
Abstract
Introduction:
Long-term storage of bee semen by freezing is a critical process for both the preservation of the genetic material and the sustainability of beekeeping activities. It has been observed that low-density lipoproteins (LDLs) increase sperm quality after freezing and thawing. Although studies have been conducted on the use of LDL for this purpose in different animal species, no research has been conducted on honeybee semen to date.
Objectives:
This study aimed to evaluate the potential effects of using LDL instead of egg yolk (EY) on sperm quality and fertilization rate by examining the effects of different LDL ratios (2.5%, 5%, 10%, 25%) on bee semen.
Methods:
Sperm collection was conducted using a Schley-type device, resulting in six distinct groups, including both no-supplemented and experimental groups. In the first experiment, sperm collected from 36 drones were diluted with varying LDL concentrations before being frozen and thawed; motility, membrane integrity, viability, and longevity were measured. In the second experiment, a total of 56 virgin sister queens, 8 from each group, were inseminated.
Results:
In the group containing 25% LDL, a significant increase was observed in the motility, membrane integrity, and viability rates of frozen–thawed honeybee sperm. In the group containing 25% EY, there was a clear decrease in these parameters; moreover, the lifespan of the sperm was significantly reduced. In the groups, the highest value in terms of fertility was observed in the 25% LDL group, and the lowest value was determined in the 25% EY-added group.
Conclusion:
The findings demonstrated that the addition of 25% LDL significantly enhanced both sperm quality and fertility rate in honeybees.
Introduction
The only living species that pollinates approximately 80% of the world’s plants is the honeybee. 1 Therefore, declines in the bee population have extremely negative effects on the natural ecosystem. 2 The production of qualified queens is directly dependent on the quality of the sperm obtained from male bees. In this context, artificial insemination applications enable the production of high-quality queens. In addition, freezing and preserving drone sperm facilitates its transportation to remote areas, supports the preservation of genetic diversity, and acts as a precaution against infectious bee diseases. 3 Today, the degradation of the bee gene pool is shown as one of the main reasons for the problems related to beekeeping. For this reason, the widespread use of biotechnological applications to protect genetic material is of great importance. Various methods have been developed for the cryopreservation of bee sperm, and studies are ongoing in this field.4–6
Effective storage of bee sperm and fertilization of queens with this sperm provide important contributions not only to the beekeeping sector but also to scientific research. The use of frozen sperm ensures the preservation of select genetic material and reduces the risk of disease spread during the import and export of genetic material. In this process, the content of the protective cryoprotectants used and the application protocols contribute to the sustainability of genetic variation. 7 Low viability rates have been observed in frozen drone sperm, which has negatively affected the fertilization rates of queens. The fertilization rate, which is between 95% and 99% with fresh sperm, drops to 47% with frozen sperm. 6 Various solutions and alternative cryoprotectants are being investigated to overcome this problem.8,9
Egg yolk (EY), which is widely used in sperm extenders, provides a protective effect during freezing by stabilizing sperm membranes. 10 However, some substances contained in EY reduce sperm motility, create structural inconsistencies, and increase the risk of bacterial contamination. In addition, high-density lipoproteins (HDLs) found in EY increase the sensitivity of sperm to cold shock.11–14 For this reason, in recent years, instead of EY, only the protective fractions have been emphasized. Studies have revealed that this protective fraction is primarily composed of low-density lipoproteins (LDLs).15–17 LDL has been obtained by purifying it from EY and has been used in different animal species at various rates, with successful results.18–20 However, the optimum LDL ratio is determined to be dose- and species-specific due to the variability of the sperm membrane. 21 However, to date, no study has been found that has examined the effects of LDL addition on bee sperm quality and fertilization success after freezing. In this context, the present study is the first to systematically examine the effects of different LDL ratios (2.5%, 5%, 10%, 25%) on bee sperm quality and fertilization.
Materials and Methods
Bees, semen collection, and dilution
This study was conducted at Hatay Mustafa Kemal University between May and August 2023. The Hatay Mustafa Kemal University (HMKU) Scientific Ethical Committee approved the experiments. All worker and queen bees used in the experiments were obtained from individuals of the Apis mellifera Anatolian raised at HMKU. Mature male bees, 16 days old, were collected from colonies during their routine daytime flights and placed in common flight cages with conspecific individuals. Sperm collection was performed from 36 mature male bees, following the classical methods described in the literature. 22 The head and thorax were kept fixed, and light pressure was applied to the abdomen; semen was collected under a stereo microscope using a Schley instrument. Queen bees were reared in queenless colonies and inseminated with thawed sperm. Eight queen bees were used in each group. After insemination, the queens underwent wing clipping, labeling, and were placed in three-frame compartment hives containing young worker bees.
