
Abstract
Introduction:
The development of ice crystals during the freezing process can be detrimental to the viability and fertilization capacity of frozen–thawed spermatozoa. The unique properties of antifreeze proteins allow them to inhibit the formation of ice crystals during cell cryopreservation.
Objective:
Previous studies have assessed the general sperm quality parameters, reactive oxygen species (ROS) level, lipid peroxidation, and sperm apoptosis after using antifreeze protein III (AFP III) for cryopreservation. However, the data regarding changes in protein expression and their relation to sperm quality after thawing are still lacking. Therefore, this work addresses associated proteomic changes in post-thawed dog sperm.
Methods:
Two experiments were conducted using high (Experiment I) and low (Experiment II) concentrations of AFP III. Semen samples from four dogs were divided into aliquots and diluted with Tris-egg yolk extender supplemented with 0 (control), 1, 5, 10, or 15 µg/mL AFP III (Experiment I) or 0, 1, 2, 3, or 4 µg/mL AFP III (Experiment II) based on a previous literature review. After being frozen in LN2 and thawed, sperm motility parameters, viability, acrosome integrity, and apoptosis were evaluated. Furthermore, the AKAP4, ATP1B1, and HSP70 proteins, which are associated with good sperm quality and freezability, were subjected to sodium dodecyl sulfate–polyacrylamide gel electrophoresis and Western blotting to assess their expression and levels.
Results:
In the high-concentration experiment, AFP III at 1 µg/mL significantly increased (p < 0.05) total and moderate-progressive motility compared with the control and other AFP III concentrations. Moreover, the 1 µg/mL group had higher HSP70 and AKAP4 protein levels than the control and other concentration groups, though the differences were not significant.
Conclusion:
These findings suggest that adding 1 µg/mL AFP III to the semen extender enhances the motility of post-thawed dog sperm and suggest the potential use of HSP70 and AKAP4 proteins as biomarkers for good semen quality.
Introduction
Cryopreservation of spermatozoa is a crucial technique because it promotes genetic selection and allows preservation of cells of rare breeds in cryobanks. 1 The use of cryopreserved sperm, particularly in dogs, can support future breeding projects and reduce the costs of maintaining and transporting live dogs. 2 Despite these advantages, freezing temperatures can have negative effects such as ice crystal formation and high osmotic pressure, which can compromise the lifespan of frozen–thawed spermatozoa, and unlike other domestic animals, dog sperm react differently to cryopreservation, often with low post-thawed conception rates.3,4 Such effects can be particularly severe due to disruption of sperm cell cytoplasm, cytoskeleton, and genome structure.5–7 In addition, cooling slows cellular activity, changes protein–protein interactions, and alters membrane fluidity. 8 Over time, every crucial step in the cryopreservation of semen, including cooling and freezing duration and composition of semen extender, has undergone changes and enhancements to improve freezability and cell survival rates and increase the post-thawed characteristics of dog spermatozoa.1,9
Recent studies have explored the use of various antioxidants, including astaxanthin, resveratrol, quercetin, curcumin, myo-inositol, and olive-derived compounds, to reduce oxidative stress and improve dog sperm quality. Mesenchymal stem cells and their derivatives also provide protective effects, while low-density lipoproteins serve as alternatives to egg yolk in extenders to reduce biosecurity risks associated with animal-origin components. 4
Antifreeze proteins (AFPs) have recently come into use as cryoprotectants due to their ability to preserve cell membranes and inhibit ice crystal formation during the freezing of living cells.5,10 AFPs, which are naturally present in cold-adapted bacteria, algae, nematodes, fish, insects, and plants, remain functional at temperatures below freezing.11,12 Based on their origins and structural characteristics, AFPs are classified into five groups (antifreeze glycoproteins and types I–IV). 13 AFP type III, derived from eelpout and northern or Antarctic zoarcid fish, possesses a three-dimensional structure resembling a triangular prism with a large hydrophobic core and two moderately flat, nonpolar surfaces that bind to ice and inhibit its growth through an adsorption–inhibition mechanism.14–16 According to earlier studies, addition of AFP III reduced freeze–thaw damage to sperm of various species.17–34 The efficiency of AFP varies by species, AFP concentration, and cryopreservation protocol. 35
To the best of our knowledge, no previous studies have investigated protein expression levels after adding AFP III to the freezing extender during dog sperm cryopreservation. Moreover, although high AFP III concentrations were used with a positive effect in chimpanzees, 18 only low concentrations of AFP III have been tested in dogs.33,34 Therefore, the goal of this study was to evaluate the protective effects of AFP III at low and high concentrations in freezing extender on post-thawed sperm characteristics (motility parameters, viability, acrosome integrity, apoptosis) and the expression levels of A-kinase anchoring protein 4 (AKAP4), ATPase Na+/K+ transporting subunit beta 1 (ATP1B1), and heat shock protein 70 (HSP70). These proteins were selected as biomarkers to assess sperm freezability because they are associated with high sperm quality: AKAP4 for motility, hypermotility, capacitation, and fertility36,37; ATP1B1 for sperm freezability 38 ; and HSP70 for viability, motility, and fertility.39–41
Materials and Methods
Unless otherwise stated, all chemicals were purchased from Sigma-Aldrich (St. Louis, MO, USA).
Animals
Semen samples were collected from four trained 2- to 5-year-old dogs, two Pomeranians and two Beagles. All experimental dogs were housed in isolated pens and provided daily commercial dog food (BioMill®, Jindo, Republic of Korea) and fresh drinking water ad libitum. The dogs used in this study were treated and received care under the Guiding Principles for the Care and Use of Research Animals, as established by Jeonbuk National University (NON, 2023-019).
