
Abstract
Bone regeneration remains a significant clinical challenge in conditions such as trauma, osteoporosis, and aging-related bone loss. Recent advances have highlighted the crucial role of extracellular vesicles, especially exosomes, in intercellular signaling pathways that support bone homeostasis and repair. Among their bioactive cargoes, exosomal RNAs—particularly messenger RNAs and microRNAs—have emerged as central regulators of osteogenesis by modulating gene expression, cellular differentiation, and communication within the bone microenvironment. In this review, we provide a comprehensive summary of exosome biology, including their biogenesis, secretion, uptake mechanisms, and RNA cargo characteristics. We critically examine current evidence on how exosomal RNAs influence the molecular mechanisms of bone formation, remodeling, and regeneration under both physiological and pathological conditions such as fractures, diabetes, osteoporosis, and osteoarthritis. Furthermore, we discuss the emerging therapeutic potential of engineered exosomes as RNA delivery systems in bone tissue engineering and regenerative medicine. A better understanding of the functional roles and clinical relevance of exosomal RNAs may pave the way for next-generation, RNA-based therapies in skeletal repair and treatment of bone-related diseases.
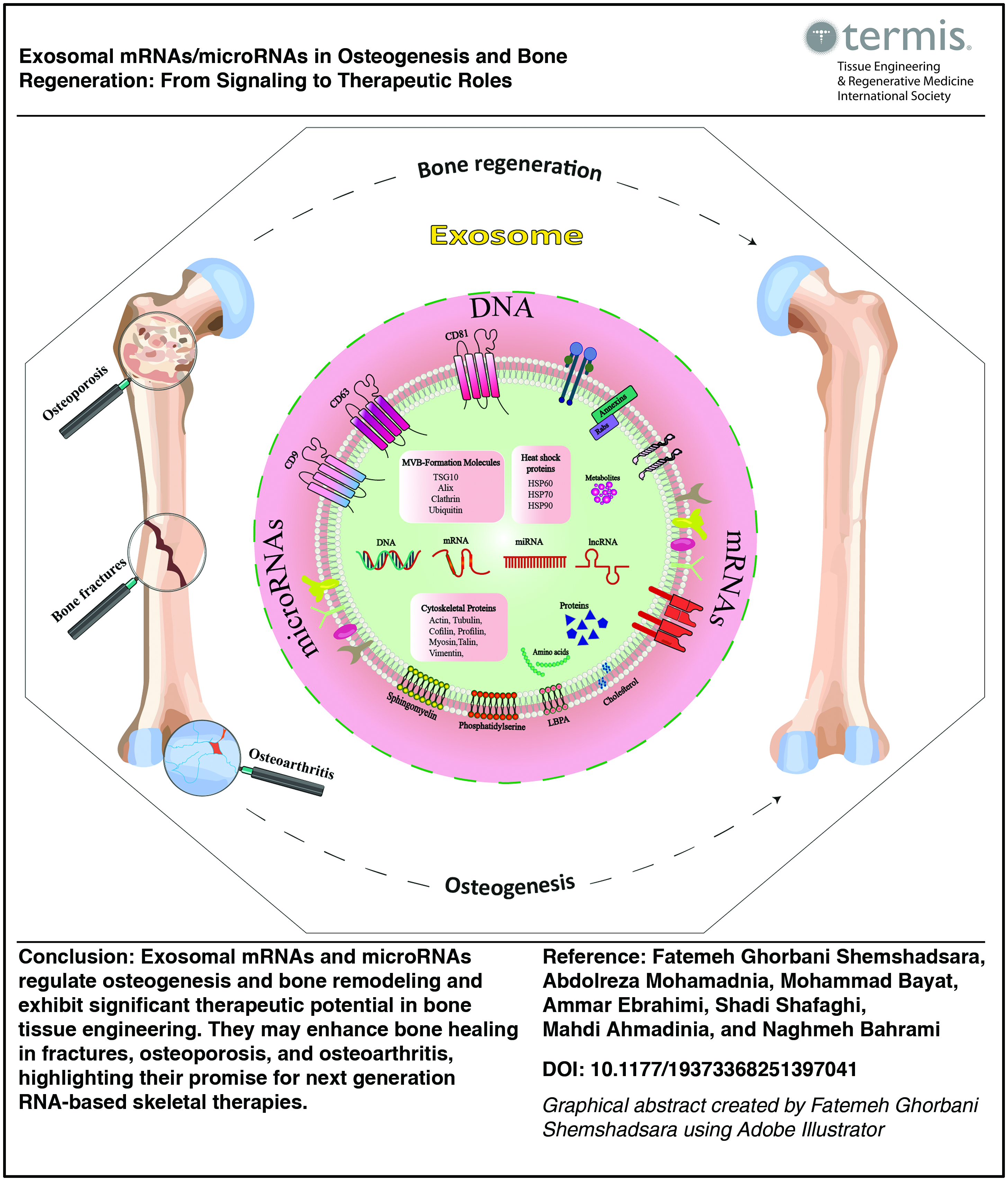
Impact Statement
This review highlights the crucial role of exosomal mRNAs and microRNAs in regulating osteogenesis and bone regeneration. By elucidating the molecular mechanisms and signaling pathways involved, it provides new insights into the potential of exosome-based therapies in bone tissue engineering. This work may accelerate the development of innovative RNA-based regenerative strategies, ultimately improving treatment outcomes for bone diseases and injuries.
Introduction
Bone is a metabolically active tissue with an intrinsic capacity for regeneration and remodeling. 1 These processes are regulated by the coordinated activity of osteoblasts, osteoclasts, osteocytes, and their interactions with the surrounding stromal, endothelial, and immune cells.2,3 However, in pathological conditions such as osteoporosis (OP), large fractures, aging, or chronic inflammation, this regenerative capacity becomes impaired, necessitating therapeutic intervention. 1
Recent research has shifted attention toward the role of extracellular vesicles (EVs), particularly exosomes, in mediating communication between bone cells and modulating bone regeneration. 4 Exosomes are nanosized vesicles secreted by various cell types, enriched with a complex cargo of RNAs, proteins, and lipids. Among them, exosomal RNAs—especially microRNAs (miRNAs) and messenger RNAs (mRNAs)—have garnered significant interest due to their ability to posttranscriptionally regulate gene expression and influence osteogenic differentiation, matrix mineralization, and angiogenesis.2,5 These RNA molecules can influence osteogenic differentiation, matrix synthesis, and angiogenesis by targeting specific signaling pathways in mesenchymal stem cells (MSCs), osteoblasts, and other bone-resident or supporting cells. 4
This review provides an updated overview of exosome biology, encompassing their biogenesis, secretion, uptake mechanisms, and functional cargo profiles. We discuss the emerging mechanistic insights into how exosomal RNAs influence bone formation, remodeling, and regeneration under physiological and pathological conditions such as fractures, OP, osteoarthritis, and diabetes. Furthermore, we highlight the translational potential of exosome-based RNA therapeutics in bone tissue engineering and regenerative medicine. Understanding the dynamic landscape of exosomal RNAs offers a promising avenue for developing next-generation strategies for bone repair and skeletal disease treatment.
Exosome
In 1981, “small EVs” were defined as a subset of microvesicles about 40 nm in diameter that were separated from larger microvesicles between 500 and 1000 nm in studies on normal and cancerous cell cultures with 5-nucleotidase activity. 6 Later research revealed the genesis of these tiny EVs that secrete complexly in mature sheep reticulocytes. Transferrin receptors on 50-nm vesicles produced from mature blood reticulocytes into the extracellular environment are among these findings.7–9 Johnston and colleagues labeled small EVs as exosomes to differentiate them from other forms of EVs. 9 Exosomes are the smallest members of EVs with an average size of 100 nm and a density of 1.13–1.19 g/mL, with a lipid bilayer membrane and spherical shapes that are secreted by a wide variety of cell types, including embryonic cells, endothelial cells, epithelial cells, neuronal cells, immune cells, cancer cells, and stem cells.9,10 Due to their small size and the inherent stability provided by their lipid bilayer membrane, exosomes are readily secreted into various bodily fluids, including urine, blood, saliva, breast milk, lymph, bile, and other physiological fluids, following their synthesis.11–13 These vesicles carry a variety of bioactive molecules essential for cellular communication and function, including diverse classes of proteins, lipids, genetic materials, signaling molecules, and other biologically active compounds.14–16 Subsequent research demonstrated that exosomes play a significant role in regulating the functions and activities of target cells by facilitating the transfer of mRNAs and miRNAs.10,17 Additionally, they are critically involved in immune responses, antigen presentation,18,19 diagnosis, treatment, and progression of various diseases.10,20 So, originally perceived as cellular waste devoid of biological functions, exosomes are now widely recognized for their therapeutic potential and promising applications in regenerative medicine.16,21
Biogenesis of exosomes
Multiple signaling pathways govern the intricate biological process of exosome production, comprising several steps. These processes enhance the diversity of cargo molecules, protein composition, and categories in exosomes. 22 Additionally, they differentiate from other EV kinds (microvesicles and apoptotic bodies) by their endosomal biogenesis. 23 In exosome biogenesis, cells use different endocytosis pathways to internalize materials from the extracellular environment. The process begins with the inward invagination of the plasma membrane, resulting in the formation of early endosomes.24,25 These early endosomes undergo a maturation sequence, during which a secondary inward budding event leads to the creation of intraluminal vesicles (ILVs), transforming them into multivesicular bodies (MVBs). These MVBs can follow distinct pathways: they may either fuse with the plasma membrane to release exosomes into the extracellular space or interact with lysosomes or autophagosomes for degradation and recycling.26–28 The entire process of ILV formation and MVB trafficking is tightly orchestrated by protein complexes such as the ESCRT (endosomal sorting complex required for transport) machinery, as well as by ESCRT-independent routes22,29,30 (Fig. 1).
