
Abstract
The thyroid gland is an endocrine organ responsible for production of triiodothyronine and thyroxine, essential hormones that regulate human metabolism. A wide range of conditions can impair its function, leading to potential life-threatening consequences such as myxedema coma. The standard treatment for hypothyroidism is lifelong levothyroxine supplementation, which, despite being a significant therapeutic breakthrough, has notable limitations and does not fully restore quality of life for many patients. Biomimetic thyroid gland has emerged as a promising alternative treatment strategy for patients with hypothyroidism. Most research to date has focused on generating thyroid organoids from primary thyroid cells or stem cells. However, there is growing interest in other approaches, including the use of biomaterials, bioreactors, and 3D bioprinting as potential alternatives or supplementary technologies to the organoids. While in vitro and preclinical studies have shown encouraging results, clinical application of biomimetic thyroid gland requires further studies in several key areas, including long-term functional validation, studies on large animal models, immunological compatibility and scaffold biodegradation, and absence of standardized good manufacturing practice (GMP)-compliant production protocols.
Impact Statement
Endocrine gland regeneration remains a significant challenge in tissue engineering due to their morphological and physiological complexity. Although hormonal replacement therapy with levothyroxine has been a major breakthrough in managing hypothyroidism, there is still a pressing need for developing a biomimetic thyroid gland. This comprehensive review examines the rapidly advancing area of thyroid gland regeneration, highlighting the most recent and most impactful developments in both cell biology and bioengineering disciplines. By emphasizing the complementary roles of cellular and bioengineering approaches to thyroid regeneration, the review offers an encouraging outlook for the development potential of functional, physiologically regulated biomimetic thyroid gland.
Introduction
The thyroid gland is an endocrine organ composed of two lobes and an isthmus that connects both lobes in the anterior part of the neck. 1 It is a vital organ responsible for producing triiodothyronine (T3) and thyroxine (T4), hormones essential for development and metabolism regulation. 2 It also produces calcitonin, which regulates calcium metabolism. The complex endocrine function of the thyroid gland and its proper response to thyroid-stimulating hormone (TSH) are enabled by its advanced histological structure, which consists of a collagenous matrix with a dense web of blood vessels and multiple follicles composed of follicular cells that produce hormones and colloid, which is stored inside the follicle as a reservoir of prohormones. 3
The absence of a sufficient amount of thyroid hormones due to malfunction of the thyroid gland is known as primary hypothyroidism. This condition can be caused by several clinical entities, including autoimmune thyroiditis or radioiodine treatment. 4 In addition, thyroid lesions are common findings in ultrasound imaging, with studies indicating that such phenomena may affect up to 67% of the population at some point. 5 Consequently, surgical removal of thyroid, otherwise known as thyroidectomy, is a common procedure performed for various indications. These include benign disorders such as multinodular goiter, toxic adenomas, and malignant thyroid cancers, including anaplastic thyroid carcinoma. 6 Total thyroidectomy leaves the patient without an internal source of T3 and T4. In its natural course, hypothyroidism can result in severe complications, including infertility, abnormal bleeding, renal dysfunction, or even the life-threatening myxedema coma. 7 The significant consequences of hypothyroidism resulted in development of hormonal replacement therapy with levothyroxine (LT4) back in the 1950s. 8 Although major therapeutical breakthrough, LT4 monotherapy, despite complex recommendations for therapy management, still causes some noticeable drawbacks (Fig. 1). It has been asserted that the definition of euthyroidism cannot be determined by single value given that the physiological levels of hormones are tissue specific. 9 Furthermore, up to 20% of patients may receive inappropriate or excessive treatment, resulting in emergence of adverse effects and significant decline in the quality of life. It is important to notice that LT4 supplementation requires additional adjustments to take into consideration intricate relationships between free T3, free T4, and TSH. The additional requirements for monitoring of hormonal levels and adjusting therapy have been shown to result in diminished adherence to the treatment and suboptimal outcomes. 10
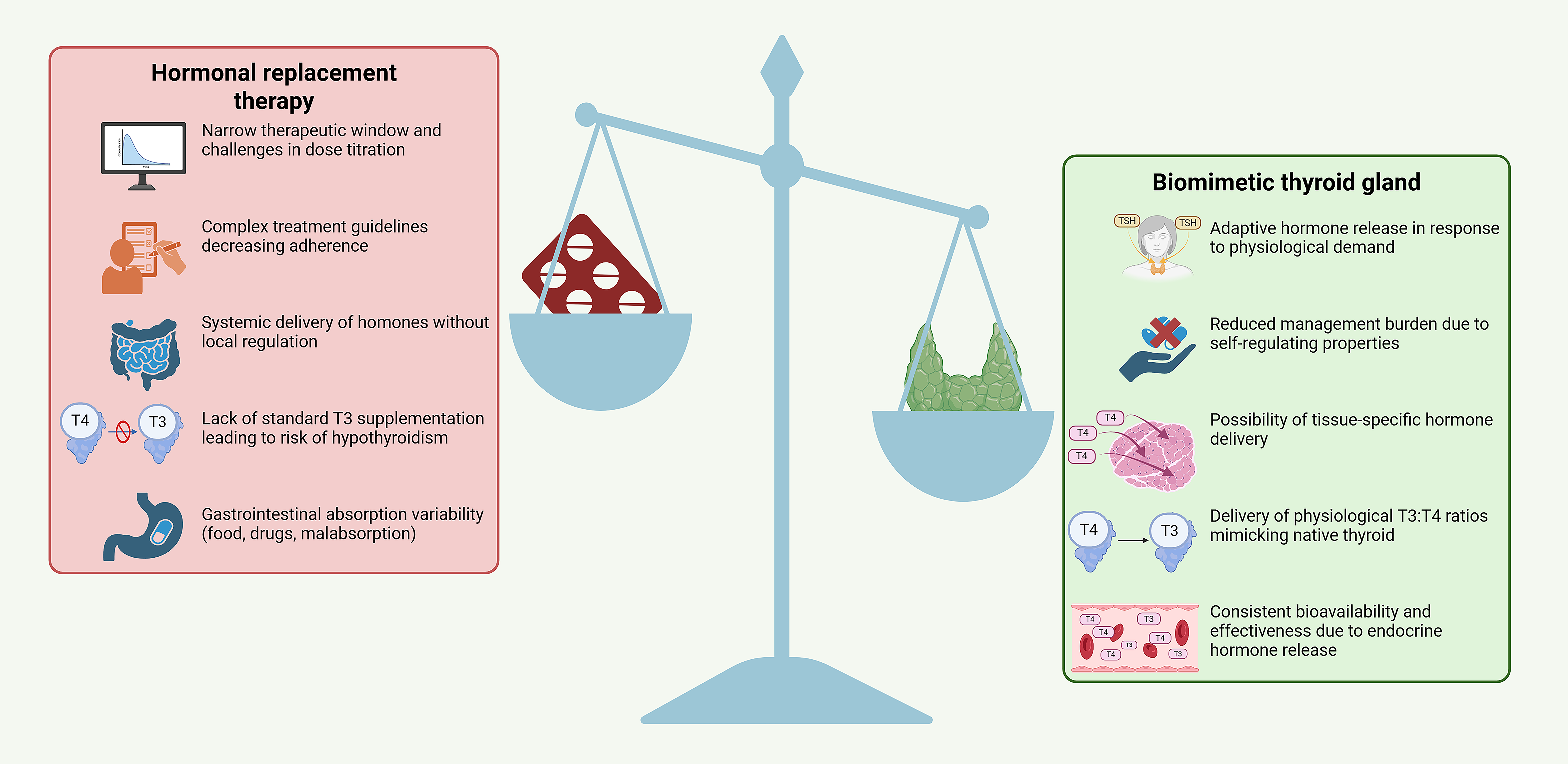
Comparison of hormonal replacement therapy and potential biomimetic thyroid implants. Created with Biorender.com.