Semen collection and preparation
Sixty-four samples were obtained from 4 dogs in 16 sessions (one ejaculate from each dog per session) by digital manipulation. The second fraction (sperm-rich) of each ejaculate was collected using pre-warmed (37°C) calibrated plastic tubes (SPL Life Sciences Co., Gyeonggi, Republic of Korea) connected to a funnel. The samples were immediately transferred to the laboratory of Theriogenology and Reproductive Biotechnology in a Thermos flask at 37°C. Only normal ejaculates with sperm concentrations ≥2 × 108 sperm/mL, motility ≥70%, and normal morphology ≥80% were used in this study. Each ejaculate from the four dogs was centrifuged (12 minutes at 300g), and the pellets were pooled. To calculate the sperm concentration, 10 µL of the sperm pellet was placed in a 1.8 mL microcentrifuge tube containing 990 µL of water and mixed by inverting slowly before 10 µL was aliquoted into a Neubauer chamber (Marienfeld, Lauda-Königshofen, Germany). The chamber was positioned on a phase contrast microscope (Zeiss, Oberkochen, Germany), and 25 squares were analyzed at ×400 magnification. The total sperm concentration is expressed in millions per mL.
Sperm freezing extenders and freezing procedure
Sperm dilution was performed in two steps. First, extender 1 (Tris 3.025 g, citric acid 1.7 g, fructose 1.25 g, Na benzylpenicillin 0.06 g, streptomycin sulfate 0.1 g, and 20 mL egg yolk per 100 mL ultrapure water [ProGen Life Sciences, NY, United States]) was added to each of the experimental samples to a final concentration of 200 × 106 cells/mL. The extended sperm samples were placed at 4°C for 1 hour for equilibration. Subsequently, extender 2, comprised of extender 1 and glycerol (4% final concentration), was added to each sample 1:1 (v/v). Both extender sample types 1 (osmolarity, 390) and 2 (osmolarity, 1578) were supplemented with 0 (control), 1, 5, 10, or 15 µg/mL of AFP III (A/F Protein Inc., Waltham, MA, USA) as a final concentration in experiment 1 or with 0 (control), 1, 2, 3 or 4 µg/mL of AFP III as a final concentration in experiment 2. Samples were maintained at 4°C for 30 minutes (equilibration time), manually loaded into 0.5 mL IMV French straws (IMV Technologies, Saint Quen-Surlton, France), and sealed with an impulse bag sealer NT-300 (Hanato Corporation LTD, Gimcheon City, Republic of Korea).
The straws (100 × 106 cells/mL final concentration) were frozen in a Styrofoam box (29.5 × 18.7 × 24 cm3) that contained liquid nitrogen (LN2) to a height of 5 cm. The straws were placed 7 cm above the LN2 surface for 20 minutes and then were immersed in LN2 until use. The cooling rate for the present study was similar to that used by Nabeel et al. 42 as follows: 30.6°C/min from 5.5°C to −15.0°C, 88.2°C/min from −15.0°C to −60°C, and 14.7°C/min from −60°C to −116°C.
Post-thawed sperm evaluation
The frozen straws were thawed in a water bath at 37°C for 25 seconds. Then, the thawed samples were placed into 1.8 mL microcentrifuge tubes. The sperm quality variables were analyzed as described below.
Sperm motility parameters
Motility and kinematic parameters were measured by a computer-assisted sperm analysis system using Sperm Class Analyzer version 6.3.0.32 software (Microptic S.L., Barcelona, Spain). A phase contrast microscope (Nikon Eclipse E-200; Nikon Corporation, Kanagawa, Japan) equipped with a Basler camera (acA1300-200uc, Ahrensburg, Germany) and an attached heating stage set at 37°C (Tokai Hit Co., Ltd., Shizuoka-ken, Japan) was used in this study. A 2.5 µL aliquot of thawed spermatozoa was placed into a preheated (37°C) Leja standard-count 8-chamber slide with a 20 μm (Leja Products B.V., Nieuw-Vennep, Netherlands). For each sperm sample, five videos lasting three seconds each were recorded in different fields.
The following sperm motility parameters were analyzed: total motility (%), progressive motility (%), average path velocity (VAP, μm/s), curvilinear velocity (μm/s), straight line velocity (μm/s), linearity (%), straightness (STR, %), wobble (%), amplitude of the lateral head displacement (μm), and beat cross frequency (Hz). As described by Dorado et al., 43 with minor modifications, the manufacturer’s recommended software settings were adjusted as follows to identify individual spermatozoa: grid distance, 100 μm; frame rate, 25 frames/s; number of images, 25; depth of sample chamber, 20 μm; volume per chamber, 2.5 µL; temperature of analysis, 38°C; area (min), 10 μm2; area (max), 80 μm2; static, 10 μm/s; slow-medium, 25 μm/s; rapid, > 45 μm/s; progressive, STR > 45%; connectivity, 14 pixels; VAP points, 5 pixels; number of sperm analyzed, 500; number of fields analyzed, 5.
Sperm viability
A sperm viability kit (Thermo Fisher Scientific, Eugene, OR, USA) was used following the procedure of Yu. 44 Briefly, SYBR-14 (5 µL, 100 nM) was added to 50 µL of frozen–thawed spermatozoa, and the mixture was incubated for 5 minutes at room temperature in the dark; 5 µL of propidium iodide (PI, 12 μM) was then added, and the mixture was incubated for an additional 5 minutes. For each sample, at least 500 spermatozoa were assessed using a fluorescence microscope (Axio, Carl Zeiss, Göttingen, Germany). Sperm cells labeled with a green stain were alive with an intact membrane, and those stained red were dead with damaged membranes.