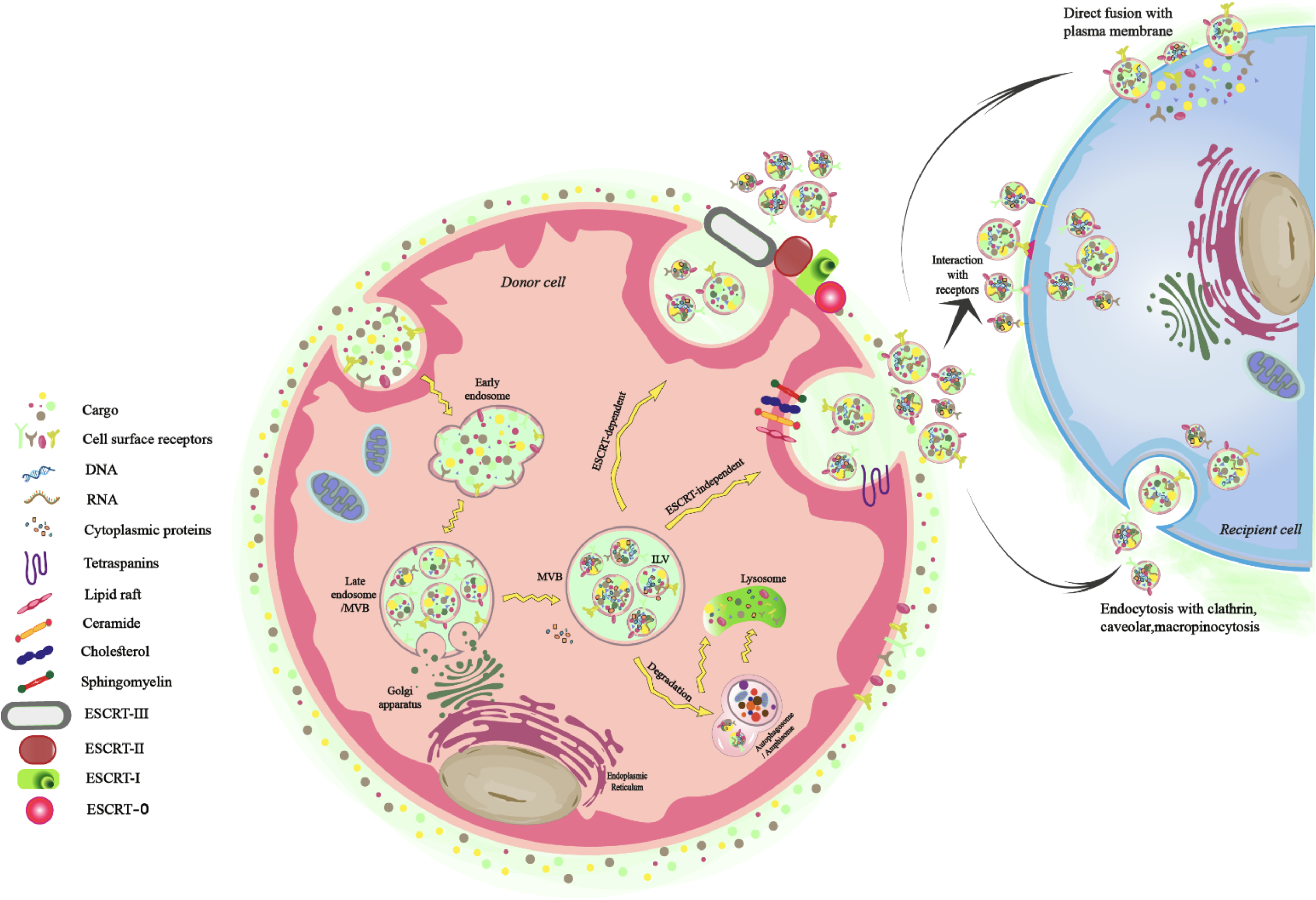
Schematic representation of exosome biogenesis.
ESCRT-dependent pathway
One of the primary and earliest identified mechanisms contributing to exosome biogenesis involves the ESCRT system. This pathway includes a hierarchical series of protein complexes—namely ESCRT-0, ESCRT-I, ESCRT-II, and ESCRT-III—alongside associated regulatory proteins such as ATPase VPS4 and VPS20-associated protein 1 (VTA1).31–33 These complexes localize to the cytoplasmic side of MVB membranes and coordinate the formation of ILVs, cargo recognition, and vesicle scission events.34–36 ESCRT-0, -I, and -II are equipped with ubiquitin-binding domains, enabling them to identify ubiquitinated proteins like the epidermal growth factor receptor and various ligand-receptor assemblies. 37 Key ESCRT-0 components—such as signal-transducing adaptor molecule 1 and hepatocyte growth factor-regulated tyrosine kinase substrate (Hrs)—mediate the selection of tagged cargos in conjunction with phosphatidylinositol-3-phosphate (PI3P), which is enriched in the endosomal membrane.38–40 Hrs interacts with PI3P to facilitate cargo organization and subsequently engages with Tsg101, a component of ESCRT-I. This leads to sequential recruitment of ESCRT-II, composed of subunits such as Vps36, Vps22, and Vps25. 40 ESCRT-I and ESCRT-II jointly promote membrane invagination around ubiquitinated proteins, resulting in ILV formation within MVBs. 41 The process concludes with ESCRT-III filament assembly, which enables membrane scission, detaching the nascent ILVs from the endosomal membrane. 36 ESCRT-III is composed of several core subunits, including charged multivesicular body protein (CHMP)6 (Vps20), CHMP4 (Vps32), CHMP3 (Vps24), and CHMP2 (Vps2), as well as accessory proteins such as Did2, Vps60, and Ist1. 42 The CHMP6 segment of ESCRT-III next associates with ESCRT-II and employs CHMP4, which polymerizes in a helix structure across the neck of the budding ILV sack. Finally, with the incorporation of CHMP3, the bud is finally broken down to create ILVs, and ESCRT-III then disintegrates via ATP hydrolysis that is facilitated by vacuolar protein sorting 4 (Vps4).31,34,37,38 Hence, in the last stage, ATPase VPS4 removes the ubiquitin tags and promotes the disassembly of ESCRT subunits from the MVB membrane. 41 Crucially, the removal of many ESCRT protein subunits or VPS4 can have a substantial effect on the formation of exosomes, resulting in changes in the number, size, and protein makeup of exosomes to different degrees. 43 Also, several investigations have shown that exosomes separated from various cells include several ESCRT-related proteins.34,44
ESCRT-independent pathway
The ESCRT machinery is crucial for exosome formation and intraluminal vesicle generation; however, alternative ESCRT-independent mechanisms have been identified, involving specific proteins like tetraspanins and lipids such as ceramides. 40 Tetraspanins comprise a conserved family of transmembrane proteins, each containing four membrane-spanning domains, that are crucial in facilitating the biogenesis and functionality of exosomes. 45 These proteins are typically enriched in small EVs and interact closely with integrins and other membrane-associated molecules, promoting the assembly of specialized tetraspanin-enriched microdomains (TEMs), which serve as functional platforms for exosome formation.43,46 Studies indicate that tetraspanins are prevalent in endocytic compartments, with members such as CD9, CD63, and CD81 commonly used as molecular markers for exosomes due to their significant vesicular localization.47–49 Beyond serving as markers, these “classical” tetraspanins actively coordinate exosome formation through multiple mechanisms. They assemble into TEMs to induce localized membrane curvature and facilitate vesicle budding. Importantly, they ensure the selective sorting and incorporation of TEM-associated proteins into exosomes, despite lacking intrinsic catalytic activity or receptor-like functions. This process is mediated through their interaction with cytosolic adaptor proteins, such as syntenin and ALG-2-interacting protein X (ALIX), which link tetraspanins to ESCRT-independent pathways, thereby guiding specific cargo such as integrins, major histocompatibility complex (MHC) molecules, and signaling receptors into exosomal membranes.43,47,50–53 Out of the variety of lipids found in considerable quantities in exosomes, each of which has specific roles in exosome synthesis and release and which impact the recipient cells, ceramides are significant in exosome biogenesis. 43 These membrane sphingolipids, known as ceramides promote microdomain-induced endosomal membrane budding through their cone-shaped structure, which allows them to participate in ESCRT-independent membrane deformation.31,54
Secretion pathway
Exosome secretion occurs through three critical stages: targeted transportation of MVBs, the attachment of MVBs to the plasma membrane, and the fusion of the MVB limiting membrane with the plasma membrane. The efficiency of this coordinated process depends on proteins located on the MVB surface, which, after identification, bind to receptors on the target membrane. This mechanism operates much like a conveyor belt, directing MVBs to their intended destinations. 27 Importantly, a number of essential components, such as Rab GTPases and other related proteins, as well as the soluble N-ethylmaleimide-sensitive fusion attachment protein receptor (SNARE) complex—more especially, v-SNARE on the vesicle and t-SNARE on the target membrane—play critical roles in promoting the binding and integration of MVBs into the plasma membrane.40,55 Studies have shown that v-SNAREs and t-SNAREs establish multiple interactions critical to vesicular transport. Additionally, Rabs such as Rab27a, Rab27b, and Rab35 actively interact with SNARE components, including vesicle-associated membrane proteins (VAMPs, v-SNAREs), syntaxins (t-SNAREs), and synaptosome-associated proteins (SNAPs, t-SNAREs). These intricate interactions are vital for precise vesicle targeting, binding, and secretion, thereby ensuring the effective operation of the exosome secretion pathway. 40
