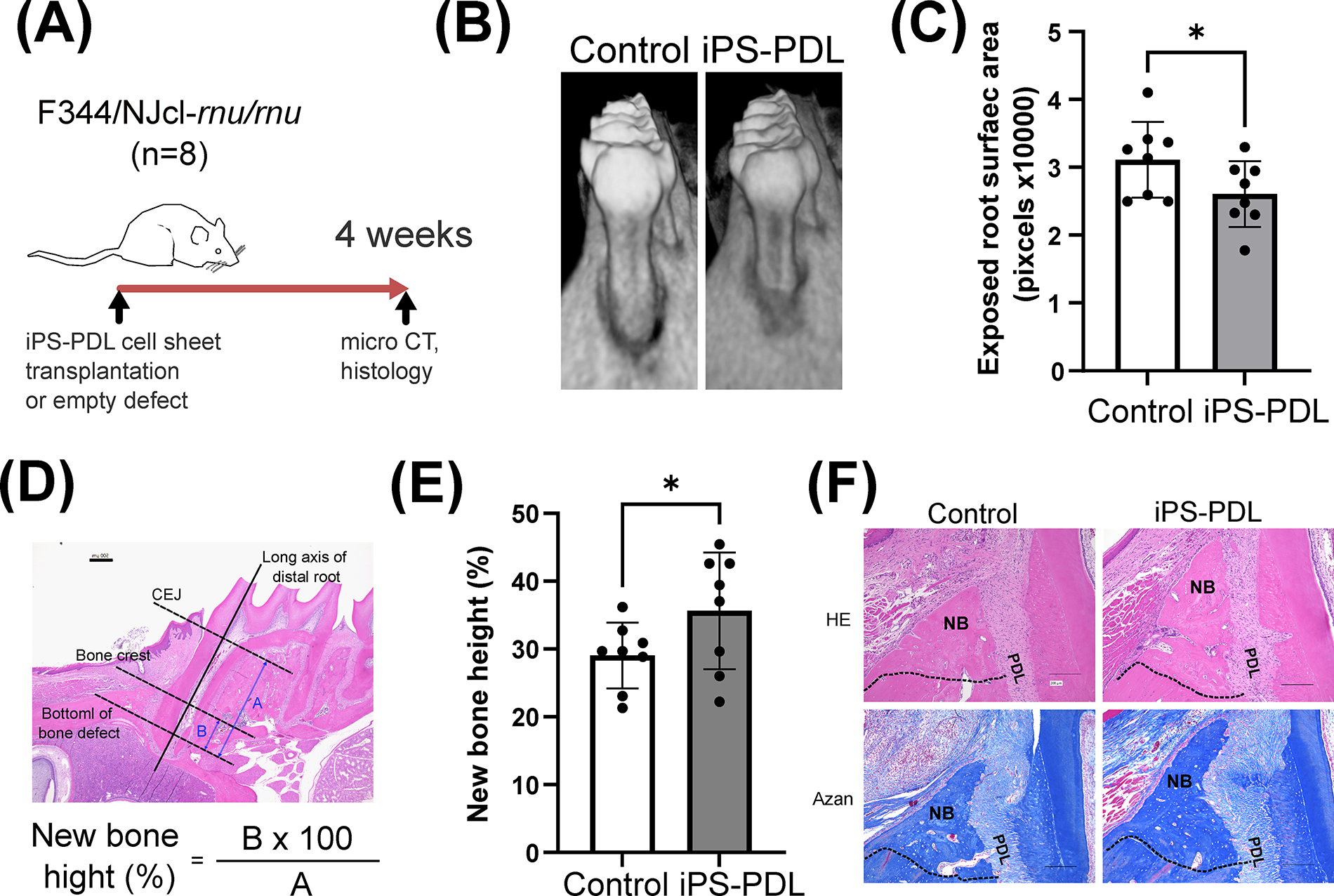
Transplantation of iPS-PDL cells enhances periodontal tissue regeneration in a surgical periodontal defect model. The figure shows an overview of the animal experiment
Karyotyping of iPS-PDL
To test the chromosome stability, karyotyping of the three iPS-PDL lines (#1–3) was performed using G-banding. As shown in Figure 6, additional fragments of unknown origin were observed on chromosomes 4 and 6 in iPS-PDL #2.