Acrosome integrity
Acrosome integrity was measured with Fluorescein Isothiocyanate-conjugated lectin from Pisum sativum (FITC-PSA) using the procedure described by Yu with minor modifications. 44 After thawing, samples were diluted 1:1 with tris extender (without egg yolk). Smears were prepared from the samples, air dried, fixed in absolute methanol, stained with 30 µL of FITC-PSA (100 mg/mL) solution, covered with parafilm, incubated for 20 minutes in darkness, placed in a container of distilled water for 15 minutes, and air dried. The slides were observed using an epifluorescence microscope (Axio, Carl Zeiss, Göttingen, Germany). At least 500 sperm cells were counted for each sample. The presence of green acrosomal fluorescence was considered indicative of an intact acrosome, and partial or total absence of fluorescence indicated acrosomal membrane damage.
Phosphatidylserine translocation
An annexin V-FITC apoptosis detection kit I (BD Pharmingen, San Diego, CA, USA) was used to evaluate the phosphatidylserine (PS) translocation index (apoptosis), following the procedure described by Yu. 44 Briefly, frozen–thawed sperm were centrifuged at 300g for 5 minutes, and the supernatant was discarded. The sperm pellet was washed twice using 1× Dulbecco’s phosphate-buffered saline (DPBS; 14190, Gibco, Grand Island, NY, USA) and resuspended (1 × 106 sperm/mL) in 1× annexin binding buffer at room temperature. Aliquots (100 µL of 1 × 105 cells) of the sperm suspension were transferred to 5 mL tubes. For proper cell gating, Annexin-only, PI-only, and unstained controls were analyzed. Annexin V-FITC (5 µL) and 5 µL of PI were added to the samples. The tubes were gently mixed and incubated at room temperature for 15 minutes in the dark. After incubation, additional 1× binding buffer (400 µL) was added to each tube.
All fluorescence signals from the labeled spermatozoa were analyzed with a BD AccuriTM C6 plus flow cytometer (BD Biosciences, NJ, USA) equipped with a 15-mW air-cooled 488-nm argon-ion laser. FL1 (Annexin V) signals were detected through a 530/30 nm bandpass filter, and FL2 (PI) signals were detected through a 585/42 nm bandpass filter, and compensation between FL1 (Annexin V) and FL2 (PI) channels was applied to correct for spectral overlap. Ten thousand sperm cell events were recorded in list mode at a flow rate <200 events/s and analyzed using BD AccuriTM CFlow software. The sperm population was gated based on the log forward and side-scatter properties of the sperm, and flow cytometric analysis was conducted within 1 hour. We defined the ratio between PS-translocated (Annexin +/PI−) sperm and total living (PI−) sperm as the PS translocation index.
Protein analysis
Total protein extraction
Protein extraction from frozen and thawed spermatozoa was performed using a total protein extraction kit (TPETM, G-Biosciences, St. Louis, MO, USA) according to the manufacturer’s instructions, with modifications. In brief, frozen–thawed sperm were centrifuged at 300g for 5 minutes, and the supernatant was removed. The sperm pellet was washed twice using 1× Dulbecco’s phosphate-buffered saline (14190, Gibco) and resuspended in 100 µL of Tris-phosphate-EDTA buffer (TPE Buffer-I). The tube was vortexed for 1–2 minutes and then placed on ice. Then, 60 µL of TPE Buffer-II was added to the mixture, which was vortexed for 30 seconds and placed in a water bath (100°C) for 30 seconds. Another two cycles of boiling and vortexing for 30 seconds were conducted. The samples were incubated in a water bath (100°C) for 10 minutes and centrifuged for 5 minutes at 15,000g at 4°C. The supernatant was transferred to a clean Eppendorf tube and stored at −20°C. Typically, 60 µL of supernatant was recovered following extraction. The protein extracts were used directly for quantification and downstream analyses without any additional concentration or precipitation steps. A 25 µL was used to determine the protein concentration using PiercetTM BCA protein assay kits (Thermo Scientific™, Rockford, IL, USA) according to the manufacturer’s instructions. The total protein yield was 2.04 ± 0.54 mg/mL on average.
Western blot analysis
A total of 50 µg proteins were separated by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis for 80 minutes at 300 V and 100 mA with constant current. A prestained protein ladder (PageRuler™, Thermo Scientific, Rockford, IL, USA) was used as a molecular weight marker. Next, the proteins were transferred onto polyvinylidene fluoride membranes (Immun-Blot® PVDF Membrane, Bio-Rad Laboratories, CA, USA) at 150 amp, 300 V, and constant current for 2 hours. Each membrane was stained with 10 mL of commercial Ponceau stain solution (ProNATM Ponceau S Solution, TransLab, Gunpo-si, Gyeonggi-do, Republic of Korea) for 3 minutes at room temperature, rinsed with distilled water to remove the background stain, and then imaged. The membranes were destained with 1× tris-buffered saline (TBS, 50 mM tris, 150 mM NaCl, pH 7.5) solution containing 0.05% Tween 20 (TBST).
Afterward, the membranes were blocked with 5% nonfat milk in 1× TBS at room temperature for 2 hours. Then, they were incubated separately overnight at 4°C with anti-AKAP4 monoclonal antibody (1:10,000, 4biox, SPQI Company, Lille, France), ATP1B1 monoclonal antibody (1:10,000, Thermo Scientific, Invitrogen, Rockford, IL, USA), and HSP70 monoclonal antibody (1:1000; Thermo Scientific, Invitrogen). After washing three times with 1× TBST (10 min each), the membranes were incubated with goat antimouse IgG (H + L), SuperclonalTM recombinant secondary antibody with HRP (1:10,000; Thermo Scientific, Invitrogen) at room temperature for 2 hours. The membranes were washed three times with 1× TBST (10 minutes each) and stained with a western blotting detection kit (WesternBrightTM Sirius, Advansta, San Jose, CA, USA) to analyze immunoreactive proteins. Images were acquired using a ChemiDoc XRS+ gel imaging system (Molecular Imager®, Bio-Rad, Hercules, CA, USA) and analyzed using Image Lab 6.0 (Bio-Rad). As a loading control, the same membranes were incubated with a mouse monoclonal antibody against β-actin (1:10,000; CUSABIO®, Houston, TX, USA).