Migration and chemotaxis
After being secreted into the extracellular space, exosomes do not diffuse randomly; rather, they exhibit directed migration and chemotaxis, which regulate their biodistribution and tissue tropism. This mechanism is primarily governed by molecular signatures expressed on their membranes, notably integrins, tetraspanins, and adhesion molecules, which interact with ECM components and chemokine gradients to enable targeted trafficking.56–59 These surface proteins function as molecular “addresses,” binding to extracellular matrix ligands and cell surface receptors, enabling exosomes to follow chemotactic gradients in the extracellular environment. 56 Moreover, spatially polarized exosome secretion mediated by Rab GTPases (e.g., Rab27a, Rab35) directs vesicle release to certain cellular locales, hence enhancing directed movement. 60 Beyond surface interactions, exosomal cargo such as cytokines, growth factors, and matrix-remodeling enzymes can modify the extracellular environment, amplifying recruitment signals for subsequent exosomes and recipient cells. 61
Internalization pathway
For exosomes to carry out their function as messengers in intercellular communication processes, they must first enter the extracellular space, where they must contact receptor cells and cause them to undergo modifications. There are now three known mechanisms for cells to communicate with one another through exosomes: (1) endocytosis, phagocytosis, or micropinocytosis; (2) interaction between receptors and ligands; and (3) direct membrane fusion. The first method entails the internalization of exosomes through processes such as endocytosis, phagocytosis, and macropinocytosis. During this process, the target cell engulfs the exosome’s membrane and contents, encapsulating them within a newly formed vesicle. 62 According to experimental evidence, the main way EVs enter cells is by endocytosis. This process is typically a fast and temperature-dependent mechanism, which is reduced by low temperatures.25,62
Endocytosis can occur through the mediation of clathrin, lipid rafts, heparin sulfate proteoglycans, and caveolin-dependent mechanisms. Clathrin-mediated endocytosis is a conventional pathway for exosome uptake, relying on the assembly of transmembrane receptors and ligands. This approach utilizes a clathrin triskelion scaffold to form clathrin-coated vesicles, which subsequently undergo uncoating and merge with endosomes.25,62 Furthermore, this tightly controlled process may be impacted by the makeup of the exosome and cargo. 25 Exosomes derived from phagocytic cells are primarily internalized by immune cells such as dendritic cells and macrophages, which depend on the actin cytoskeleton for vesicle trafficking. These cells utilize macropinocytosis, a nonspecific uptake mechanism that engulfs extracellular fluids, nutrients, and antigens into large vesicular structures called macropinosomes. As they mature, macropinosomes can either fuse with lysosomes for degradation or interact with the plasma membrane for recycling purposes. This uptake pathway is modulated by various molecular cues, including growth factors, cholesterol levels, the activity of Rac1 GTPase, Na+/H+ exchange, and, in certain contexts, dynamin function.25,62 Beyond uptake mechanisms, exosomes also engage in receptor-mediated signaling, wherein ligands or membrane proteins present on exosomes bind to specific receptors on recipient cells. This interaction initiates intracellular cascades that contribute to intercellular communication and modulation of cellular behavior. Finally, another often-seen process is that an acidic pH helps the destination cell’s plasma membrane merge with the donor cell’s derived exosome. This might be because the overall ionic charge on the surface of the exosome changes when it is released or because the lipid content changes. 62
Exosome Components
Numerous types of cargo bioactive molecules are found in exosomes. 63 The composition and abundance of these molecules vary depending on the origin and physiological state of the parent cell and can be influenced by external factors such as stress, hypoxia, specific treatments, and environmental stimuli.64,65 recipient cells, thereby initiating targeted signaling cascades. Internally, they encapsulate a broad spectrum of bioactive molecules—including nucleic acids, proteins, and lipids—that can impact the recipient cell’s genomic, proteomic, or metabolic pathways upon delivery. This combination of surface-mediated targeting and internal cargo delivery underscores the dual functional capacity of exosomes in mediating complex intercellular communication. As such, they serve as pivotal agents among EVs, playing a crucial role in the coordination and regulation of cellular signaling networks. 43 Figure 2 provides a schematic overview of the exosomal membrane surface molecules and internal cargo components, highlighting key elements involved in biological functions and intercellular communication.
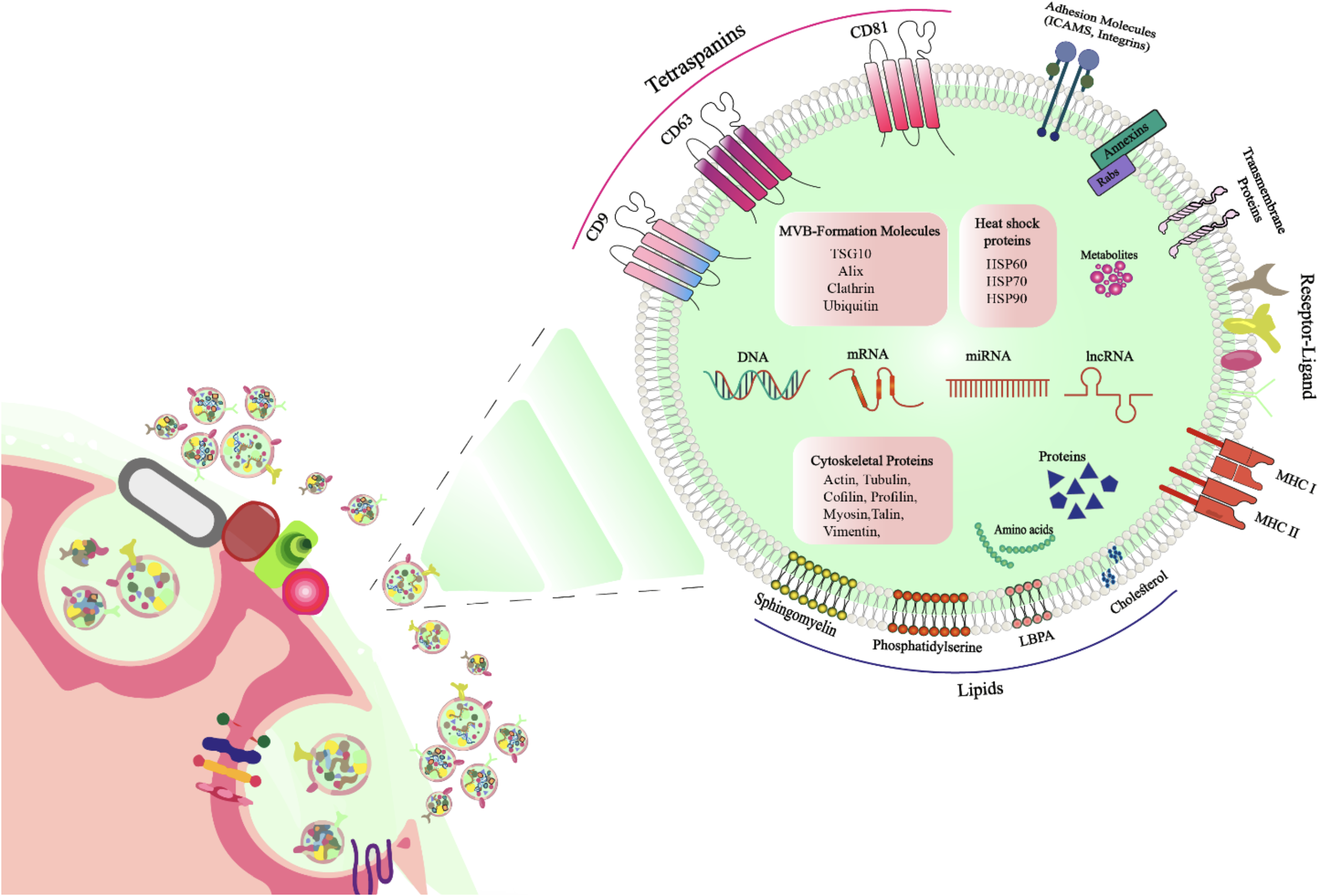
Schematic representation of exosome structure, showing surface-associated molecules and internal cargo.