The inherent limitations of LT4 therapy have stimulated substantial scientific interest in developing a functional biomimetic thyroid implant that can replace an absent or nonfunctional native thyroid gland. Such a construct should respond to hormonal stimulation in a manner analogous to the native gland, thereby restoring physiological hormonal balance. This review provides a comprehensive analysis of the current advancements in thyroid gland regeneration, with a focus on novel emerging technologies that expand upon progress in thyroid organoid development.
Thyroid Organoids: Main Theme in the Thyroid Regeneration Field
Given the thyroid glands’ extremely long cell turnover rate of approximately 10 years and its lack of intrinsic regenerative capability,11–14 thyroid gland regeneration represents a significant challenge for the scientific community. However, rapid development in stem cell research has enabled the development of thyroid organoids, currently representing the most advanced attempt to recreate a functional artificial thyroid capable of replacing damaged or missing tissue. 15 Organoids are three-dimensional (3D) cellular structures that form through self-organization. They can be created from various types of stem cells and are capable of mimicking essential functions of their tissue of origin. 16 Two primary categories of thyroid organoids have been developed: primary organoids, derived by culturing native thyroid tissue, and stem cell-derived organoids, generated by differentiating embryonic stem cells (ESCs) or induced pluripotent stem cells (iPSCs).17,18 Summary of thyroid organoid generation is presented in Figure 2.
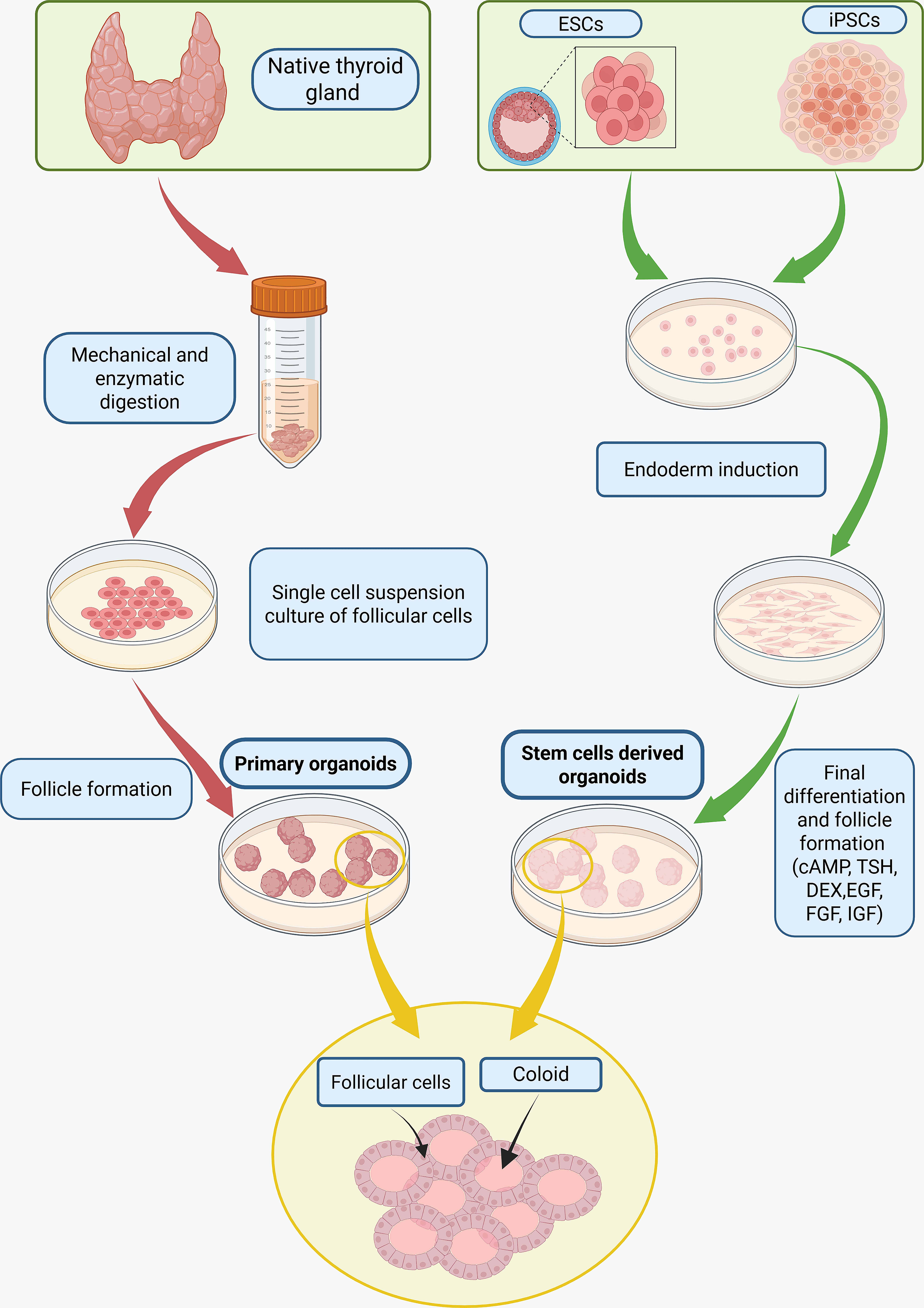
Main sources and methods of thyroid organoids generation. Created with Biorender.com.