Experimental design
Experiment 1: Effects of high AFP III concentrations on post-thawed dog sperm motility, viability, acrosome integrity, apoptosis, and protein expression levels
This experiment investigated the effects of high AFP III concentrations on the cryopreservation of dog spermatozoa after freezing and thawing. Spermatozoa were extended in Tris-egg yolk-glycerol (TEY) supplemented with 0 (control), 1, 5, 10, or 15 µg/mL AFP III (A/F Protein Inc., Waltham, MA, USA), cooled, frozen, and thawed as described above. The motility, viability, acrosome integrity, apoptosis, and relative expressions of AKAP4, ATP1B1, and HSP70 proteins of the dog spermatozoa were analyzed after thawing.
Experiment 2: Effects of low AFP III concentrations on post-thawed dog sperm motility, viability, acrosome integrity, apoptosis, and protein expression levels
This experiment investigated the effects of low AFP III concentrations on the cryopreservation of dog spermatozoa after freezing and thawing. Spermatozoa were extended in TEY supplemented with 0 (control), 1, 2, 3, or 4 µg/mL AFP III (A/F Protein Inc., Waltham, MA, USA), cooled, frozen, and thawed as described above. The motility, viability, acrosome integrity, apoptosis, and relative expressions of AKAP4, ATP1B1, and HSP70 proteins of the dog spermatozoa were analyzed after thawing.
Statistical analysis
For each experiment, four replicates were prepared from pooled samples on different days, and the mean was calculated to increase the validity of the experimental data and results. The effects of AFP III in the extender on the post-thawed quality of dog sperm were analyzed using the Statistical Package for the Social Sciences (IBM SPSS Statistics 26.0, Armonk, NY, USA). All results are presented as mean ± standard error of the mean (SEM) and were assessed with one-way analysis of variance (ANOVA) and Tukey’s statistical tests. p < 0.05 was considered significant.
Results
Experiment 1: Effects of high AFP III concentrations on post-thawed dog sperm motility, viability, acrosome integrity, apoptosis, and protein expression levels