Exosomal mRNAs in Osteogenesis and Bone Regeneration
Messenger RNA is initially transcribed in the nucleus and then transported to the cytoplasm for translation by ribosomes. Mature mRNA in eukaryotes consists of five key structural components: a 5′-methylated guanosine cap (m7Gp3N), 5′ and 3′ untranslated regions (UTRs), an open reading frame (ORF), and a poly(A) tail with 50–250 adenosine residues. The ORF encodes the protein sequence, while other elements are vital for regulating mRNA translocation, translation, and stability. 66 A methyl guanosine cap, or m7Gp3N structure, is a posttranscriptional modification found at the 5′-end of mature mRNA in eukaryotes. This modification is crucial for initiating mRNA translation.67–69 This signal aids ribosome recognition of mRNA, enhances ribosome binding, and initiates translation at AUG. The cap structure improves mRNA stability and protects it from 5′ → 3′ exonuclease degradation. 69 Since the first discovery of mRNA in the 1960s and the synthesis of biologically active mRNA in 1984,70,71 research has uncovered the presence of functional mRNAs in exosomes derived from both mouse and human mast cells. These mRNAs and microRNAs, collectively termed exosome shuttle RNAs, can maintain their biological activity in recipient cells.72,73
Exosomal mRNAs, among the largest known transcripts, can be horizontally transferred and subsequently translated into functional proteins within recipient cells.74,75
Significantly, mRNAs from mouse mast cells have demonstrated the ability to infiltrate human mast cell lines, highlighting the potential of exosomes as efficient vehicles for intercellular mRNA delivery. Analysis of MC/9-derived exosomes revealed that 270 of 1,300 mRNAs were missing in donor cells, indicating that RNA incorporation into exosomes is a highly selective process.72,76–78 While the mechanisms of selective RNA sorting remain not entirely clear, data from microarray and next-generation sequencing investigations suggest that this process is governed by various molecular pathways. This encompasses the identification of particular RNA sequence motifs, interactions with RNA-binding proteins including Heterogeneous Nuclear Ribonucleoprotein A2/B1 (hnRNPA2B1), Y-Box Binding Protein 1 (YBX1), and Synaptotagmin Binding Cytoplasmic RNA Interacting Protein (SYNCRIP), as well as the involvement of ESCRT.43,79–82 These coordinated processes ensure that exosomal RNA profiles are distinct from their parent cells and can vary according to cell type and species. Importantly, the translation of exosomal mRNAs in recipient cells confirms that their functional integrity is maintained during transfer. Collectively, these findings highlight that exosomal mRNAs are selectively packaged and transferred in a functional form, ensuring their role as active mediators of intercellular communication. 65
To date, over 1600 unique mRNAs have been identified in mammalian exosomes, many of which contribute to the reprogramming of target cell protein expression.10,77,83,84 The implications for bone regeneration are particularly significant. Exosomal mRNAs can influence the behavior of osteoblasts and other bone-forming cells. Studies identified seven mRNAs (RPS2, DGKA, ACIN1, DKK2, Xsox17, DDX6, and Lsm2) in exosomes from differentiated bone marrow mesenchymal stem cells (BMSCs) that were implicated in osteogenic differentiation and mineralization.85,86 Moreover, exosomes can deliver specific mRNA molecules that encode growth factors, transcription factors, or other regulatory proteins essential for bone formation and healing. A study by Yang et al. showed that inhibiting the expression of the target mRNA (Bmp2 mRNA) in the parent cell results in the accumulation of exosomes containing high levels of desired mRNA, and then these exosomes may be transferred to the target cell to make therapeutic proteins for disorder treatment. The approach of this study was one of the new strategies for bone regeneration therapy, namely, RNA therapy techniques with exosomes as carriers that involved the construction of bone morphogenetic protein-2 (BMP2) and NoBody plasmids, resulting in the production of modified exosomes enriched in Bmp2 mRNA. This carrier exosome of mRNA had such a tremendous impact on the process of bone repair that the binding of engineered exosomes to the hydrogel scaffold (GelMA) with modified CP05 had been shown to significantly enhance the sustained release of exosomes and facilitate the process of osteogenesis in critical bone defects. 69 Similarly, Ma et al. used parental cells transfected with vascular endothelial growth factor (VEGF)-A and BMP2 plasmids to generate therapeutic exosomes (t-sEVs) loaded with osteogenic mRNAs. These were delivered via PEGS-A hydrogels in a critical-sized bone defect model, showing simultaneous induction of osteogenesis and angiogenesis. 87 In another notable study, Guo et al. explored exosomes from stem cells of human exfoliated deciduous teeth (SHED), which contained mitochondrial transcription factor A (TFAM) mRNA. SHED-derived exosomes successfully transferred TFAM mRNA into dental pulp stem cells, promoting glutamate metabolism, mitochondrial oxidative phosphorylation, and osteogenic differentiation in both cranial and mandibular bone defect models. 88 The immunomodulatory functions of exosomal mRNAs are also relevant. Chen et al. reported that M2-like macrophage-derived exosomes were enriched in IL-10 mRNA, which, upon transfer to BMSCs and bone marrow-derived macrophages, upregulated IL-10 expression and activated the IL-10/IL-10R pathway, promoting anti-inflammatory responses and enhancing bone metabolism. 89 Understanding the composition and regulatory roles of these mRNA molecules can provide insights into designing more effective therapeutic approaches for bone injuries and diseases. Recent findings suggest that manipulating the efficacy of exosomal mRNA could potentially lead to breakthroughs in regenerative medicine by optimizing the regenerative environment and accelerating bone repair. Among these manipulations is the direct modification of exosomes through electroporation and sonication to enhance their functionality. 90 Gene editing is another promising method for improving the functionality of exosomes. A notable example is the study by Li et al., which implemented a stem cell-based gene therapy technique. In this approach, MSCs were genetically engineered to carry the BMP2 gene, resulting in the production of exosomes (MSC-BMP2-Exo) with improved capabilities for bone regeneration. Finally, the overall findings of this study indicated that MSC-BMP2-Exo demonstrated excellent biocompatibility and significantly accelerated bone healing, highlighting its strong potential for clinical applications. 91 Table 1 summarizes the key exosomal mRNAs associated with osteogenesis and bone regeneration, including their cellular origin, molecular function, model of application, and therapeutic impact.
The Exosomal mRNA Cargos Involved in Osteogenesis and Bone Regeneration

MSC, mesenchymal stem cells.
Exosomal miRNAs in Osteogenesis and Bone Regeneration
Mechanistic roles of miRNAs in osteogenesis
MicroRNAs are small, highly conserved noncoding RNAs with a typical length of 18–26 nucleotides. Their biosynthesis begins with the transcription of primary miRNAs (pri-miRNAs) by RNA polymerase II, which are subsequently processed by Drosha into ∼60–100 nucleotide precursor miRNAs (pre-miRNAs) in the nucleus. These pre-miRNAs are exported to the cytoplasm via Exportin-5/Ran-GTP, where Dicer cleaves them into ∼22 nucleotide RNA duplexes, later matured into single-stranded miRNAs.92–94 miRNAs function as posttranscriptional regulators of gene expression and are involved in various biological processes, including stem cell self-renewal, differentiation, growth, apoptosis, immune responses, tumor progression, and metabolic regulation.76,95–97 Additionally, they function as negative regulators of gene expression after transcription and negative mediators of mRNA translation efficiency.97,98 miRNAs attach to the 3′-UTR of target mRNAs by binding to the seed sequence, which consists of the first 2–7 nucleotides in the miRNA 5 region. This binding, in turn, ultimately causes instability, degradation of the mRNA, and suppression of the expression of target genes.76,98,99 Likewise, given that a single miRNA can target multiple mRNAs—and conversely, several miRNAs may target the same mRNA—their regulatory influence is extensive and multifaceted. 100 Both cellular and exosomal miRNAs are implicated in key biological functions, including cell cycle progression, immune modulation, apoptosis, cancer progression, and metabolic control.76,98 To date, over 700 distinct exosomal miRNAs have been identified across various cell types, and their presence in circulation allows them to influence gene expression in local and distant recipient cells. 84
In the context of osteogenesis, exosomal miRNAs play vital roles by modulating pathways central to bone formation and remodeling. These miRNAs influence the behavior of osteoblasts, osteoclasts, and MSCs by facilitating intercellular signaling. 101 For instance, exosomal miR-26a-5p derived from M2 macrophages enhances osteogenic differentiation of bone marrow MSCs (BM-MSCs) by suppressing adipogenesis and upregulating osteogenic markers such as alkaline phosphatase (ALP), runt-related transcription factor 2 (RUNX2), osteopontin (OPN), and collagen type II (COL2). 102 Similarly, exosomal miR-101 has been shown to regulate osteogenic differentiation through modulation of the FBXW7/HIF1α/FOXP3 axis. FBXW7, an E3 ubiquitin ligase, promotes degradation of HIF1α, a transcription factor involved in osteogenesis. Yanhong Li and colleagues demonstrated that miR-101 inhibits FBXW7, thus stabilizing HIF1α and promoting osteogenesis in MSCs. 103 Further evidence by Jiang et al. identified miR-25, encapsulated in BMSC-derived exosomes, as a positive regulator of bone regeneration. Through inhibition of SMURF1—a factor that degrades RUNX2—miR-25 preserves RUNX2 levels, thus enhancing osteogenesis. 104 In a similar vein, osteogenic differentiation in human mesenchymal stem cells (hMSCs) is promoted by miRNAs such as miR-146a-5p, miR-503-5p, miR-483-3p, and miR-129-5p, and inhibited by miR-32-5p, miR-133a-3p, and miR-204-5p. These miRNAs modulate critical signaling pathways including the phosphatidylinositol 3-kinase (PI3K)/protein kinase B (AKT) and mitogen-activated protein kinases (MAPK) to orchestrate osteogenic outcomes. 105 The inflammatory microenvironment is also intricately linked to osteogenesis. Exosomal miRNAs secreted by immune cells such as macrophages have been shown to influence bone regeneration. Liu et al. reported that exosomes derived from M1 macrophages enriched in miR-21a-5p significantly enhanced osteogenic differentiation in vitro, especially during early phases of inflammation. These findings support the dual pro-osteogenic capacity of exosomes from both M1 and M2 macrophage subsets. 106 To further elucidate the impact of exosomal miRNAs from diverse cellular origins, Yang et al. isolated osteoclast-derived exosomes and identified miR-23a-5p as a negative regulator of osteogenesis. 107 This miRNA targets and suppresses RUNX2, thereby inhibiting osteoblast function and reducing osteogenic potential. Collectively, these studies highlight the mechanistic complexity of miRNAs in osteogenesis, emphasizing the dynamic interplay between exosomal cargo and cellular signaling networks in bone formation.