The composition of the diluent solution used in this study was adapted from the original formula developed by Hopkins et al. 23 (14.2 mg disodium phosphate, 30 mg sodium citrate, 611.3 mg potassium chloride, 490 mg sodium chloride, 42 mg sodium bicarbonate, 687.8 mg TES buffer, 363.4 mg Tris base, 0.37 mg EDTA, 0.07 g penicillin, and 0.05 g streptomycin in 100 mL of distilled water). All chemicals were obtained from Sigma (St. Louis, MO, USA).
Extraction of LDL
According to the method described by Moussa et al., 24 the LDL fraction was obtained by separating the EY. The EY was diluted 1:1 with 0.17M sodium chloride solution and then centrifuged at 10,000 g for 45 minutes at 4°C. The upper phase was carefully removed and centrifuged a second time. The plasma obtained was mixed with a solution containing 40% (w/w) ammonium sulfate (1.01217) to achieve pH 8.7. Ammonium sulfate was added to this mixture at a 1:1 ratio, and the process was continued until the formation of a precipitate was observed. The precipitate and the supernatant were separated by centrifugation at 10,000 g for 45 minutes. The supernatant phase obtained was dialyzed against distilled water (D6066) for 12 hours at 4°C, changing every 2 hours, to remove ammonium sulfate. After dialysis, it was centrifuged again to obtain the upper phase rich in LDL. This phase was divided into small volumes and stored frozen at −80°C for future use.
Experimental design
Freshly collected sperm samples without any treatment were evaluated as the positive control for fertility. Frozen and thawed sperm without any additives was the negative control (LDL 0%). The other groups were prepared by adding 25% EY, 2.5% LDL, 5% LDL, 10% LDL, and 25% LDL to the diluents, respectively.
Freezing and thawing
The collected semen was first mixed with the diluent solution prepared in proportions appropriate for the experimental groups. In this mixture, two units of diluted solution were added to three units of sperm sample, and the resulting homogeneous mixture was then combined with the specially prepared cryoprotectant solution. The cryoprotectant solution consists of a 75% phosphate buffer, which contains 79.7 mM NaH2PO4 and 31.6 mM Na2HPO4, its volume is completed with distilled water to 25 mL, and the pH level is adjusted to 7.2 with 6M NaOH. In addition, 10% DMSO was added to this mixture. The prepared sperm–cryoprotectant mixture was transferred to special 0.25 mL straws. The ends of these straws were heat-sealed to ensure liquid tightness. Each straw was then placed in a water bath inside the refrigerator to slowly cool the samples to 4°C and was subjected to an equilibration process for approximately 2 hours. The obtained straws were placed vertically in the cryo chamber (Crysalys Cryocontroller, PTC-9500). The cooling protocol was programmed to reach −40°C with a decrease rate of 3°C per minute, starting from 5°C, and was kept constant at this temperature level for 5 minutes. After the cooling process was completed, all straws were transferred to the liquid nitrogen tank and stored for about 2 weeks until insemination at −196°C.
Each straw was removed from the liquid nitrogen and immersed in a water bath prepared at 35°C for 20 seconds to thaw.
Spermatologic evaluation
Motility
Motility assessment was performed by phase-contrast microscopy from three different areas. These observations were made by only one expert observer for all samples. Motility rate was scored between 0% and 100% based on visual observation at ×400 magnification.
Viability
The eosin-nigrosin staining based on the method developed by Björndahl et al. 25 was applied to determine the dead/live sperm. A total of 300 sperm cells were counted on each slide at 1000× magnification.
Membrane integrity
The method is based on the procedure developed by Jeyendran et al. 26 and was used in this study with minor modifications. For the analysis, 10 µL of semen sample was mixed with 100 µL of hypoosmotic solution (1.1 g of fructose and 0.55 g of sodium citrate in 100 mL of distilled water) and incubated at 37°C for 60 minutes. After incubation, a total of 200 sperm cells in each sample were examined under a phase-contrast microscope. In cells with intact membrane functions, typical swelling and curling of the tail were observed at ×400 magnification.