A 1 µg/mL concentration of AFP resulted in significantly (p = 0.0003) higher total sperm motility than the control and other AFP III concentrations (p value for 5, 10, 15 µg/mL were p = 0.0004, p < 0.0001, and p < 0.0001, respectively), and motility with the highest concentration (15 µg/mL) was significantly lower than that with the other concentrations (p value for 0, 1, 5, 10 µg/mL were p = 0.0009, p < 0.0001, p = 0.0007, and p = 0.0212, respectively), (Fig. 1). Except for 15 µg/mL (p = 0.0481), the AFP III-treated groups and the control did not differ significantly in sperm progressive motility (p value for 0 µg/mL vs. 1, 5, 10 µg/mL were p = 0.8190, p = 0.9982, and p = 0.9996, respectively), (Fig. 1). Additionally, Figure 1 shows that the percentage of immotile sperm in the 1 µg/mL group was significantly lower than in the other groups, and the percentage of immotile sperm in the 15 µg/mL group was significantly higher than in the control and other AFP III concentration groups. The effects of AFP III supplementation on the progressive motility types in dog sperm are presented in Supplementary Table S1. The percentage of moderate-progressive sperm after freezing and thawing was significantly higher in the 1 µg/mL group than in the other concentrations and the control conditions. Also, the nonprogressive sperm percentage was significantly higher at 1 µg/mL than at 15 µg/mL. The AFP III-treated groups showed no significant differences from the control group in the motility kinematic parameters (Table 1).
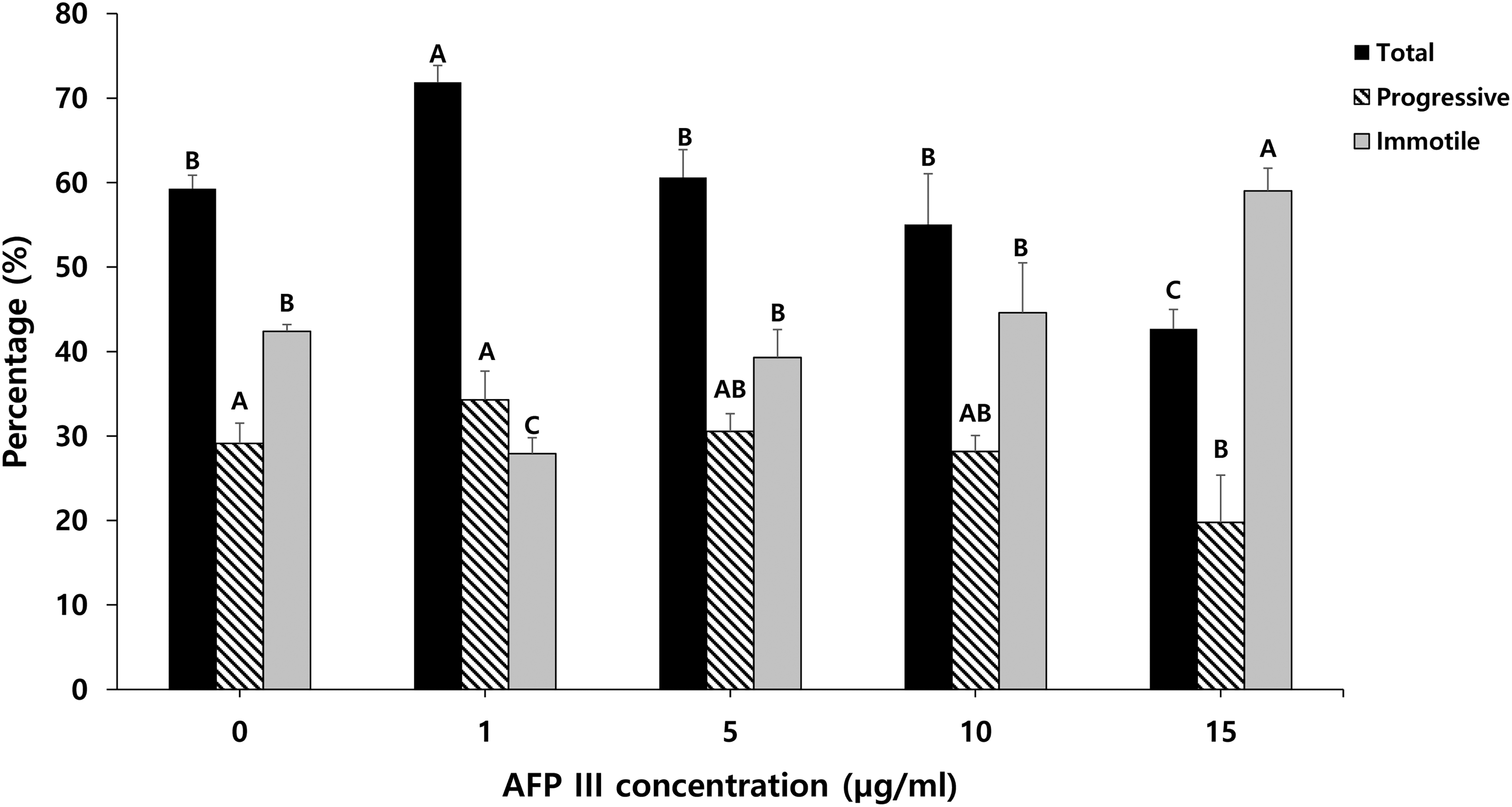
The effect of AFP III supplementation on dog sperm total, progressive motility, and immotile sperm after freezing and thawing. Values are expressed as mean ± standard error (SE). A, B, C: Different letters indicate differences between groups (p < 0.05). Sample size (n = 4 replicates). AFP III, antifreeze protein III.
The Effect of AFP III Supplementation on Dog Sperm Motility Kinematic Parameters After Freezing and Thawing
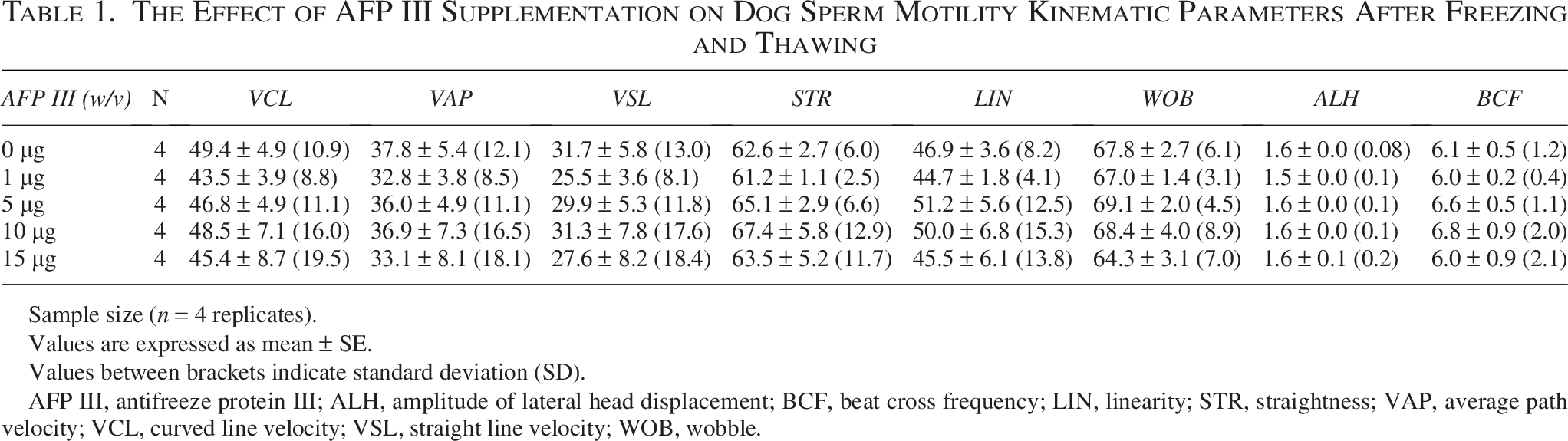
Sample size (n = 4 replicates).
Values are expressed as mean ± SE.
Values between brackets indicate standard deviation (SD).
AFP III, antifreeze protein III; ALH, amplitude of lateral head displacement; BCF, beat cross frequency; LIN, linearity; STR, straightness; VAP, average path velocity; VCL, curved line velocity; VSL, straight line velocity; WOB, wobble.
Sperm viability did not differ significantly among the concentrations of AFP III and the control (Fig. 2). Higher acrosome integrity percentages, but without significance, were observed in the AFP III groups compared with the control group. As demonstrated in Figure 3A and B, the addition of AFP III had no significant effect on the sperm PS translocation index.