Primary thyroid organoids
The generation of thyroid organoids from primary thyroid cells has been reported in several studies involving human, mouse, and rat tissue. This process typically involves mechanical and enzymatic digestion of native tissue to obtain a homogenous cell suspension, which is then cultured on basement membrane extracts (BMEs) using selective cell culture media. It was presented that the self-organization of thyroid cells into follicle-like structures can be enhanced through various methods. Early studies highlighted the importance of supplementing culture media with keratinocyte growth factor and the beneficial effects of a microgravity environment. 19 Current protocols often involve highly complex, heterogeneous culture media supplemented with diverse growth factors, such as epidermal growth factor (EGF) and fibroblast growth factor (FGF), as well as proteins and signaling molecules like Noggin, R-spondin1, and TSH. 20 Analyses of these organoids provided evidence of the proliferative capabilities of thyrocytes both in terms of building of the organoid itself and in terms of the demonstration of self-renewal properties. Moreover, organoids cells exhibit expression profiles of key molecular markers analogous to the native thyroid cells. Functional assessments indicate that these organoids are capable of hormone production and can respond to changes in TSH concentration in a manner similar to native thyroid glands.20–23 In addition, the long-term stability of organoid cultures—over periods exceeding 1 year—has been demonstrated. 20 Taking into consideration potential long-term stability, proliferative properties, and retained in vitro physiological functions, primary thyroid organoids are presented both as model to study thyroid gland biology and as a potential source of material for thyroid gland regeneration. To date, only two in vivo studies have evaluated the implantation of primary thyroid organoids in animal models. Both experiments used mice with induced hypothyroidism. Moreover, the implantation of organoids was performed in the same location, namely, beneath the kidney capsule. The first study results demonstrated the successful implantation of murine organoids under the kidney capsule, as confirmed after period of 4 weeks through the utilization of hematoxylin and eosin staining and immunofluorescence as well as augmented 125I uptake in the area of organoid implantation. Unfortunately, no functional studies were conducted. 24 The second study provided evidence of successful transplantation of 600,000 human and murine organoids. In the course of long-term observations, scientists have demonstrated time-dependent increase in the number and size of thyroid follicles in the histological samples, as well as an increase in the expression of key thyroid proteins. Functional testing showed a moderate increase in the levels of T4 that authors attributed to low quantity of implanted organoids as well as the absence of neovasculogenesis at the implantation site. 25
Stem cells-derived organoids
Compared with primary thyroid organoids, stem cell-derived organoids are relatively better characterized, with greater number of organoids’ generation protocols developed and greater number of in vivo studies conducted. This discrepancy may be attributed to the augmented potential for practical applications in regenerative medicine as well as enhanced accessibility of diverse stem cell types possessing almost infinite proliferative capacity.
Thyroid organoids derived from stem cells can be achieved through the utilization of two stem cell types, iPSCs and ESCs. Numerous studies involving human and murine iPSCs have demonstrated their ability to differentiate into thyroid follicular cells via a multistep process. This typically includes the overexpression of PAX8 and NKX2-1, followed by endodermal induction and final differentiation into follicular cells—commonly cultured on growth factor-reduced BMEs. 26 Murine-differentiated iPSCs demonstrated significant increase in the expression of thyroid-specific proteins (SLC5A5 -NIS, TSHR, Tg, and thyroid peroxidase [TPO]), functional response to TSH, and ability to form follicles by approximately 50% of cells. 27 Research conducted on human iPSCs has yielded similar outcomes to mouse counterpart. However, a higher degree of efficiency has been demonstrated in the follicles formation, and the expression of sodium/iodide symporter protein encoded by SLC5A5 gene (NIS) was only detectable in limited population of studied cells. The in vivo analysis of murine iPSCs-derived thyroid organoids was limited to observation of follicles formation and thyroglobulin (TG) deposition inside them after 4 weeks of implantation. In case of differentiated human iPSCs, the results demonstrated that following transplantation after approximately 4 months, between 63% and 74% of the implanted cells still exhibited coexpression of PAX8 and NKX2-1 and formed follicular structures filled with TG-positive colloid. It is important to note that neovasculature development was observed in the graft. Notwithstanding the fact that the presence of vasculature was a highly positive indicator of successful engraftment, there was no increase in T4, and decrease in TSH levels was recorded. The absence of hormonal activity was attributed to the low expression profile of SLC5A5 in organoid cells, indicating necessity for the incorporation of additional maturation steps into the protocol.28,29