Exosomal miRNAs in disease models
Osteoarthritis and bone fracture
Exosomal miRNAs are increasingly recognized as crucial mediators in osteoarthritis and bone fracture repair. BMSC-derived exosomal miR-206 promotes osteoblast proliferation and differentiation by targeting the 3′-UTR of E74-like factor 3, reducing its expression and consequently decreasing osteoblast apoptosis in osteoarthritic conditions. 108 The role of exosomes in intercellular signaling is further exemplified in fracture healing. Research by Si Chen et al. demonstrated that exosomes from human adipose-derived stem cells overexpressing miR-375 significantly enhanced osteogenic differentiation by targeting IGFBP3, a negative regulator of osteogenesis. When combined with a hydrogel, these miR-375-enriched exosomes facilitated sustained release and markedly improved bone repair in a rat calvarial defect model, highlighting a novel therapeutic strategy for bone regeneration. 109 Additionally, miR-25 delivered via BMSC-derived exosomes was shown to promote bone healing by downregulating SMURF1, thus preserving Runx2 and enhancing osteogenesis 104 (Fig. 3).
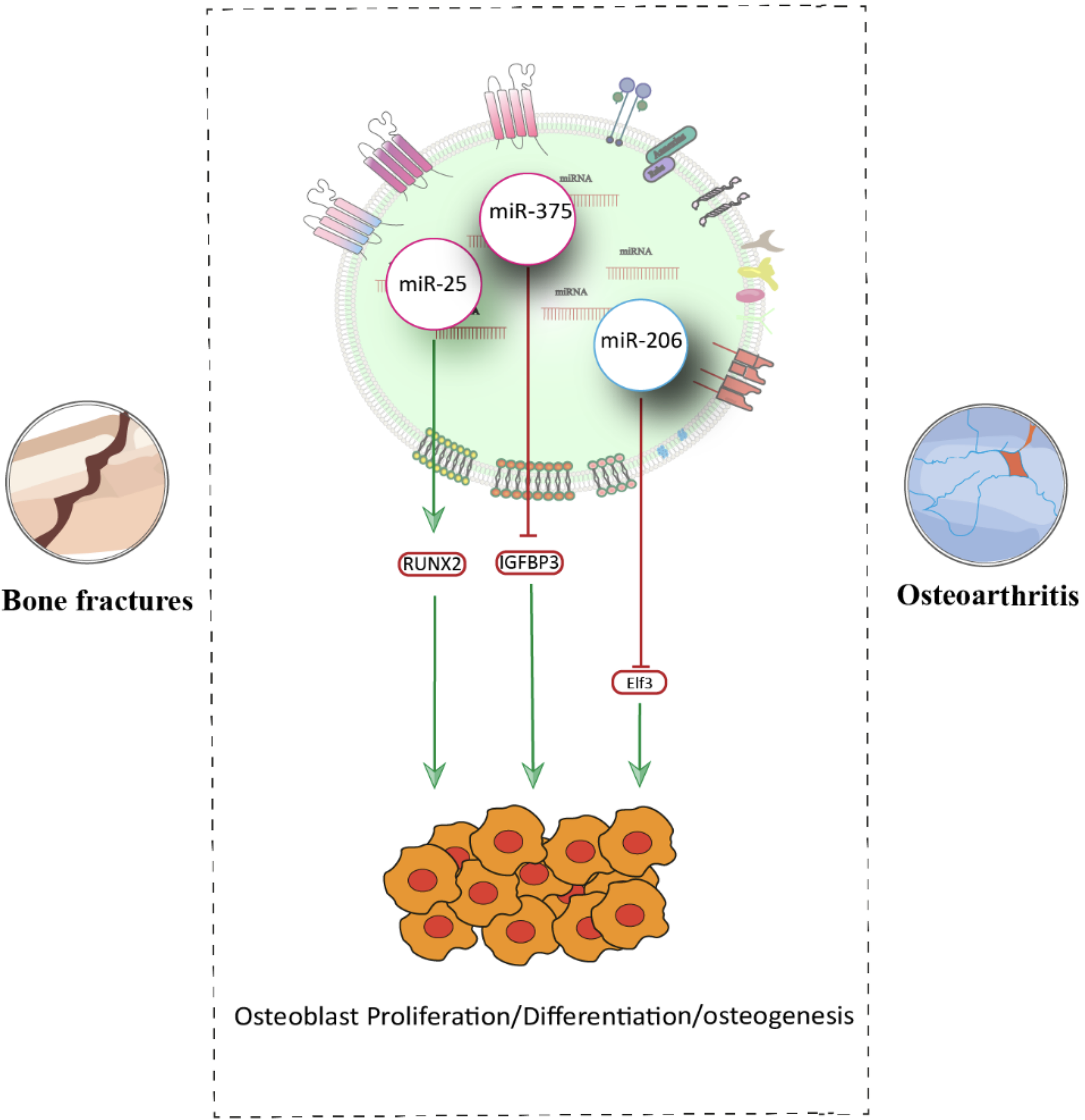
Schematic representation of exosomal miRNAs in osteoarthritis and bone fracture models.
Diabetes
Diabetes mellitus (DM) is a chronic metabolic disorder associated with impaired bone healing and increased fracture risk.100,110 Wang et al. reported that exosomes derived from diabetic BMSCs (DM-Exos) exhibit reduced osteogenic potential compared with those from nondiabetic controls (N-Exos), due to decreased levels of miR-140-3p. Restoration of miR-140-3p or application of N-Exos enhanced osteogenesis by suppressing the plexin B1 (Plxnb1)/Sema4D/RhoA/Rho-associated coiled-coil kinase pathway. 100 Diabetes has been linked to changes in bone metabolism, according to earlier research, although the exact mechanism is yet unknown, and these changes in bone metabolism may manifest with changes in bone mass. For instance, Han et al. study revealed that diabetic BMSC-derived exosomes promote adipogenesis and inhibit osteogenesis, primarily through the action of miR-221, thus contributing to bone–fat imbalance in diabetic conditions. 110 Furthermore, Zhang et al. showed that exosomes from diabetic macrophages carry elevated levels of miR-144-5p, which downregulates Smad1 and impairs osteogenic differentiation of BMSCs. 111
Osteoporosis
OP, characterized by decreased bone density and increased fragility, is influenced by multiple exosomal miRNAs. Hairong Su found that exosomal miR-382, derived from Human Bone Marrow Mesenchymal Stem Cell (hBMSCs), is downregulated in OP patients, whereas its target gene SLIT2 is upregulated. Exosomal miR-382 enhances osteogenesis by binding to the 3′-UTR of SLIT2, promoting its degradation and facilitating osteoblast differentiation. 112 Similarly, Murong You demonstrated that exosomal miR-21-5p from BMSCs enhances proliferation, ALP activity, and osteogenic differentiation in human fetal osteoblast cells by targeting Krüppel‐like factor 3 (KLF3). Upregulation of miR-21-5p in exosomes resulted in improved bone mass and osteogenesis in OP models. 113 Furthermore, Jiang et al. confirmed the presence of significant amounts of miR-21 in exosomes derived from MSCs extracted from patients with OP compared with healthy individuals. Overexpressing miR-21 suppressed osteogenesis by binding to the 3′-UTR of SMAD7. SMAD7 is a crucial transcription factor in the downstream pathways of bone morphogenetic proteins, regulating the osteogenic differentiation of osteoblasts generated by these proteins. 114 Additionally, miR-935 has been identified as a positive regulator of osteogenesis. BMSC-derived exosomes transfer miR-935 to osteoblasts, where it downregulates STAT1, which is an important gene involved in OP, and promotes bone formation, alleviating OP symptoms in vivo. 115 In contrast, miR-424-5p inhibits osteoblast differentiation by targeting WIF1, leading to suppression of Wnt/β-catenin signaling and decreased expression of osteogenic markers such as RUNX2, OCN, and OPN. 116 Postmenopausal osteoporosis (PMO) is the most common form of primary OP in elderly women and is driven by estrogen deficiency. Li’s study showed that miR-186 is upregulated in PMO mice treated with BMSC-derived exosomes. This miRNA promotes osteoblast proliferation and differentiation via the Hippo signaling pathway, thus counteracting PMO progression. 117 Li et al. also reported elevated levels of miR-214-3p in osteoclast-derived exosomes from OVX rats and elderly women with fractures. This miRNA inhibits osteogenesis by binding to the 3′-UTR of ATF4 mRNA, a key transcription factor in osteoblast function 118 (Fig. 4).