Longevity
The time-dependent motility decreases of sperm samples were analyzed using a phase-contrast microscope and on a heated microscope table fixed at 37°C. The first evaluation was made immediately after the sperm was thawed. Motility examinations of sperm samples were performed at hour zero. Then, samples were taken on a 10 µL Makler slide at 1-hour intervals, and 10 squares were counted starting from top to bottom or left to right under a phase-contrast microscope (Olympus CX21, Japan) at 20× magnification. Motile spermatozoa with strong, smooth, and linear forward movement and immobile spermatozoa were counted in the field. This process was repeated —three to four times, and motility was calculated as %. The durability period was evaluated until motility ended, and this period (hour) was recorded.
Fertility
A total of 56 virgin queen bees were reared under special conditions. Eight queen bees, randomly selected to represent each group, were artificially inseminated with previously frozen and thawed sperm. Insemination was performed using a Schley device (Schley, Germany), using standard techniques and a 5 µL volume of semen sample. Immediately after insemination, the right wing of each queen was carefully cut to prevent flight; an identifying plastic numbering disk was placed on her and marked. These queens were then equipped with young worker bees and placed in three-frame nucleus hives. For the queens to get used to the hives before they came into direct contact, they were kept in special queen cages. After 24 hours of insemination, each queen was briefly treated with CO2 to stimulate the egg-laying reflex. 27 After a 3-day adaptation period, the queens were released from their cages and allowed to interact freely with the colony. Following the initiation of the egg-laying process, the larvae hatched from the eggs were allowed to reach the pupal stage, and the comb cells were allowed to close. After the brood development was completed in all colonies, the closed brood areas in each comb were documented with high-resolution photographs. These images were used, and counting operations were performed via ImageJ. 28 The fertility rate was calculated by counting the worker bee pupae in three distinct locations chosen for each mini hive.
Statistical analyses
The normality of the data distribution was assessed before statistical analysis. All statistical analyses were performed using SPSS 23. One-way analysis of variance was used to detect differences between groups. The Duncan test was applied to determine statistically significant differences between the groups. Experiments regarding sperm freezing–thawing processes were performed with 10 repetitions, and fertility assessments were performed in 4 repetitions.
Results
Sperm motility
In fresh semen, motility was 95 ± 5.0% (±standard error [SE]). After freezing and thawing, significant differences in motility were observed between the groups (Table 1). The highest motility was in the 25% LDL group (90.6 ± 2.8%), which was significantly higher than in the 0% LDL and 25% EY groups (p < 0.05) (Table 1). The lowest motility was observed in the 25% EY group (65 ± 5.0%). Among the LDL groups, 2.5% LDL had significantly lower motility (p < 0.05).
Post-Thaw Sperm Quality of Cryopreserved Bee Semen in Groups

Data are presented as mean ± standard error.
Within a column, means without a common superscript differ (p < 0.05; n:10).
EY, egg yolk; LDL, low-density lipoprotein.
Viability
In fresh semen, viability was determined as 98 ± 2.0% (±SE). The lowest viability was recorded in 25% EY (71.6 ± 3.0% dead sperm rate), and the highest viability was recorded in 25% LDL (7.4 ± 0.8% dead sperm rate) (p < 0.05). No significant differences were found between the 0% LDL and 2.5%, 5%, or 10% LDL groups (p > 0.05).
Membrane integrity
In fresh semen, membrane integrity was measured as 98 ± 4.2% (±SE). The 25% LDL group exhibited the highest integrity (96.1 ± 1.8%), while the 25% EY group had the lowest (70 ± 4.2%) (p < 0.05). Among LDL groups, 2.5% LDL displayed significantly lower integrity (p < 0.05).
Longevity
In fresh semen, the longevity was measured as 120 ± 5.0 (±SE). The longest post-thaw motility duration was observed in the 0% and 5% LDL groups (120.0 ± 6.0 and 120.0 ± 4.0 hours, respectively). The shortest duration occurred in the 25% EY group (72.0 ± 5.0 hours) (p < 0.05).