ESC-derived thyroid organoids have been developed using two principal strategies. The first one, known as forward programming, involves direct induction of key transcription factors (TFs) overexpression through the utilization of genetic engineering tools. The second approach, called directed differentiation, revolves around obtaining the final phenotype by series of changes in cell culture medium composition and introduction of various TFs, proteins, and small particles to replicate natural process of differentiation.30,31 A number of protocols for generation and enrichment at various stages of the process have been developed, presenting different levels of differentiation efficiency oscillating around 50%.32–35 All organoids displayed expression of key thyroid-specific proteins, follicles’ formative abilities on BME, TG accumulation in spaces corresponding to lumen of native thyroid follicles, and polarization of hormone transport-related proteins similar to native thyrocytes. Studies conducted on ESCs-derived thyroid organoids frequently presented protein expression level that was analogous to or even superior to those observed in native thyrocytes from the respective species. 35 With regard to their functional properties, organoids displayed active iodine organification.35–37 Notwithstanding the considerable amount of molecular and morphological similarities to native thyroid grand only in three instances, authors reported expression of T4 by the generated organoids.38–40 It was also posited that T4-producing organoids could be obtained from fetal human thyroid cells. 41
In vivo studies have shown that murine ESC-derived organoids implanted under the kidney capsule could partially restore thyroid function in hypothyroid mice. Then organoids exhibited partially retained histological structure. It was also noted that a neovascularization process was taking place, as evidenced by the formation of angiofollicular units—the fundamental functional thyroid units. In addition to structural and molecular similarities to native thyroid gland, organoid grafts were found to be capable of assuming physiological functions in a mouse model of hypothyroidism. Despite the constrained timeframe for observation, studies demonstrated potential normalization of T4, T3, and TSH levels in subject animals 4–8 weeks after implantation. In addition, the capacity of grafts to regulate accordingly T4 levels based on artificially changed TSH levels was demonstrated. The restoration of hormonal levels by means of organoid grafts resulted in the normalization of subject’s body temperature, thereby providing evidence of successful hypothyroidism rescue. The longest reported period of observation has shown successful normalization of hormone level for 29 weeks.36,39 To date, there has been only one available study on the subject of in vivo tests of human ESC-derived thyroid organoid. In a manner consistent with the findings of other studies, organoids were grafted beneath the kidney capsule in a mouse hypothyroidism model. Following a period of 5 weeks, histological analysis of the graft revealed the presence of multiple small thyroid follicles with network of small blood vessels in close proximity, indicating formation of angiofollicular units. In addition, observable differences in cell morphology indicated that the production of hormones was occurring in a subset of follicles. Gene expression analysis demonstrated expression profile consistent with that of mature thyrocytes, with approximately 58% of cells producing T4. The biochemical analysis provided data confirming iodine uptake by graft area, normalization of serum T3 levels, and increase of T4 level. The subphysiological levels of T4 observed were attributed to the relatively brief observation period, with notable weekly tendency for T4 increase. Furthermore, the capacity of grafts to physiologically regulate hormonal response in accordance with the TSH stimulation has been demonstrated. 39
Despite significant advances in understanding thyroid organogenesis and the development of complex stem cell-derived organoids, in vivo testing remains limited. These findings underscore the major challenge of translating in vitro constructs into functionally integrated tissues capable of long-term survival and physiological regulation at the implantation site.17,42–50
Emerging Alternatives and Complementary Techniques for Thyroid Bioengineering
The complex cellular structure of the thyroid gland, combined with the fact that its physiological function is closely linked to its specific architecture, has led in the past decade to the development of numerous concepts—both alternative and complementary to stem cell-derived organoids. A considerable number of publications tackled the problem of thyroid gland regeneration from various perspectives, including adult stem cells, bioreactors, biomaterials, and bioprinting, which was presented in Figure 3.
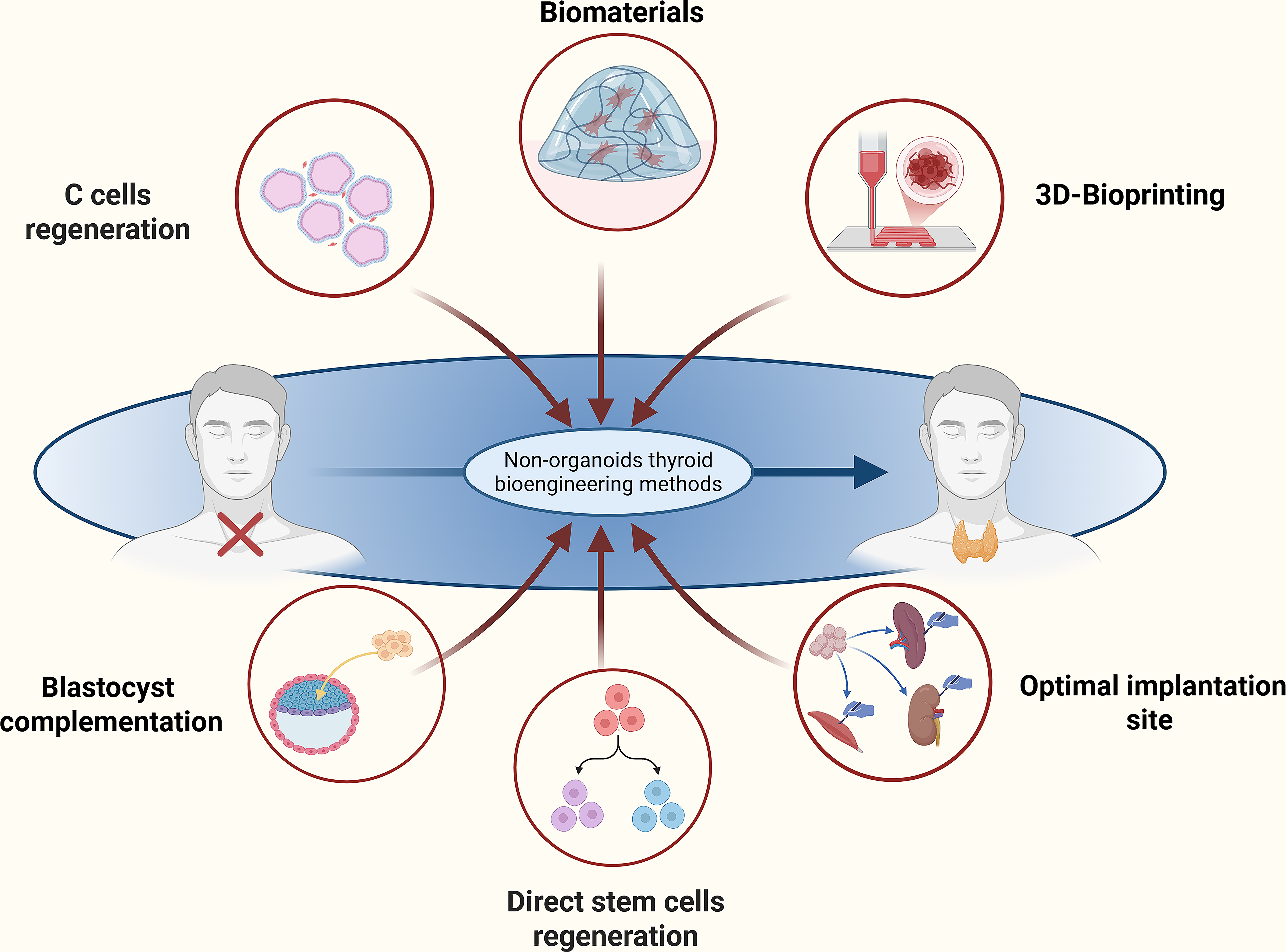
Main complementary and alternative methods in thyroid gland bioengineering. Created with Biorender.com.