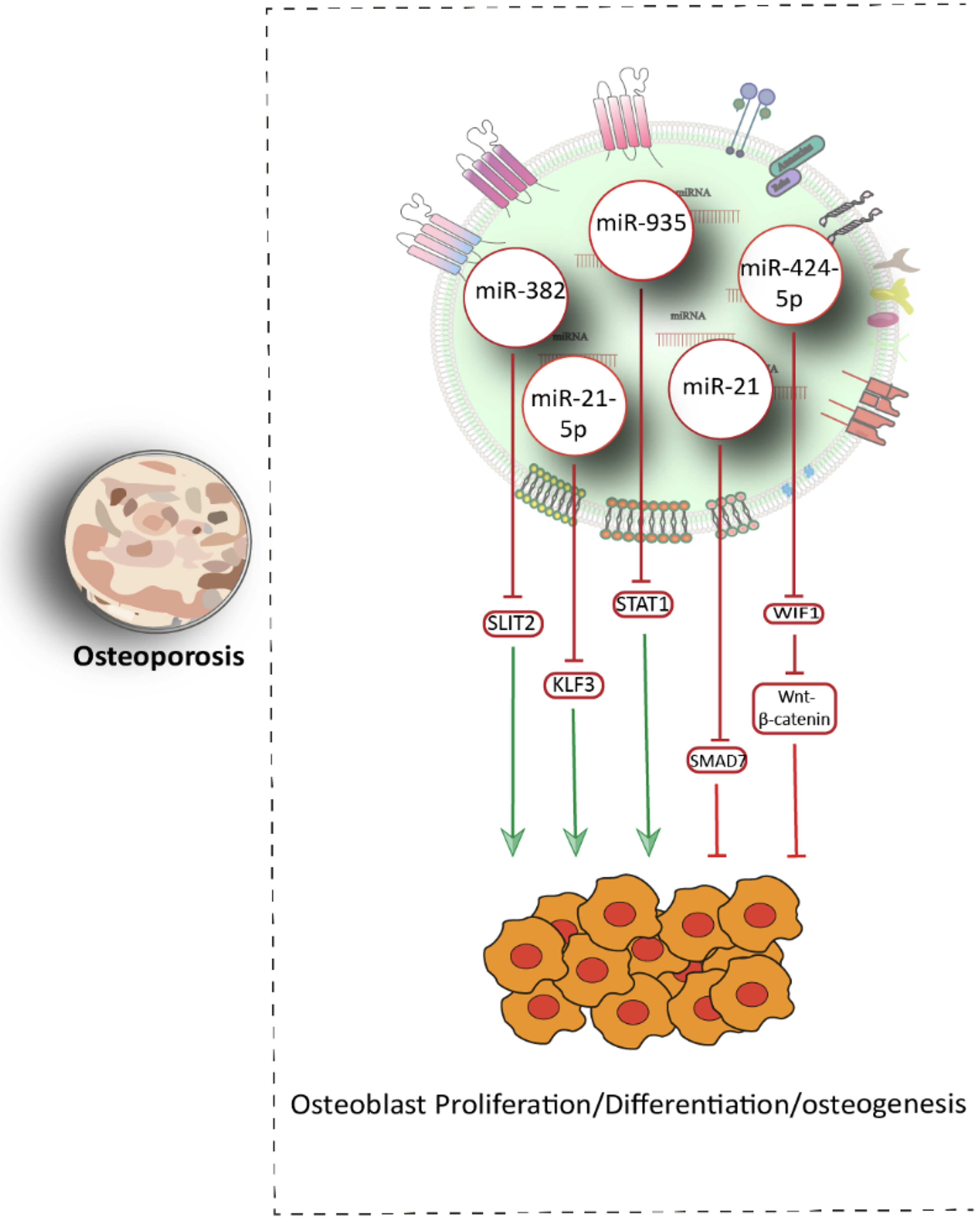
Schematic representation of exosomal miRNAs in osteoporosis model.
Aging
The aging process is associated with bone resorption and protracted fracture repair, and age-related OP is linked to markedly diminished bone production due to reduced quantity and impaired osteogenic capacity of MSCs.119,120 According to this issue, some research aims to investigate the protection of function and efficiency of exosomes on osteogenic differentiation and fracture healing in MSCs with age. Tao Xu et al. successfully isolated and identified MSCs from young and aged rats, and subsequently obtained exosomes from them. miRNA array analysis demonstrated that miR-128-3p was significantly upregulated in aged-exos, and it was also confirmed that the expression levels of miR-128-3p in MSCs and their secreted exosome increased as cell senescence manifested. So far, Smad5, Smad9, and BMP3 have been identified as downstream target genes of exosomal miR-128-3p, in this experiment, it was verified that Smad5 was a direct downstream target of miR-128-3p that was inhibited by overexpressed miR-128-3 in Aged-Exos. 95 In addition to the effect of the aging of stem cells on reducing the effectiveness of exosomes in osteogenesis and fracture repair, the aging of other cells may have similar results. Chen Yao investigated the impact of senescent osteocytes on bone homeostasis during the progression of age-related OP and the underlying mechanism. Exosomes from tert-butyl hydroperoxide (TBHP)-induced senescent MLO-Y4 cells exhibited reduced levels of miR-494-3p, leading to upregulation of PTEN in osteoblasts. This resulted in suppression of the PI3K/AKT pathway and impaired osteogenic differentiation, thus linking osteocyte senescence to age-related OP. 121
Exosomal miRNAs and angiogenesis–osteogenesis crosstalk
Angiogenesis, the formation of new blood vessels, is integral to the regenerative processes of bone fracture repair, including inflammation, stem cell differentiation, osteogenesis, and chondrogenesis, and plays a crucial role in bone osteogenesis process and formation.122,123 To achieve the desirable level of bone regeneration, particularly in significant bone defects, dual-functional regulation of angiogenesis and osteogenesis is essential. While exosomes have been shown to promote bone regeneration by enhancing osteogenesis and angiogenesis, they have also been reported to further enhance their proangiogenic ability through functional stimulation of mesenchymal stromal cells.124,125 Liu et al. found that BMSC-derived exosomes under strontium-substituted calcium silicate stimulation enhanced HUVEC angiogenesis by elevating proangiogenic miR-146a cargos and inhibiting Smad4 and NF2, positioning Sr-CS-Exo as a dual-action agent in vascularized bone repair. 124 In another study. Pan et al. developed multifunctional hydrogel microparticles loaded with exosomal miR-29a, which promoted osteogenesis and angiogenesis by suppressing HDAC4 and increasing expression of RUNX2 and VEGF, demonstrating the therapeutic potential of hydrogel-based delivery systems. 126 Wu et al. investigated the stimulating effect of magnetic nanoparticles on bone MSCs and their derived exosomes with the aim of bone regeneration as well as proangiogenic activities. upregulated miR-1260a in exosomes derived from BMSCs preconditioned with a low dose of Fe3O4 nanoparticles, and the static magnetic field (SMF) called BMSC-Fe3O4-SMF-Exos enhanced osteogenesis and angiogenesis by suppressing HDAC7 and COL4A2 expression 125 (Fig. 5).