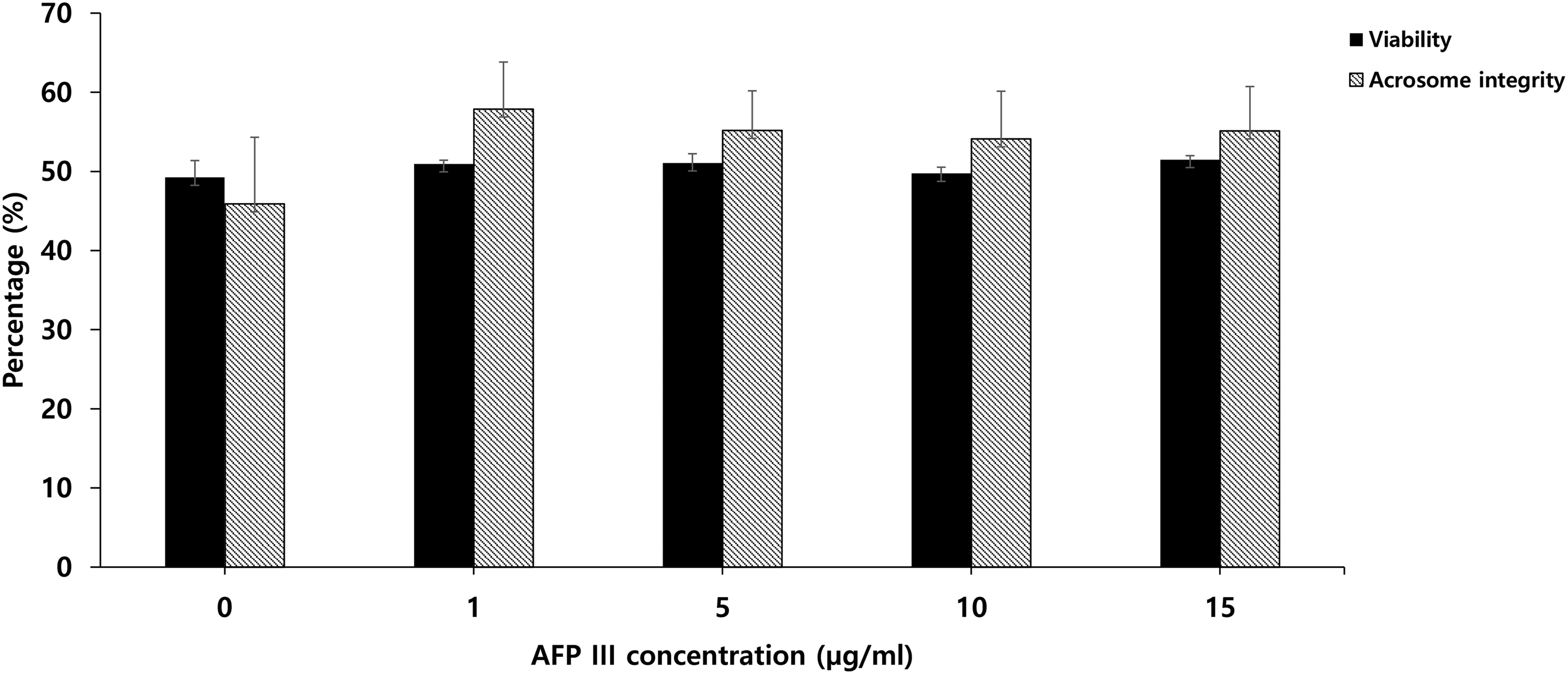
The effect of AFP III supplementation on dog sperm viability and acrosome integrity after freezing and thawing. Values are expressed as mean ± SE. Sample size (n = 4 replicates).
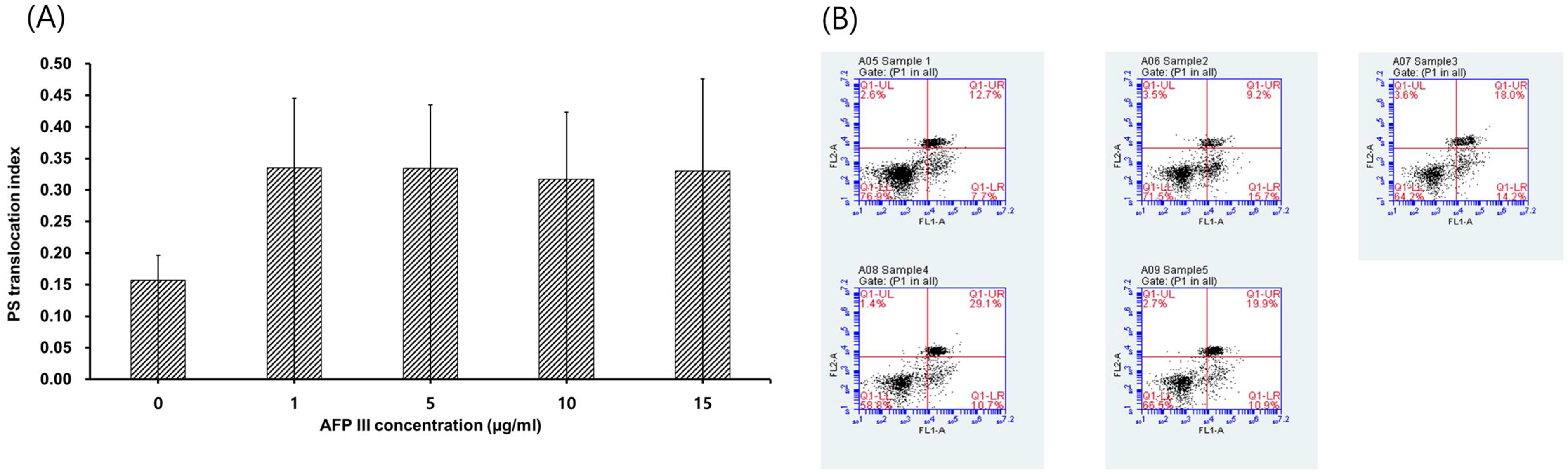
The effect of AFP III supplementation on dog sperm PS (phosphatidylserine) translocation index after freezing and thawing:
The results of western blotting analysis are presented in Figure 4A, B, and C. There was a numerical but not statistically significant increase in HSP70 and AKAP4 protein expression in the 1 µg/mL group compared with the control and other concentrations (Fig. 4B and C), and the same concentration showed lower ATP1B1 protein expression than in the other groups (Fig. 4A). Bands representing 82 kDa for AKAP4, 70 kDa for HSP70, and approximately 38 kDa for ATP1B1 were detected in lysates from the sperm (Fig. 4D).