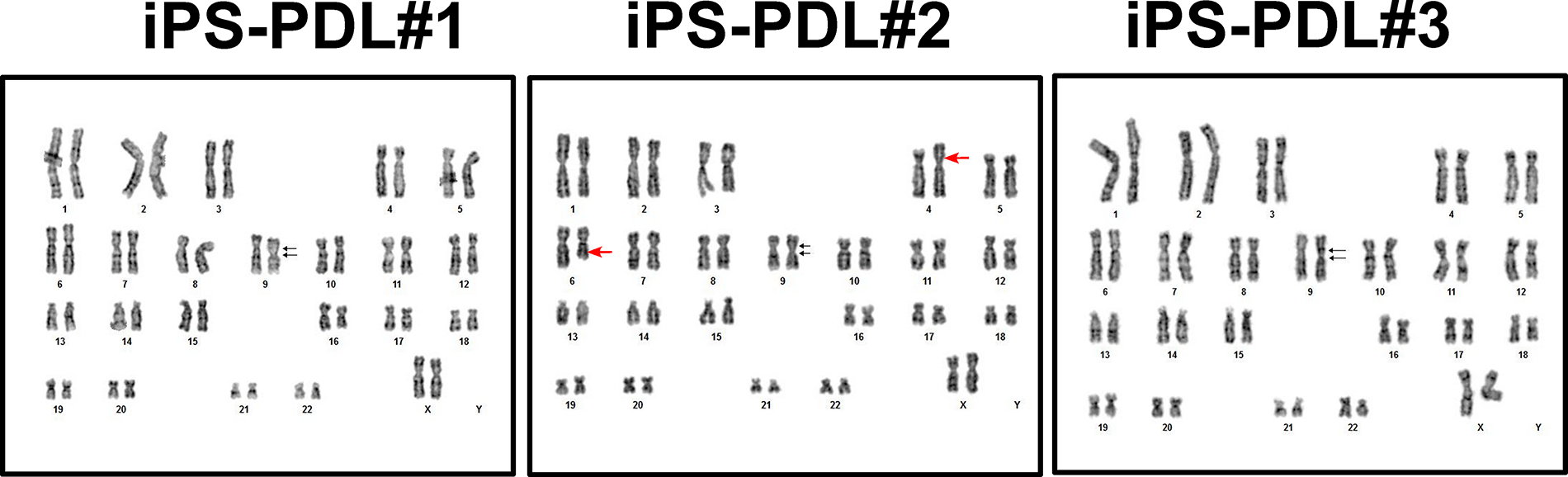
Karyotyping of iPS-PDL cells by G-banding. Karyotyping results for iPS-PDL#1, iPS-PDL#2, and iPS-PDL#3 using the G-banding method are shown. All samples exhibit 46 chromosomes, including sex chromosomes (XX). A pericentric inversion of p12q13 on chromosome 9, which originates from the original iPS cells, is consistently observed in all iPS-PDL lines. Additionally, iPS-PDL#2 shows extra fragments of unknown origin on chromosomes 4 and 6. Black arrows indicate karyotypic mutations. Red arrows highlight the karyotypic change found in iPS-PDL #2.
Discussion
In this study, we examined the effects of culturing iPSCs with CM derived from PDL cells. The differentiated cells expressed various PDL-related genes, and their cell surface antigen expression patterns in flow cytometry analysis, as well as gene expression profiles in RNA sequencing, were similar to those of PDL cells. These findings indicate that iPSCs induced to differentiate with PDL-CM acquire PDL-like properties. RNA-sequencing analysis showed that 82.4% of the genes expressed in iPS-PDL cells were not significantly different from those in primary PDL cells, and 84.9% of the genes expressed in primary PDL cells were not significantly different from those in iPS-PDL cells. PCA further demonstrated that iPS-PDL and PDL cells clustered closely on PC1, indicating a high degree of similarity, although both were clearly distinct from iPSCs. Despite these similarities, iPS-PDL and PDL cells formed separate clusters in the PCA. These results indicate that although iPS-PDL cells are similar to PDL cells, they are not identical. The difference between primary PDL cells and iPS-PDL will require further investigation.
Seo et al. reported that a population of MSC-like cells can be cultured from the PDL, which they named periodontal ligament stem cells (PDLSCs). PDLSCs express MSC markers in vitro; differentiate into osteoblasts/cementoblasts, adipocytes, and collagen-producing cells; and form cementum–PDL-like tissues in vivo. 24 The iPS-PDL cells we induced from iPSCs in this study exhibited similar characteristics to PDLSCs in terms of marker gene expression, cell surface antigen expression patterns, and in vitro differentiation potential. Furthermore, iPS-PDL cells have been shown to promote periodontal tissue regeneration when transplanted into periodontal defects. However, we concluded that iPS-PDL cells more closely resemble PDL cells rather than PDLSCs, as they possess the ability to differentiate into osteoblasts and chondrocytes, but not adipocytes, and do not meet the criteria for MSCs as defined by the International Society for Cellular Therapy. 25 Further studies using PDLSCs or MSCs as controls are required to allow a more direct comparison with iPS-PDLs. Additional functional assays (e.g., differentiation potential into osteoblasts/cementoblasts, response to mechanical stress) are also required to establish their mature PDL-like functionality.