Fertility rate
Fresh semen fertility was 97.3 ± 3.76% (±SE). Among frozen–thawed groups, 25% LDL had the highest fertility, significantly higher than the 25% EY and 2.5% LDL groups (p < 0.05). The 5% and 10% LDL groups exhibited similar fertility to the 0% LDL group (p > 0.05) (Table 2).
Fertility Results of the Bee Sperm Extender Groups
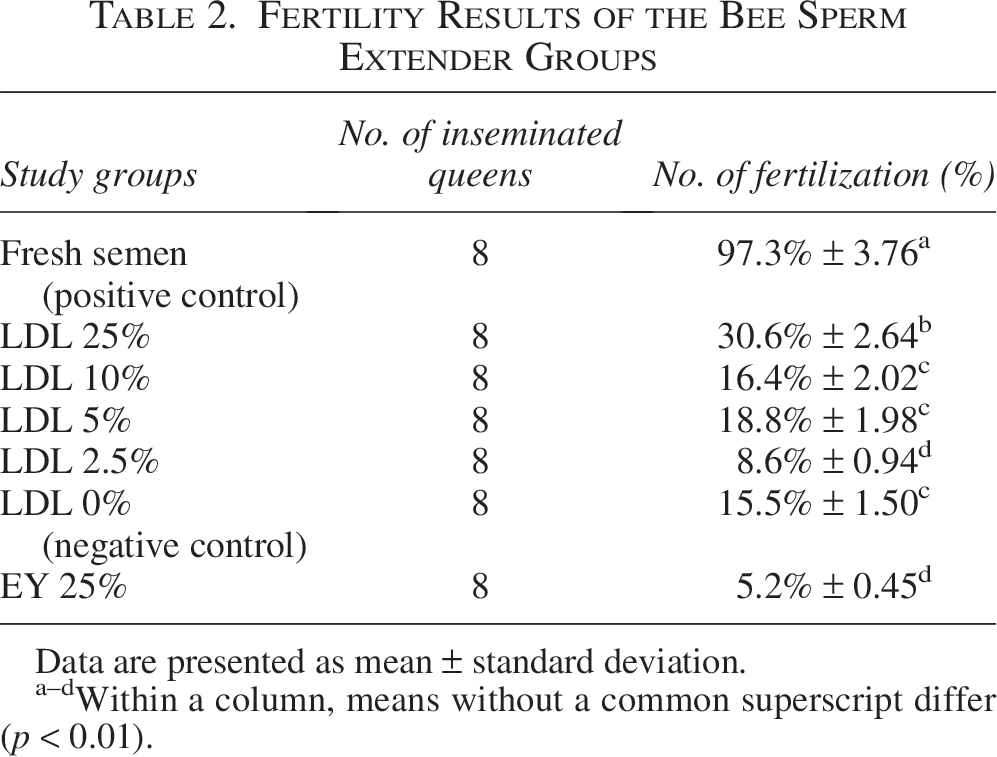
Data are presented as mean ± standard deviation.
Within a column, means without a common superscript differ (p < 0.01).
Discussion
According to the literature reviews, this study is one of the first systematic studies using LDL in the cryopreservation of honeybee sperm. In the experimental process, the effects of diluents prepared using different LDL concentrations (0%, 2.5%, 5%, 10%, and 25%) on post-thaw sperm quality and fertilization success were comprehensively evaluated. The obtained data revealed that sperm parameters were preserved at the highest level, especially in groups containing high levels of LDL (25%), and fertilization capacity increased accordingly.