Adult stem cells in thyroid regeneration
Recent studies have provided compelling evidence for the presence of a population of progenitor cells within the adult thyroid gland—a possibility that had long been suspected.47,51,52 In an experimental model of severe thyroid injury induced by tamoxifen and diphtheria toxin, researchers observed a rapid recovery of thyroid function within 8 weeks of damage induction. This recovery was attributed to the activation of dormant cells expressing stem cell markers such as Oct3/4, NANOG, and SOX2, as well as progenitor cells characterized by PAX8 expression and the absence of TG expression. It was established that these cells tend to accumulate in close proximity to newly forming follicles. 53 In another study in mice that have undergone partial thyroid lobectomy, a population of thyroid progenitor cells was discovered to form new primitive follicles with present mitotic activity and low levels of TG in the lumen. It has been demonstrated that those primitive follicles exhibited capacity to divide into multiple smaller follicles, which underwent final maturation. This process resulted in the presentation of morphology and expression profile of adult normal thyroid follicle on day 21. 54 Hidden regenerative potential of thyroid follicular cells was also indicated in the study conducted on thyroid cancer metastases, were highly differentiated thyrocytes gained tissue-context-dependent high proliferative capacity independent from TSH presence. This may suggest that this cell population may be the population responsible for metastasis regeneration rather than small number of thyroid cancer stem cells. 55 It is evident that progenitor thyroid cells and residual stem cells are not the only candidates for cellular component of biomimetic thyroid. In 2025, a study was published exploring the application of bone marrow-derived mesenchymal stem cells (BMSCs) in the treatment of Hashimoto’s thyroiditis. The results presented indicated the immunomodulatory potential of BMSCs in reducing inflammation as well as their ability to increase the concentration of growth factors in their immediate proximity and their potential to differentiate into fibroblasts or thyrocytes. 56
Following their recent identification and characterization, dormant adult thyroid stem cells and progenitor cells may represent a viable alternative to primary cells, iPSCs, and ESCs in generation of functional thyroid implants. This potential can be attributed to their higher level of differentiation, ease of accessibility, and possible personalization of the implants based on them. The presence of adult stem cells is known to be critical in maintaining both physiological function and structural integrity in almost every organ throughout the human body’s lifespan. 57 Incorporating such elements into biomimetic thyroid constructs could enhance the differentiation potential of organoids, improve functional performance, and replace apoptotic cells—thereby increasing the longevity and stability of engineered implants.
Whole thyroid gland production in the bioreactor
Blastocyst complementation is a powerful chimeric organ engineering strategy that enables the in vivo generation of various organs and specialized cell types.58–62 The technique involves the microinjection of stem cells into embryos in the blastocyst-stage embryos that have been genetically modified to lack the ability to form a specific organ or to undergo normal organogenesis altogether.63,64 These modified embryos function as biological bioreactors, allowing the development of chimeric organs based on the provided stem cells. There are documented attempts to utilize this method in the fabrication of thyroid tissue and follicular cells (Fig. 4). Two distinct mouse models were generated: one with depletion of FGF10 and the other with NKX2-1 deletion. In the first model, complementation with murine ESCs resulted in development of normal and fully functional thyroid tissue. The majority of generated tissue was comprised of cell derived from the introduced ESCs cells. Although part of the cells, they were originating from the host blastocyst. In the second model, results were similar, with development of thyroid tissue almost entirely derived from injected stem cells.65,66 Despite the substantial promise of blastocyst complementation as a method for thyroid tissue engineering, several critical challenges remain. Chief among them is the necessity to achieve complete purification of the regenerated thyroid gland from host-derived cells in order to prevent immunological rejection during future transplantation. Another major concern relates to the ethical implications of creating interspecies chimeras, as donor-derived cells may integrate not only into the targeted organ but also into other parts of the host organism.
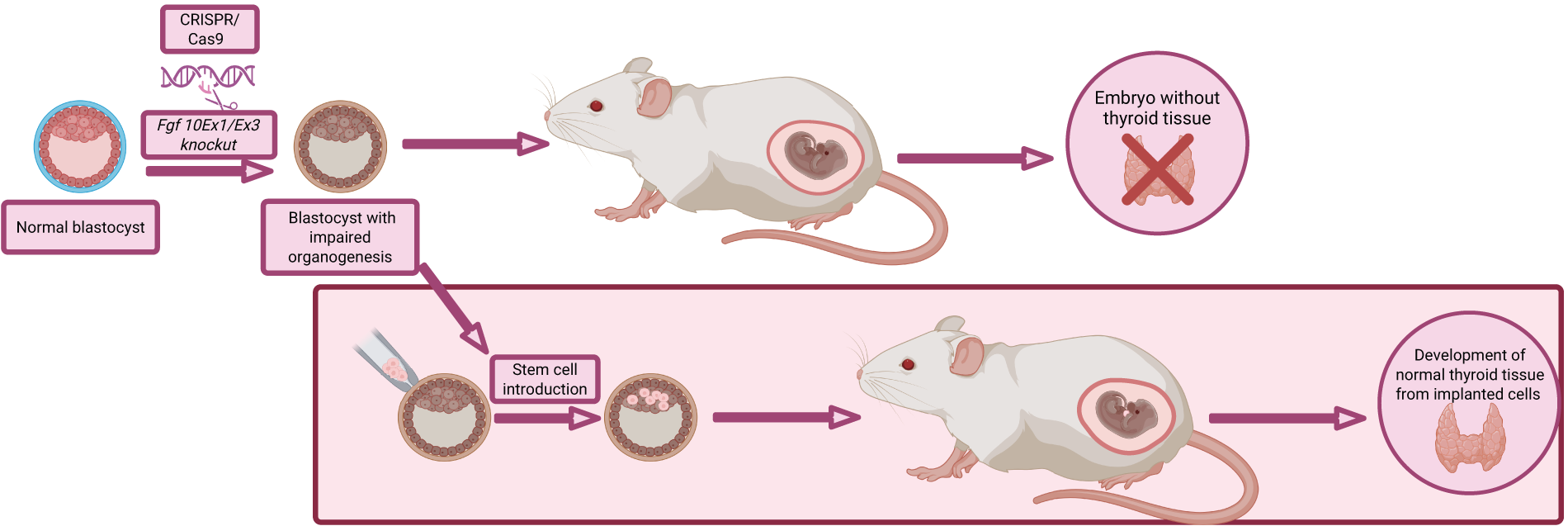
Blastocyst complementation technique in regeneration of thyroid gland. Created with Biorender.com.