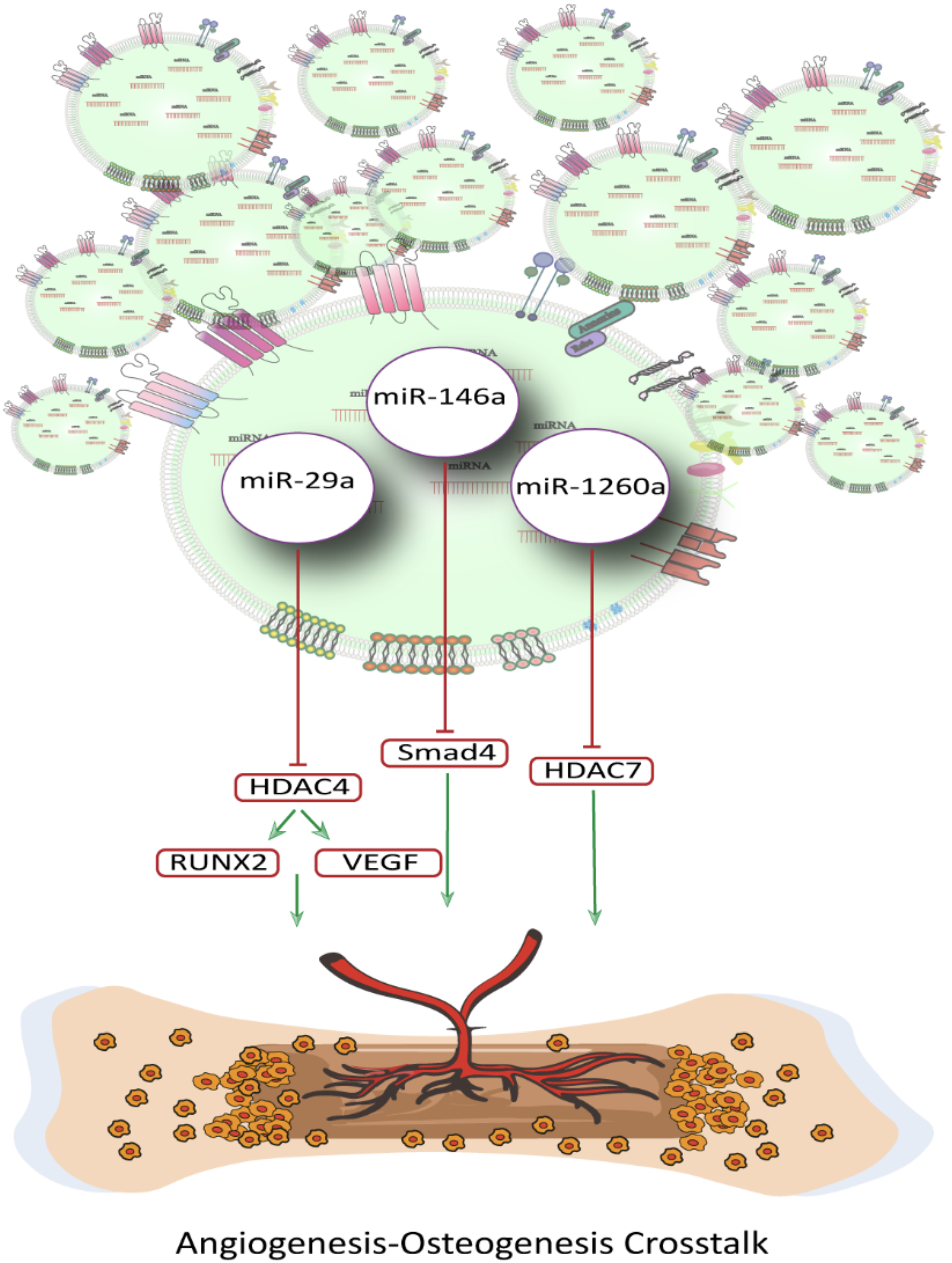
Schematic representation of exosomal miRNAs and angiogenesis–osteogenesis crosstalk.
Exosomal miRNAs and immune–osteogenesis crosstalk
Bone regeneration involves intricate communication not only among resident osteogenic cells but also with immune cells that share developmental origins and overlapping signaling networks. In the bone microenvironment, immune cells actively sense cues from osteoblasts and osteoclasts, releasing cytokines and regulatory factors that can either promote or suppress osteogenic processes. Emerging evidence highlights that exosomal microRNAs secreted by macrophages, dendritic cells, and T cells serve as critical mediators of this intercellular crosstalk, influencing stem cell differentiation and bone repair.
Macrophage-derived exosomal miRNAs play a central role in regulating osteogenesis. Luo et al. demonstrated that exosomes from M1 macrophages are enriched in miR-21a-5p, which enhances osteoblast differentiation by directly targeting GATA2 and activating downstream osteogenic pathways. 127 Similarly, Liu et al. showed that miR-21a-5p-enriched M1 macrophage exosomes can be internalized by bone marrow-derived MSCs, promoting early osteoblastic differentiation while concurrently inhibiting osteoclastogenesis through targeting key regulators such as SKP2. 106 These findings highlight the multifaceted role of miR-21a-5p in macrophage–BMSC crosstalk and its therapeutic potential for cell-free bone regeneration strategies, particularly under inflammatory conditions. Exosomes from M2 macrophages also exhibit osteoinductive properties. Li et al. reported that miR-690-enriched M2 macrophage exosomes promote osteogenic differentiation and suppress adipogenesis in BMSCs via the miR-690/IRS-1/TAZ axis, suggesting potential applications in bone loss disorders. 128 Insights from macrophage-derived exosomal miRNAs have driven studies examining the effects of immunomodulatory peptides on osteogenic signaling. The immunomodulatory peptide DP7-C has been shown to enhance osteogenesis by increasing miR-21b levels in macrophage-derived exosomes, targeting SOCS1, and activating the JAK2/STAT3 pathway, providing a promising cell-free strategy for bone regeneration in periodontitis. 129 Another study demonstrated that DP7-C-mediated delivery of miR-26a into BMSCs, subsequently loaded into secreted exosomes, promoted proliferation, migration, and osteogenic differentiation, mitigating bone loss in a periodontitis model. 130 Biomaterial-induced exosomal miRNAs further illustrate the crosstalk between immune modulation and osteogenesis. Macrophages treated with biphasic calcium phosphate ceramics featuring submicron surface architecture secreted exosomes with elevated miR-142a-5p, promoting MSC osteogenic differentiation via PTEN/AKT signaling. 131 Similarly, titanium nanotube (TNA-40) stimulation induced exosomal miR-3473e production, enhancing osteogenesis in BMSCs and angiogenesis in endothelial cells through Akt1 activation, underscoring a key osteoimmunomodulatory axis for improved bone implant integration. 132
Dendritic cell-derived exosomes also contribute to bone regeneration. For example, mature dendritic cell exosomes carrying miR-335 promote BM-MSC proliferation and osteogenic differentiation by suppressing Hippo signaling through LATS1 targeting. 133 In this context, Hou et al. reported that M2 macrophage-derived exosomes enriched with miR-365-2-5p enhance osteogenic differentiation in progenitor cells via direct targeting of OLFML1 and activation of downstream osteogenic pathways. 134
Regulatory T cell-derived exosomes represent a promising therapeutic avenue. Chen et al. demonstrated that Treg exosomes transfer miR-142-3p to BMSCs and human umbilical vein endothelial cells, simultaneously promoting osteogenesis and angiogenesis through inhibition of TGFBR1/SMAD2 signaling, ultimately accelerating fracture healing in vivo. These findings highlight the potential of Treg-derived exosomes and their miRNA cargo as innovative, cell-free therapies for bone regeneration. 135 A summary of key exosomal miRNAs, their cellular sources, molecular targets, associated disease models, and therapeutic impacts is presented in Table 2.
The Exosomal miRNAs Cargos Involved in Osteogenesis and Bone Regeneration
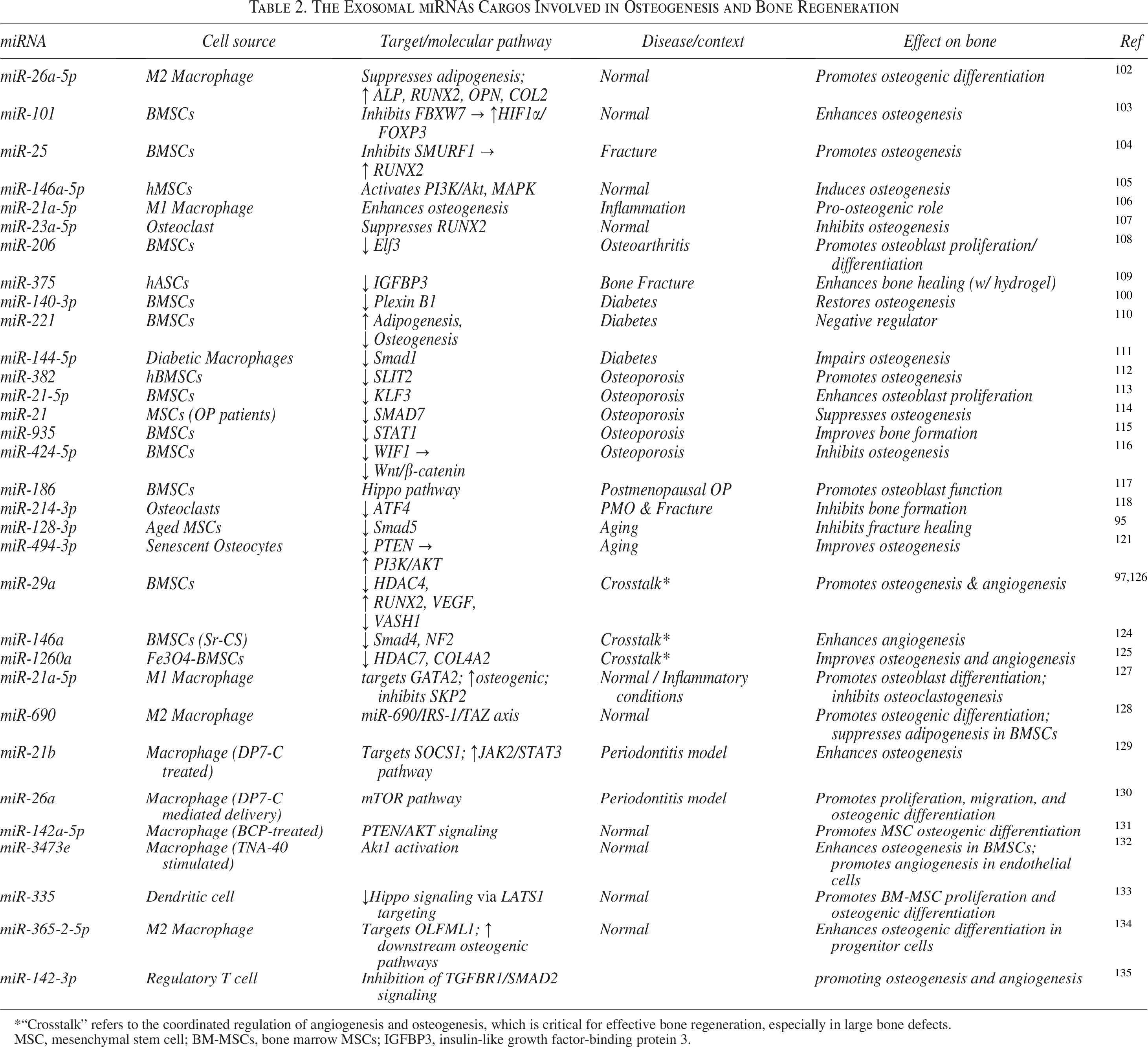
*“Crosstalk” refers to the coordinated regulation of angiogenesis and osteogenesis, which is critical for effective bone regeneration, especially in large bone defects.