Freezing and storing reproductive cells for long periods is one of the most important strategies for the sustainability of biodiversity. The indispensable role of honeybees, which are at the center of agricultural production, in the ecosystem has led to the acceleration of genetic improvement studies for this species.29–31 With the development of artificial reproduction techniques as an alternative to natural mating, there is great potential for the preservation of select genotypes and the safe transfer of genes between geographically isolated populations. However, scientific studies on the application of these technologies to honeybees are still limited. Many researchers have focused on new extender compositions and freezing techniques that will preserve both the viability and functionality of bee sperm.6,32
One of the most common biological additives used to protect sperm against cold shock during the freezing–thawing process is EY. EY has the potential to reduce cold-induced structural deterioration by stabilizing sperm membranes thanks to the lipids and proteins it contains.24,33–36 However, it also has some serious disadvantages. This component of animal origin has a direct effect on mitochondrial respiration and is also a potential source of bacterial contamination. In particular, HDL disrupts membrane fluidity by causing cholesterol in the sperm membrane to be transported outward, making sperm more vulnerable to cold shock.11,19,37–39 Another disadvantage is its high content of steroid hormones and calcium. These substances can initiate a premature acrosome reaction in sperm and significantly reduce fertilization potential.19,33,34 For these reasons, in recent years, instead of the whole EY, the use of subfractions, which are thought to be the main source of its protective effect, in the extender composition has been on the agenda.14,40–43
Many scientific studies have shown that positive results are obtained by adding LDL fractions obtained from EY to sperm freezing solutions at certain rates. It has been reported that extender compositions, especially those using LDL rates ranging from 6% to 10%, provide significant improvements in parameters such as sperm motility, acrosome integrity, and plasma membrane protection.13,19,43–45 These findings reveal that LDL forms an effective protective shield against cold shock on frozen sperm cells and reduces loss of viability. In summary, the findings obtained within the scope of this study are largely consistent with the existing literature. In the extender group prepared with high doses of LDL, significant improvements were observed in the determinant parameters of sperm quality, and it was confirmed that the traditionally used EY was weak in terms of these parameters. This situation once again reveals how important it is to carefully select extender contents, especially in species with sensitive sperm structures such as honeybees. The main damage that occurs in the plasma membrane, one of the vital structures of the sperm cell during the cryopreservation process, is mostly due to the removal of cholesterol from the membrane structure. This causes the membrane to lose its flexibility and durability, making the cell more vulnerable to environmental stresses.46,47 Phospholipids play a fundamental role in maintaining this structural stability. In particular, the integrity of the LDL structure largely depends on these phospholipid-containing molecules. 24 The phospholipids contained in the LDL components form a protective layer that directly supports the membrane structure of the sperm cell during cryopreservation, forming a lipid film that acts as a shield on the outer surface of the spermatozoa.11,45,48
In studies where LDL is used in different animal species, it is noteworthy that optimum usage rates vary. This difference is most likely due to the unique biochemical properties of each species. 19 In addition, some lipophilic antioxidants in the LDL composition help reduce oxidative stress on the membrane. 49 These molecules protect both membrane integrity and intracellular order by minimizing membrane damage caused by reactive oxygen species. Many studies have shown that LDL increases sperm motility after freezing and particularly significantly improves resistance to cold shock.13,19,24,34,35,50–53 Similarly, it has been repeatedly proven in the literature that the addition of LDL into the extender offers a much safer and more effective alternative to the traditionally used EY.21,24,43,44,54–60
In our research with fresh semen, the values for motility, viability, membrane integrity, and longevity were recorded as 95 ± 5.0, 98 ± 2.0, 98 ± 4.2, and 120 ± 5.0, respectively. After thawing, the 25% LDL group exhibited the highest motility. The lowest motility was observed in the 25% EY group. Similarly, the viability rate was lowest in the 25% EY group, while the highest viability was in the 25% LDL group. The greatest membrane integrity was found in the 25% LDL group, whereas the lowest was in the 25% EY group. Following thawing, longevity was significantly reduced in the 25% EY group. Dong et al. 61 reported that motility levels were significantly higher in the LDL-added groups compared with the groups containing 20% EY after cryopreservation of monkey sperm. Similarly, a study conducted on goats by Anand et al. 62 reported that LDL played a protective role in terms of sperm kinetics under short-term semen storage conditions and also improved sperm quality during the freezing process. This result shows that LDL has positive effects not only during the cryopreservation stage but also at intermediate stages in the cold chain. On the contrary, in a study conducted on cattle by Ondrej et al. 63 the addition of an LDL additive based on soy lecithin extender resulted in a significant improvement in spermatozoa quality after thawing. This finding suggests that LDL can also exhibit synergistic effects with diluent structures of different bases.
The findings obtained in our current study are generally consistent with the studies mentioned above. Especially in the groups containing 25% LDL, a significant increase was observed in the motility, membrane integrity, and viability rates of frozen–thawed honeybee sperm. On the contrary, in the group containing 25% EY, a clear decrease occurred in these parameters; moreover, the life span of the sperm was significantly reduced. These results are consistent with the general literature trend that the protective effect of LDL is clearly observed, while EY has some negative effects.