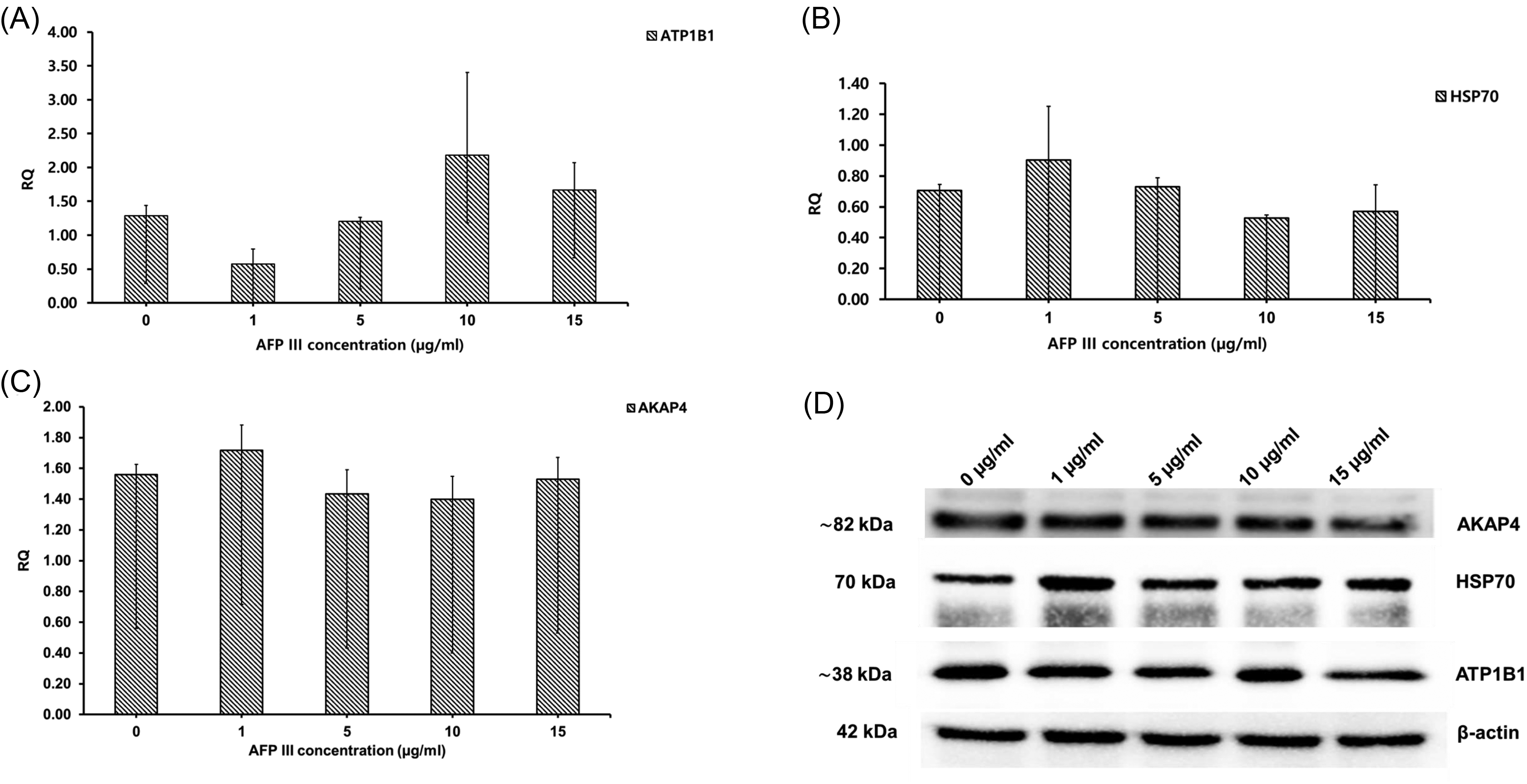
The effect of AFP III supplementation on dog sperm protein expression after freezing and thawing. Values are expressed as mean ± SE.
Experiment 2: Effects of low AFP III concentrations on post-thawed dog sperm motility, viability, acrosome integrity, apoptosis, and protein expression levels
As shown in Table 2, viability did not differ significantly among the experimental groups. In addition, total motility showed no significant differences, although the AFP III groups were higher in value than the control (Table 2). The AFP III concentrations of 1, 2, and 3 µg/mL showed higher progressive motility than the control(p = 0.9964, p = 0.8977, and p = 0.8845, respectively), and 4 µg/mL groups (p = 0.9985, p = 0.9214, and p = 0.9100, respectively), but the differences were not significant (Table 2). Moreover, the AFP III groups showed no significant differences from the control group in motility kinematic parameters (Table 3). The western blotting examination revealed no significant differences among the groups in AKAP4, ATP1B1, or HSP70 protein expression (Fig. 5A, B, and C). Similar to experiment 1, the 1 µg/mL group showed lower ATP1B1 protein expression than the other groups (p = 0.7314, p = 0.3547, p = 0.9993, and p = 0.9820, for 0, 2, 3, and 4 µg/mL, respectively). Bands representing 82 kDa for AKAP4, 70 kDa for HSP70, and approximately 38 kDa for ATP1B1 were detected in lysates from the sperm (Fig. 5D). A heatmap highlighting the main differences in motility and protein expression across the different AFP III concentrations is provided in Supplementary Figure S1.