Advancements in C-cells regeneration
In the context of thyroid gland regeneration, it is important to acknowledge that the gland is physiologically responsible not only for the production of TG, T3, and T4 but also calcitonin. While calcitonin deficiency is typically compensated by parathyroid hormone and vitamin D, in rare cases, the absence of calcitonin—produced by parafollicular cells (C-cells)—can lead to increased bone resorption. This may result in osteopenia and elevate the risk of bone fractures. 67 In order to create a fully functional artificial thyroid, it is essential that it possess all of the necessary functionalities. To the best of our knowledge, currently only one study tackled the subject of C-cells regeneration. The utilization of human ESCs has enabled the development of the protocols for both two-dimensional (2D) and 3D differentiation into C-like cells. The subjected cells were differentiated through the implementation of forward programming approach and were split into two parts. The first was oriented in similar fashion to thyroid organoids on generation of definitive endoderm phenotype with activin A and low fetal bovine serum (FBS) concentrations. The second one, the final differentiation into C-cell like cells, was performed by addition of 10% FBS, retinoic acid, and insulin-like growth factor-1. Cells differentiated through this protocol were capable of synthesizing calcitonin and exhibited a functional response to changing calcium concentrations in the culture medium. Interestingly, while physiologically being scattered across all of the thyroid gland in the in vitro environment, 3D model presented higher expression of calcitonin in comparison with plain 2D cells. 68
Implantation site: optimal environment for graft integration
Given the thyroid glands’ dense internal vasculature, multiple arterial blood supplies, and its intrinsic endocrine function, which by definition depends on the circulatory system, it is imperative not only to restore the functionality of arteriofollicular units but also to ensure sufficient vascularization to support the high metabolic activity of a thyroid implant. 69 It is therefore crucial to determine optimal implantation site. Over the years, researchers have explored several potential locations for thyroid autotransplantation. In human subjects, these procedures have been performed predominantly within various muscles, including sternocleidomastoid, rectus femoris, or the forearm muscles. 70 Although initially very promising, thyroid autotransplantation frequently failed to result in hypothyroidism rescue or did so just for a brief period of time. 71 This phenomenon may be partially attributed to the insufficient vasculature and regenerative potential of muscle tissue impeding the integration process. In recent years, in vivo testing of thyroid organoids has been consistently performed by implanting them into the subcapsular space of the kidney. 26 This approach is associated with several advantages over intramuscular implantation. The primary one revolves around high vascularization of the kidney capsule, providing some degree of protection for the graft as well as facilitates easy access for the operator. 72 A novel potential implantation site has recently been proposed for thyroid regeneration: the spleen. In a mouse model of thyroid autotransplantation, intrasplenic transplantation was shown to support both the growth and function of thyroid follicular cell. Engraftment of 1.5 g of mouse thyroid minced into small blocks of tissue suspended in the hyaluronic acid resulted in complete integration of implanted tissue with spleen stroma. Within just 2 days posttransplantation, restoration of arteriofollicular units was obtained. Furthermore, this massive blood vessels proliferation was not accompanied by the inflammatory response. Physiological hormonal balance was restored within 4 weeks, along with measurable improvements in cardiac and hepatic function. Following the observation period, the data indicated a potential increase in the thyroid tissue mass in the spleen compared with its initial weight. This finding suggests that the spleen environment may possess proliferative stimulating properties. A comparative study demonstrated the superiority or intrasplenic implantation compared with intramuscular as evidenced by the occurrence of necrosis, and decline in the number and volume of thyroid follicles was observed. 73
Biomaterials in thyroid gland regeneration
The extracellular matrix (ECM) is a complex network of proteins and glycans that forms the scaffold surrounding and supporting nearly all cells in the human body. 74 In addition to providing tissues with physical structure and mechanical integrity, the ECM plays a key role in regulating critical cellular processes, including proliferation, differentiation, and migration.74,75 The complex composition of the ECM provides surrounding cells with a stable environment, thereby enabling them to retain their phenotype and proper physiological functioning. The gold standard for culturing stem cells and developing 3D cellular structures, both spheroids and organoids, is the use of commercially available ECM substitutes. These are typically BMEs presented as specialized hydrogel solutions, with Matrigel being the most widely used example. 76 All of the aforementioned principles were utilized as the foundation for the development of both primary and stem cells-derived thyroid gland organoids’ development. 77 In recent years, there have been significant advancements in integrating thyroid organoids into lab-on-a-chip platforms designed for in vitro studies of thyroid pathophysiology and pharmacological testing.78–81 The development of these devices was based on the microfluidic technology, a sophisticated system that enables precise manipulation of small fluid volumes and simulates dynamic physiological conditions. 82 The incorporation of a dense, predominantly collagenous ECM substitute as a support for organoids constituted a significant technical challenge. However, this notion can be surpassed by conventional enzymatic extraction of organoids from the BMEs and subsequent suspension in cell culture medium. This approach was unfortunately found to be inefficient due to the fact that organoids deprived of the contact with ECM substitute tend to degenerate and lose their organized follicular architecture. 78 Taken into consideration results of studies on microfluidics devices, there is growing body of evidence that the role of interaction between cells and the ECM in thyroid regeneration may be more significant than was previously anticipated, and the absence of collagenous support may negatively impact results of in vivo studies.