Neves et al. 64 reported that the use of low-dose LDL in cryopreservation studies in dogs had a protective effect on sperm cells. In addition, in a study conducted on mice by Yıldız and Erol, 14 it was stated that the addition of 2.5% LDL to the diluent provided significant improvements in spermatological parameters. However, in the same study, it was reported that sperm quality remained at the lowest levels after thawing in the groups where 7.5% and 10% LDL were used. This situation points to some studies suggesting that LDL may be effective at low doses but may disrupt membrane structure at high concentrations. Shahverdi et al. 65 reported that the addition of 4% LDL in the cryopreservation of rooster sperm significantly increased sperm quality. Brito et al. 66 reported successful results in their study on buffalo sperm by adding 2% LDL to the TES-Tris-milk-based diluent. These examples showed that LDL supplementation provides general protective effects on sperm cells in different species. However, it is noteworthy that the most effective results were observed at low LDL levels (2%–4%) in many studies. In this respect, although the high LDL level in our study seems to give the best results, it may be attributable to biochemical differences between species. The lipid composition of the sperm membrane varies from species to species; this may cause the optimal LDL dosage to change. In addition, many factors such as the diluent components used, freezing speeds, thawing protocols, and insemination methods can directly affect the results obtained. Therefore, it is natural that low-dose LDL gives optimum results in some species; however, higher ratios are required in species with more sensitive structures, such as honeybee sperm, and this can be explained by our findings.
Some previous scientific studies have shown that when LDLs are used in high amounts, these molecules tend to interact with each other and aggregate, which reduces the protective functions of LDL.14,24 It has been reported that this aggregation can cause physical accumulation that can damage cell membranes, especially during cryopreservation, and can cause a decrease in sperm quality. Similarly, some studies have reported that when LDL is used in excessive concentrations, sperm plasma membranes are disrupted, resulting in serious decreases in parameters such as motility and vitality after thawing.24,44,67 However, such findings do not coincide with the results obtained in our study because we have reached the highest values in terms of post-thawing sperm motility, vitality, and fertilization success in the extender group containing 25% LDL. This situation can probably be explained by the fact that the extender components used support the biological effects of LDL or that honeybee sperm have different membrane compositions depending on the species.
Numerous studies conducted on various species have shown that LDL-based extender compositions have positive effects on both motility and fertility in frozen–thawed sperm samples.54,62,68–70 These findings support the fact that LDL is an effective cryoprotectant in many organisms. However, the success levels achieved can vary greatly depending on the LDL concentrations used, the composition of the extender, the freezing and thawing protocols, and even the insemination techniques. On the contrary, traditional diluent additives such as EY, despite being used in extender composition for a long time, are also known to carry some important risks. Providing an open environment, especially against microorganisms, increases the risk of bacterial or viral contamination, which becomes an important factor limiting the fertilization success of the sperm used in artificial insemination.37,71
The fertility rate was highest with fresh semen. When inseminations using frozen semen were examined, this rate was greatest in the group prepared with 25% LDL. The groups with the lowest fertilization success were the 25% EY and low LDL-added (2.5%) groups. The findings showed that the addition of 25% LDL significantly increased both sperm quality and fertilization success. Shahverdi et al. 65 found that 4% LDL provided the highest fertilization rate in frozen–thawed rooster sperm samples. In our study, much higher fertilization rates were obtained in bee sperm with 25% LDL addition. This confirms once again that LDL requires different optimum levels in different species. In freezing studies conducted with sperm obtained from honeybee drones, it was observed that the egg-laying efficiency of queen bees decreased significantly when whole EY was used in the diluent. This situation is due to the risk of particles in the EY content blocking the sperm–ovum and may cause infections in inseminated queens and directly negatively affect fertilization success.23,72 This finding is exactly the same as the observation of both low motility and low fertility rates in the group where 25% EY was used in our study. Especially, the different success rates of LDL in different animal species are largely attributed to the differences in the biochemical content of the sperm membrane. Factors such as phospholipids, cholesterol ratios, membrane proteins, and antioxidant enzyme systems in the membrane structure can directly affect the level of effect of LDL.61,73 Therefore, the ideal dosage of LDL should be determined separately for each species.