Several studies have attempted to induce the differentiation of iPSCs into PDLSC-like cells. Hamano et al. differentiated iPSCs into neural crest cells and cultured them on the extracellular matrix (ECM) derived from human PDL cells to generate PDLSC-like cells. 26 Sugiura et al. expanded on this finding by showing that PAX9 regulates gene expression and adipogenic differentiation of PDLSC-like cells derived from iPSCs. 27 Wang et al. differentiated iPSCs into neural crest cells and cultured them with FGF8b, FGF2, and BMP4 to produce PDL-MSC-like cells. 19 It is not known to what extent the differentiated iPS-PDL cells in this study are functionally similar to the PDLSC-like cells reported above. Previous reports have not examined the ability of iPSC-derived cells to regenerate periodontal tissue in vivo. By combining the information on iPSC-derived PDLSC-like cells from previous reports with the results of this study, it may be possible to develop a method for differentiation of iPSCs into PDL-composing cells or periodontal tissue regeneration therapy using iPSCs.
The clinical application of iPSCs in regenerative medicine faces significant safety challenges, including the risks of genomic integration of oncogenes or viral vectors as well as tumorigenicity from residual undifferentiated cells.28,29 Our iPSCs were generated from human peripheral blood mononuclear cells using an episomal vector to introduce Oct4, Sox2, Klf4, L-myc, Lin28, and TP53 short hairpin RNAs.30,31 Unlike earlier protocols that used c-myc, this strain substituted L-myc, thereby reducing tumorigenicity. 32 Episomal vectors, which did not integrate into the genomic DNA, were used, and their absence was confirmed. 31 However, one of the three iPS-PDL lines used in this study exhibited chromosomal abnormalities (additional fragments), raising concerns regarding transplantation suitability. Chromosomal aberrations, which often arise during iPS and ES cell culture, may increase the risk of cancer.33,34 To address this issue, shorter differentiation periods and optimized protocols are essential. If abnormalities cannot be avoided, secreted factors such as exosomes from iPS-PDL cells could be explored as safer alternatives for periodontal regeneration.
We did not identify the specific humoral factors within PDL-CM that drive iPSC differentiation. PDL-CM is known to contain a wide variety of bioactive components, including growth factors and cytokines (e.g., VEGF, TGF-β, IL-6, IL-8, IL-10, SDF-1), ECM proteins such as fibronectin and collagen type I, and extracellular vesicles. We also observed that various growth factors were contained in PDL-CM including HGF and VEGF, in this study. These components have been reported to influence stem cell proliferation, differentiation, and tissue regenerative responses.35–38 Thus, its effects are likely mediated by the combined action of multiple molecules rather than a single factor. Future studies employing proteomic profiling and functional blocking of candidate pathways will be necessary to clarify the molecular mechanisms underlying PDL-like differentiation.
This study demonstrated that iPSCs can differentiate into PDL-like cells using PDL-CM and that these cells induce periodontal regeneration upon transplantation. However, this study had several limitations. First, the mechanism of differentiation remains unclear due to the diverse factors present in PDL-CMs. Second, the animal model employed involved young immunodeficient rats with surgically created periodontal defects, which differ from chronic periodontitis in humans. Additionally, iPS-PDL transplantation was performed in immunodeficient rats, which differs substantially from the clinical situation in humans. Therefore, the present findings cannot be directly translated into patient treatment. Last, the optimization of cell transplantation protocols has yet to be addressed. This study involved the use of FBS and xenogeneic cell transplantation, and therefore, establishment of xeno-free culture systems and exploration of clinically compatible supplements such as autologous serum or human platelet lysate will be required for clinical application. Further studies addressing these limitations are necessary to advance the clinical applications of iPS-PDLs in periodontal regeneration.
In conclusion, iPSCs differentiated into iPS-PDL cells using PDL-CM, resulting in PDL-like cells capable of inducing regeneration of periodontal tissue. This method provides a straightforward approach to generating PDL-like cells, addressing cell sourcing challenges for autologous transplantation and advancing research in periodontal regeneration.
Authors’ Contributions
Y.W. contributed to conception, data acquisition, interpretation, writing—original draft, and writing—review and editing. K.I. contributed to conception, design, data acquisition, interpretation, writing—original draft, and writing—review and editing. Y.T. and I.I. contributed to conception, interpretation, writing—original draft, and writing—review and editing. Y.H. contributed to design, data analysis, interpretation, writing—original draft, and writing—review and editing. All authors gave their final approval and agree to be accountable for all aspects of the work.
Footnotes
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.