The growing recognition of the ECM as a key component in the bioengineering of functional organs has led researchers to search for safe and renewable sources of ECM suitable for clinical use. Many currently available BMEs are derived from tumorigenic cell lines, raising safety concerns for their use in implantable grafts. One potential answer to this challenge may be the development of a range of techniques for decellularization of various tissues and organs.83–88 Decellularization is defined as the process of removing cellular content from a desired tissue without damaging the native architecture of ECM. 89 Meticulous processing, including mechanical, enzymatic end chemical methods, provides scaffolding biomimetic material that is primed for recellularization. Based on the hypothesis that the incorporation of thyroid ECM into thyroid grafts may positively impact cellular component of the graft, several protocols for thyroid tissue decellularization have been developed. The first report on the generation of a decellularized rat thyroid was published in 2012. However, there is a paucity of data concerning the outcome of the decellularization and subsequent recellularization. 90 A more comprehensive dataset emerged in 2019, describing a flow-based chemical decellularization protocol for an entire rat thyroid lobe. The biomaterial obtained in this study met all of the canonical decellularization criteria. In addition, it was found to retain the mechanical properties and vasculature bed of the native material. In the functional studies, the authors using mesenchymal stem cells and FRTL-5 rat epithelial thyroid cells demonstrated lack of cytotoxic effect of the scaffold on the cells. Furthermore, it has been demonstrated that to some extent scaffold promoted the growth of stem cells. Flow recellularization of the scaffold with FRTL-5 cells using the same blood vessels that were used for decellularization was found to be successful. Histological analysis revealed formation of cellular spherical structures around former follicles’ ECM. In addition, cells retained the expression of crucial thyroid genes, TPO and TG. 91 In the same year, another group conducted an extensive mass spectrometry analysis of the decellularized rat thyroid matrix. They developed a reproducible protocol to assess the retention of collagen types I through V after the decellularization procedure, which may be used in the future for the standardization and quality control of mass-produced scaffolds. 92 The subsequent approach to this subject was published in the 2021 and was concentrated on the rabbit decellularized thyroid. The authors provided detailed characteristics of the scaffold and for the first time demonstrated that after the decellularization, a significant amount of key growth factors, transforming growth factor β (TGF-β), vascular endothelial growth factor (VEGF), FGF, platelet-derived growth factor (PDGF), and hepatocyte growth factor, were to some extent preserved in the scaffold. This finding further substantiates the viability of this type of scaffold for thyroid bioengineering. The cellular test showed that the scaffold promotes the proliferation of the human follicular cells. However, a decline in expression of TPO after 7 days was observed, which was attributed to lack of hormonal stimulation during the cell culture. 93 In 2024, the first protocol for the decellularization of human thyroid tissue was published. In this study, authors provided comprehensive comparison between the results of various approaches on the efficiency of decellularization process in this type of material. Cell seeding experiments were conducted using human thyroid cancer cell line and provided evidence of cytocompatibility of obtained scaffolds with human cells. 94 Later in the same year, another study introduced a novel application of decellularized ECM (dECM) by developing injectable hydrogels derived from porcine thyroid tissue. These bioinks retained glycosaminoglycans and growth factors from the native ECM and exhibited concentration-dependent capabilities to promote cell proliferation and hormone production. 95 All abovementioned advancements in thyroid dECM research were summarized in Table 1.
Summary of Advancement in Thyroid dECM Research

dECM, decellularized extracellular matrix; FGF, fibroblast growth factor.
It has been established that the vast majority of cells possess the capacity to synthesize a minimal amount of ECM in their immediate surroundings. This process facilitates cell-to-cell adhesion and allows formation of 3D cellular structures. 96 This fact was instrumental in the development of the cell sheet technique, were in vitro cultured cells form cohesive, dense cellular unit that can be transferred to the desired location, thereby providing high number of cells. 97 The potential of this technology in the regeneration of the thyroid gland was also the subject of the study. Two-dimensional culturing of primary rat thyroid follicular cells on thermo-responsive plates has been shown to result in the formation of thick, cohesive cell sheets containing approximately 1 × 107 cells. Histological analysis of the sheets’ morphology unveiled the presence of small central follicles containing TG colloid, and immunohistochemical staining demonstrated the retention of NKX2-1 expression. The subcutaneous implantation of thyroid cell sheet in athymic mice resulted in the appearance of detectable levels of both T3 and T4, which did not reach the physiological levels. These findings suggest that this form of bioengineered thyroid tissue may, with further refinement, evolve into a clinically viable therapy. 98
Another promising approach to thyroid bioengineering involves the use of microencapsulation technologies to generate individual hormone-secreting capsules, each functioning as an independent thyroid unit. In the study, porcine primary thyroid cells were encapsulated in the alginate and poly-
Despite being in the early stages of development, scaffold-based techniques show considerable promise in the field of thyroid bioengineering—not only for their ability to enhance the mechanical integrity of engineered grafts, but also for introducing additional layers of molecular regulation that support and modulate the behavior of embedded cellular components.
3D bioprinting
To the best of our knowledge, as of today, the topic of thyroid bioengineering has been discussed in the context of 3D bioprinting on only a single occasion. This limited attention may stem from the insufficient resolution of current 3D bioprinting technologies and the highly complex structure of the arteriofollicular unit, which remains challenging to reproduce artificially. The objective of the referenced study was to apply 3D bioprinting technique to create a functional, vascularized mouse thyroid construct, utilizing a technique similar to the now widely recognized Kenzan method. This method revolves around the precise deposition of cellular spheroids, which form the primary architecture of the tissue, followed by a maturation period during which the spheroids fuse and reorganize into a final tissue-like structure. 100 To replicate native architecture of the thyroid, the authors used two types of spheroids: one derived from embryonic thyroid explants and the other from the allantois. Based on the prepared in silico mathematical model and the visualization of spheroid fusion, the scientists designed the construct and fabricated it using a specialized 3D bioprinter. After a 4-day maturation period, the constructs exhibited early signs of endothelial cell differentiation and the formation of primitive vascular structures. In addition, the formation of thyroid follicles was also noted. When the bioprinted grafts were implanted under the kidney capsule in mice, an 8-week follow-up revealed a significant increase in circulating T4 levels and normalization of body temperature, indicating a partial reversal of hypothyroidism. 101 These findings demonstrate that 3D bioprinting holds considerable promise as a tool for the future development of bioengineered thyroid glands, potentially enabling the precise reconstruction of complex glandular architecture and vascular systems necessary for functional endocrine integration.
Conclusions
Although current approaches are in early development, future biomimetic thyroid implants may address several critical clinical needs. Most notably, they could serve as a treatment option for patients who have undergone total thyroidectomy due to thyroid cancer and are dependent on lifelong hormone replacement. Another important group includes children with congenital hypothyroidism, particularly those who exhibit poor compliance or fluctuating hormone levels despite standard substitution therapy. Moreover, patients suffering from autoimmune thyroiditis (e.g., Hashimoto’s disease) with progressive tissue destruction and suboptimal control on LT4 alone may benefit from tissue-regenerative interventions. Finally, such implants could also offer value in the context of rare disorders of thyroid hormone synthesis or transport, where pharmacological options are limited or ineffective.