MSC, mesenchymal stem cell; BM-MSCs, bone marrow MSCs; IGFBP3, insulin-like growth factor-binding protein 3.
Therapeutic Potential of Exosomal miRNAs
Exosomal miRNAs are increasingly recognized not only as biomarkers of disease but also as active therapeutic agents in regenerative medicine. Growing evidence indicates that the transfer of miRNAs plays a pivotal role in enhancing the therapeutic potential of MSC-derived exosomes across diverse disease models. In parallel with these biological strategies and despite their high potential in therapeutic delivery, exosomes still face several limitations in clinical translation, including low yield, inefficient targeting, and suboptimal therapeutic efficacy. 136 To overcome these challenges, a number of exosome engineering approaches have been developed to optimize the therapeutic performance of exosomal miRNAs. These strategies mainly include cargo-loading approaches (endogenous and exogenous), surface modification, EV–material hybridization/delivery platforms.15,137 This section focuses on the therapeutic effects of engineered exosomes in bone regeneration and provides a foundation for their prospective clinical applications.
Endogenous cargo loading refers to the genetic modification of parental cells using transfection or transduction systems such as lentiviral vectors, plasmids, or liposome-based carriers. 136 Through this strategy, genetically engineered cells secrete exosomes that are endogenously enriched with specific miRNAs and maintain stable therapeutic profiles.136,137 For instance, Jo et al. genetically modified human adipose-derived MSCs via lentiviral transduction to stably overexpress miR-375. The resulting exosomes, enriched in miR-375, significantly promoted the repair of calvarial bone defects and facilitated bone regeneration. 109 In related studies, bone marrow-derived MSCs were transduced with lentiviral vectors to stably overexpress miR-140-3p, and the resulting exosomes promoted bone regeneration in diabetic rats by targeting the Plxnb1 pathway. 100 In another study, BMSCs were temporarily transfected with miRNA mimics to enhance exosomes carrying therapeutic cargo for miR-150-5p. The modified exosomes were later incorporated onto magnetic nanoparticles (GMNPE-EVs) to improve bone targeting. Lentiviral vectors carrying miR-150-5p mimics or inhibitors were delivered in vivo to regulate downstream signaling pathways, thereby demonstrating the osteoprotective function of miR-150-5p in diabetic OP. 138
Another widely used endogenous strategy is the transfection of parental cells with miRNA mimics or inhibitors using liposome-based carriers such as Lipofectamine, which enables the enrichment of secreted exosomes with specific functional cargos. For instance, exosomes derived from BMSCs transfected with a miR-29a mimic exhibited dual therapeutic activity by simultaneously promoting osteogenesis and angiogenesis through modulation of the Vascular Parthanatos Signaling Protein 1 (VASH1) pathways. 97 In another investigation, exo-MSCs loaded with antagomirs against miR-128-3p via Lipofectamine were shown to restore Smad5 expression, thereby improving fracture healing in aged models. 95 Likewise, mimics of miR-140-3p and miR-382 introduced into BMSCs generated exosomes capable of correcting osteogenic deficiencies under diabetic and osteoporotic conditions by targeting the Plxnb1 and SLIT2 pathways, respectively.100,112 More recently, Wu et al. transfected hBM-MSCs with an miR-181b mimic, leading to the secretion of exosomes enriched in miR-181b. These modified exosomes augmented VEGF and BMP2 expression, facilitated M2 macrophage polarization, mitigated inflammation via the PRKCD/AKT signaling pathway, and ultimately accelerated osteogenesis and osseointegration both in vitro and in vivo. 139 Based on these findings, a subsequent study employed multifunctional injectable hydrogel microparticles as a delivery platform for BMSC-derived exosomes enriched in miR-29a. In addition to facilitating the sustained release of therapeutic exosomes, this engineered system also significantly improved bone regeneration by coordinating the promotion of osteogenesis and angiogenesis. Mechanistically, the proregenerative effects were mediated, at least in part, through the regulation of key osteogenic pathways involving RUNX2 and HDAC4. 126
Although therapeutic miRNAs and proteins are most commonly incorporated into exosomes via endogenous loading strategies, exogenous loading refers to the direct encapsulation of small-molecule cargos into preisolated exosomes. A variety of physical approaches, including saponin-assisted permeabilization, repeated freeze–thaw cycles, electroporation, extrusion, and sonication, have been widely employed to enhance exosomal membrane permeability and facilitate the efficient incorporation of therapeutic payloads. These engineered exosomes have been applied across diverse disease models, highlighting the versatility of exogenous loading as a complementary strategy to augment the therapeutic utility of exosome-based delivery systems.136,140–142
Beyond cargo engineering, other strategies such as exosome surface functionalization to enhance tissue targeting have advanced exosome-based bone therapies. For instance, Wang et al. engineered MSC-derived EVs with alendronate (ALN) to improve binding to hydroxyapatite, facilitate EV targeting of bone via ALN/hydroxyapatite binding, and effectively treat OP in ovariectomized rats. 143 Similarly, Zheng et al. modified platelet-derived exosomes (PL-exo) by conjugating DSPE-PEG–ALN onto their membranes, resulting in ALN-functionalized exosomes (PL-exo-ALN) with enhanced binding affinity to hydroxyapatite and greater accumulation in bone tissue compared with unmodified exosomes. This surface functionalization significantly improved their bone-targeting specificity and therapeutic efficacy in a glucocorticoid-induced OP model. 144
Based on the EV-material hybridization and delivery platform strategy, incorporation of exosomes into hydrogels or scaffold matrices provides a practical route for sustained local delivery: hydrogel-encapsulated, miRNA-enriched exosomes exhibit prolonged bioavailability, enhanced angiogenesis and osteogenesis, and superior defect healing in multiple preclinical models. 145 A study by Le et al. demonstrated the effect of mSC-Exos encapsulated in GelMA hydrogel scaffolds with enhanced stability, osteogenic differentiation, and improved angiogenesis in preclinical bone defect models, along with favorable immune modulation through macrophage polarization. 146 Together, these findings underscore the value of engineering strategies in generating miRNA-enriched exosomes with improved osteogenic potential for skeletal repair.
Conclusion and Future Perspectives
Exosomal RNAs, including miRNAs and mRNAs, play pivotal roles in regulating bone formation and remodeling by mediating intercellular communication and modulating key cellular signaling pathways. These RNA-loaded exosomes influence osteogenesis by targeting genes involved in proliferation, differentiation, and mineralization of osteoblasts and other skeletal cells. While exosome-based therapeutic approaches have demonstrated significant potential in promoting bone repair and regeneration, numerous challenges remain to be overcome. These include the need to identify bone-specific exosomal RNA signatures, optimize methods for exosome isolating, loading and delivery, and clarify the mechanisms of RNA sorting, release, and uptake within the bone microenvironment. The integration of exosomal RNA-based approaches with current regenerative technologies—such as stem cell therapy, biomaterials, and tissue engineering—could revolutionize the treatment landscape for bone diseases. However, the successful clinical translation of such therapies will require robust preclinical data, scalable production methods, and standardized quality controls. Continued research into the biological functions and engineering of exosomal RNAs is therefore essential. These nanocarriers hold immense promise as next-generation tools for enhancing bone regeneration and offer a novel, multifaceted platform for therapeutic innovation in bone disorder and orthopedics.
Authors’ Contributions
Investigation, F.G.S. Writing—original draft preparation and visualization, F.G.S. Writing—review and editing, A.M., M.B., A.E., S.S., and M.H. Supervision, N.B. All authors have read and agreed to the published version of the article. F.G.S., M.B., and N.B. are from the Tehran University of Medical Sciences (Tehran, Iran), and A.M., S.S., and M.A. are from Shahid Beheshti University of Medical Sciences (Tehran, Iran), both where education and research are the primary functions. A.E. is employed outside of Iran.
Footnotes
Acknowledgments
The authors acknowledge the use of ChatGPT solely for superficial language editing.
Author Disclosure Statement
The authors declare no conflicts of interest.
Funding Information
This research received no funding.
Author Confirmation Statement
Drs. F.G.S., M.B., and N.B. are from the Tehran University of Medical Sciences (Tehran, Iran), and Drs. A.M., S.S., and M.A. are from Shahid Beheshti University of Medical Sciences (Tehran, Iran), both where education and research are the primary functions. Dr. A.E. is employed outside of Iran.