El-Sharawy et al. 74 concluded that 12% LDL in the cryopreservation of buffalo spermatozoa significantly increased both cryopreservation success and fertilization rates. Similarly, in another study conducted on Nili-Ravi bull spermatozoa, it was reported that extenders prepared with 10% LDL provided high fertility in vivo. 75 These results indicate that LDLs have a biofunctional role that not only contributes to membrane stabilization but also directly increases fertilization capacity. El-Siefy et al. 76 reported that rabbit sperm cells responded better to both the freezing process and the subsequent fertilization process when 10% LDL isolated from EY was added to the extender. However, in our study, the highest fertilization rate among the LDL-added groups was obtained in the diluent containing 25% LDL. This indicated that higher LDL levels may also positively affect sperm quality in some species and that the optimal dose may differ depending on the species. The technique used in inseminations, injection volume, number of spermatozoa, environmental conditions, depth of semen deposit, and frequency of insemination are all important variables that can directly affect fertilization rates.77–79
Consuegra et al. 80 reported in their vitrification study on horse sperm that even the addition of only 0.25% LDL to the vitrification medium significantly improved sperm quality. This result suggests that LDL may have a membrane-protective effect even at low levels. It has been reported that the best results in cryopreservation of bovine sperm were obtained with 8% LDL and in dog sperm with 6% LDL contribution. 19 Pytlik et al. 81 reported that the post-thaw quality of bull spermatozoa was positively affected by LDL supplementation. Snoeck et al. 82 in rams showed that 8% LDL resulted in an improvement in post-thaw velocity parameters. Perumal et al. 67 reported that high sperm quality was achieved in Mithun cattle spermatozoa with 8% and 10% LDL supplementation during liquid storage. Alvarez-Rodriguez et al. 49 on brown bear spermatozoa showed that LDL-based extenders have higher antioxidant activity and cell viability. A number of researchers have argued that LDL can protect the sperm plasma membrane against various external stress factors during cryopreservation and maintain the membrane permeability, thus making the cell resistant to oxidative damage.83,84 The findings support that LDL helps stabilize sperm membrane structure, especially providing protection against lipid peroxidation.44,85–88
The interphase film formed by LDL during freezing and thawing processes protects membrane integrity by preventing the breakdown of phospholipid structures during cryo-damage and can compensate for the lost lipid structures. In addition, thanks to the ability of LDL to bind some proteins found in seminal plasma that can damage the cell, the interaction of these harmful components with sperm can be reduced.13,34,44,89,90 The variety of LDL dosages used for semen cryopreservation in different animal species is most likely due to differences in the biochemical composition of the sperm plasma membrane. 21 These differences are related to many factors, such as phospholipid ratio, cholesterol content, structure of membrane proteins, and the effectiveness of antioxidant systems. Therefore, the relationship between cryopreservation success and LDL dosage depends not only on the amount but also on the physiological and biochemical properties of the targeted species.
Conclusions
The findings obtained within the scope of this study clearly demonstrated that it is possible to store bee sperm for long periods by freezing and that high fertility values can be achieved after thawing. The effectiveness of cryopreservation is measured not only by the freezing of the sperm cell but also by its ability to maintain its biological functions after thawing. In this context, the importance of a correctly formulated extender and an optimized freeze–thaw protocol was emphasized once again. In our study, the effects of diluents containing EY and LDL in different proportions were evaluated in detail. In line with the obtained data, it was determined that the extender group with 25% EY added gave significantly lower results in basic parameters such as sperm viability, membrane integrity, and motility after thawing. This supports the fact that the biological components contained in EY can have toxic effects on sperm in some cases and even cause damage to the membrane structure. On the contrary, in the group with 25% LDL addition, it was determined that the membrane integrity of the sperm was highly preserved after thawing, motility was higher compared to all other groups, and most importantly, fertilization rates reached the highest levels in queen bees obtained through artificial insemination. These findings show that the cryoprotective effects of LDL are not limited to membrane stabilization but also play an active role in the continuity of cellular functions. In addition, the results of the study confirm that LDL can be used as an effective alternative to EY in cryopreservation applications and even outperform it in some cases. Although the ideal LDL dose varies for each organism due to biochemical differences between species, it was concluded that the use of high levels of LDL in sensitive species such as honeybees can increase extender effectiveness.
Authors’ Contributions
N.C.: Writing—review and editing, visualization, validation, investigation, and data curation. A.G.: Visualization and methodology. C.Y.: Formal analysis, data curation, and supervision. O.K.Y.: Data curation and methodology. All authors have reviewed and implemented the article before submission.