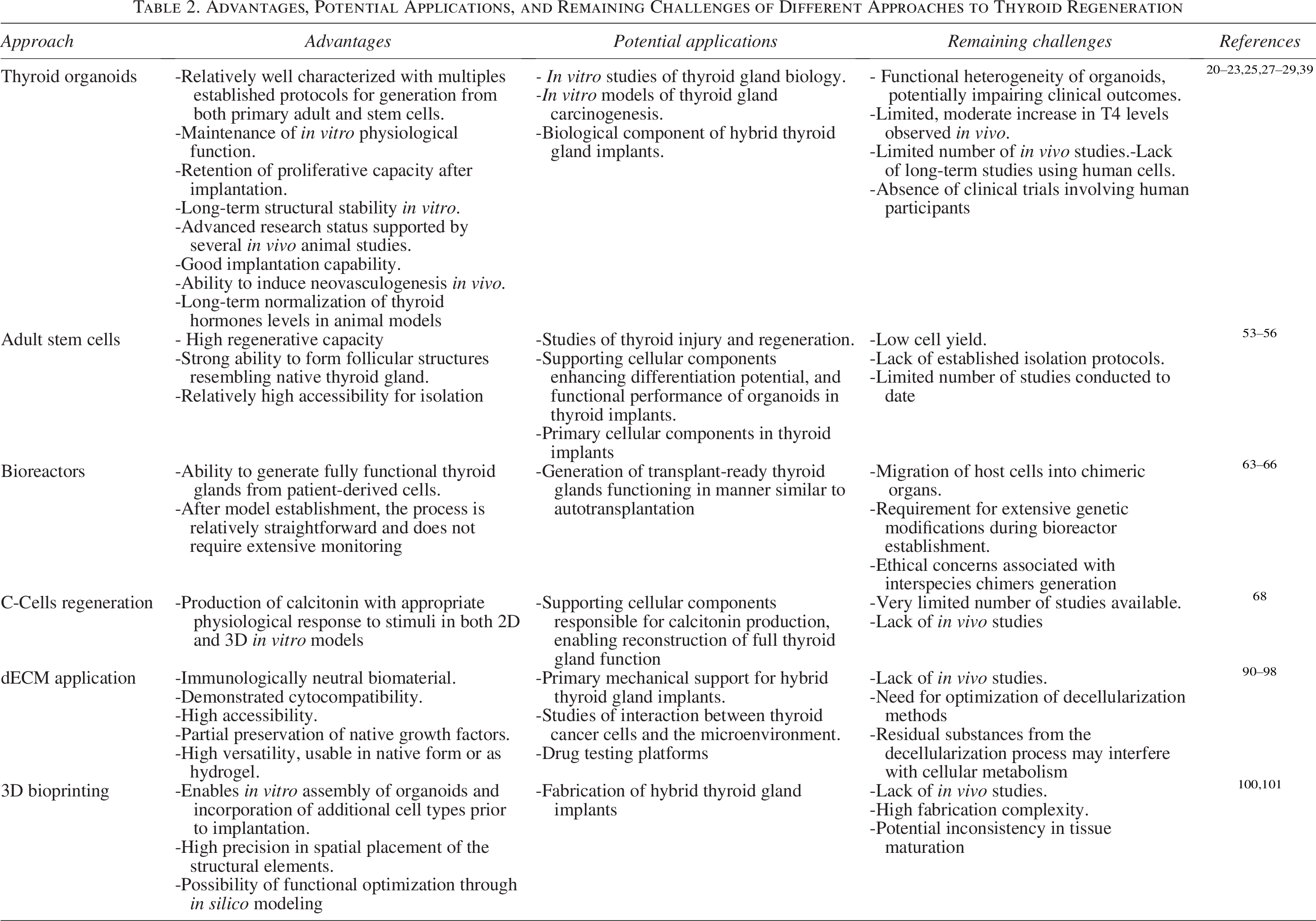
The morphological complexity of endocrine organs and their architecture-dependent physiological functions make them particularly challenging targets for developmental biology and bioengineering research.102,103 The thyroid gland is not an exception to this phenomenon. Despite promising in vitro and preclinical findings, clinical application of biomimetic thyroid implants remains hindered by several critical factors, including incomplete long-term functional validation (hormone levels, responsiveness to TSH), lack of large animal models, unresolved issues of vascularization and innervation, immunological compatibility and scaffold biodegradation, and absence of standardized GMP-compliant production pipelines. The advantages and remaining challenges of emerging technologies used in the field of thyroid regeneration were summarized in Table 2. The development of highly advanced thyroid organoids provides a promising perspective on the cellular component of the ideal implant. However, their functionality has only been demonstrated up to 29 weeks, which is a very short amount of time in clinical settings. The process of implantation of unprotected organoids has been observed to result in disruption of their in vitro structure. A potential consequence of this disruption may be alterations of this phenotype and development of neoplasms. Furthermore, if the primary cells do not originate from the recipient of the implant as in any other type of implantation, there is a high risk of implant being rejected and destroyed by the immune system. Direct regeneration utilizing stem cells presents itself more as a complementary system to the others, thereby facilitating stability and cell turnover in the implant. Recently developed scaffold-based approaches have shown promise in providing mechanical support, delivering growth factors, and restoring cell–ECM interactions. However, these methods have not yet demonstrated the capacity to recreate microvascular arteriofollicular units, which are essential for full physiological function. Looking ahead, optimal outcomes may be achieved by integrating scaffold-free and scaffold-dependent techniques. For instance, thyroid-derived dECM hydrogels represent a hybrid strategy that facilitates easier culturing and administration of follicular cells while simultaneously offering limited mechanical support. Such combinations may offer the functionality, safety, and stability required for clinical application.
Advantages, Potential Applications, and Remaining Challenges of Different Approaches to Thyroid Regeneration
Authors’ Contributions
T.G.: Conceptualization, investigation data curation, formal analysis, writing—original draft, and review and editing. M.G.S.: Writing—original draft. G.R.: Writing—original draft. G.M.F.: Conceptualization, data curation, formal analysis, and writing—review and editing. W.J.: Investigation, supervision, and writing—review and editing. A.C.: Investigation, supervision, and writing—review and editing. P.S.-Z.: Methodology, validation, formal analysis, investigation, writing—review and editing. K.K.: Supervision and writing—review and editing. P.D.: Validation, resources, funding acquisition, supervision, and writing—review and editing. F.V.: Methodology, validation, formal analysis, investigation, writing—review and editing. C.D.M.: Methodology, validation, formal analysis, investigation, writing—original draft, and writing—review and editing. B.K.: Conceptualization, project administration, resources, supervision, and writing—review and editing. G.V.: Conceptualization, project administration, supervision, and writing—review and editing.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.