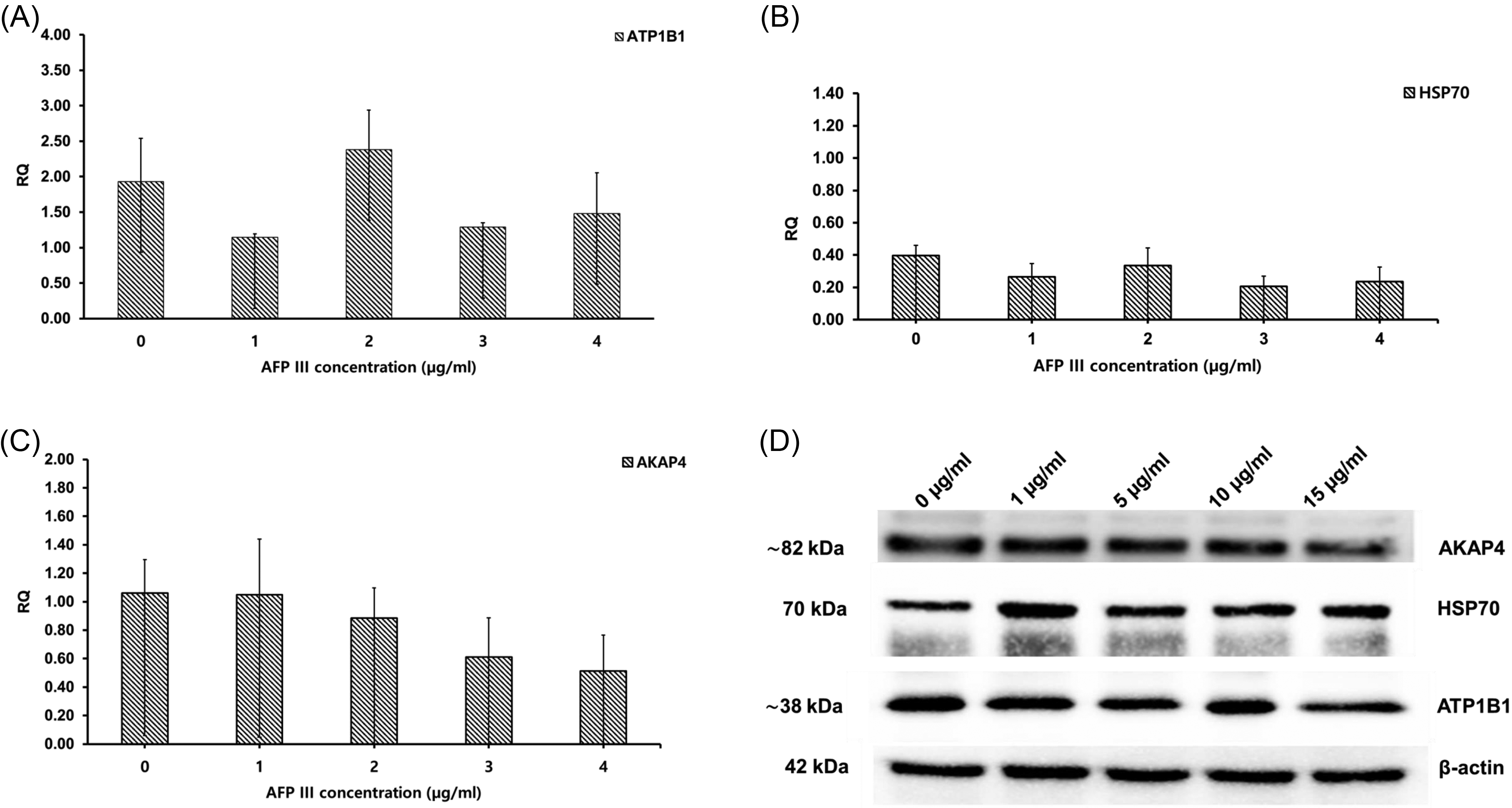
The effect of AFP III supplementation on dog sperm protein expression after freezing and thawing. Values are expressed as mean ± SE.
The Effect of AFP III Supplementation on Dog Sperm Viability, and Motility Types After Freezing and Thawing
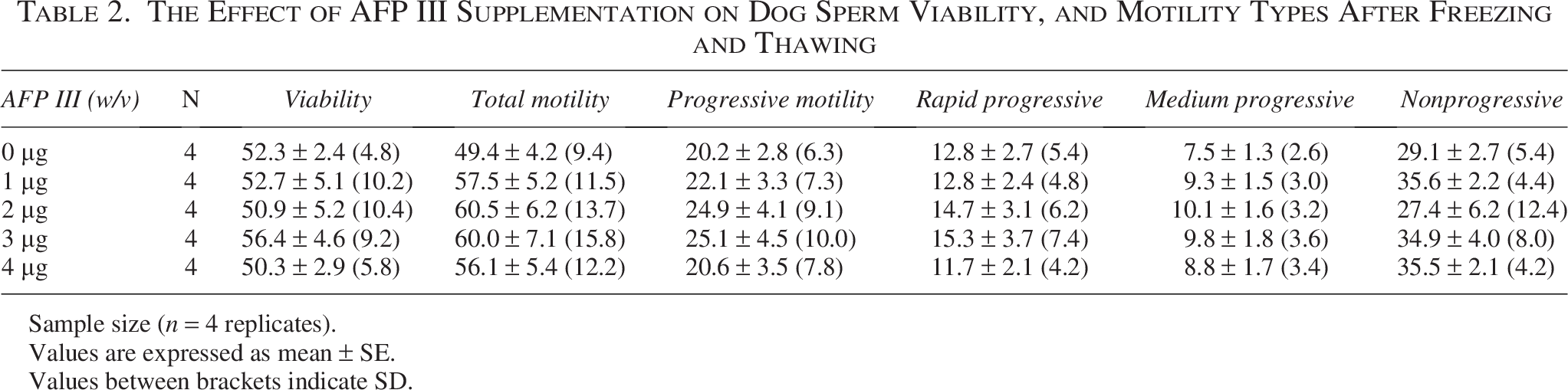
Sample size (n = 4 replicates).
Values are expressed as mean ± SE.
Values between brackets indicate SD.
The Effect of AFP III Supplementation on Dog Sperm Kinematic Parameters of Motility After Freezing and Thawing
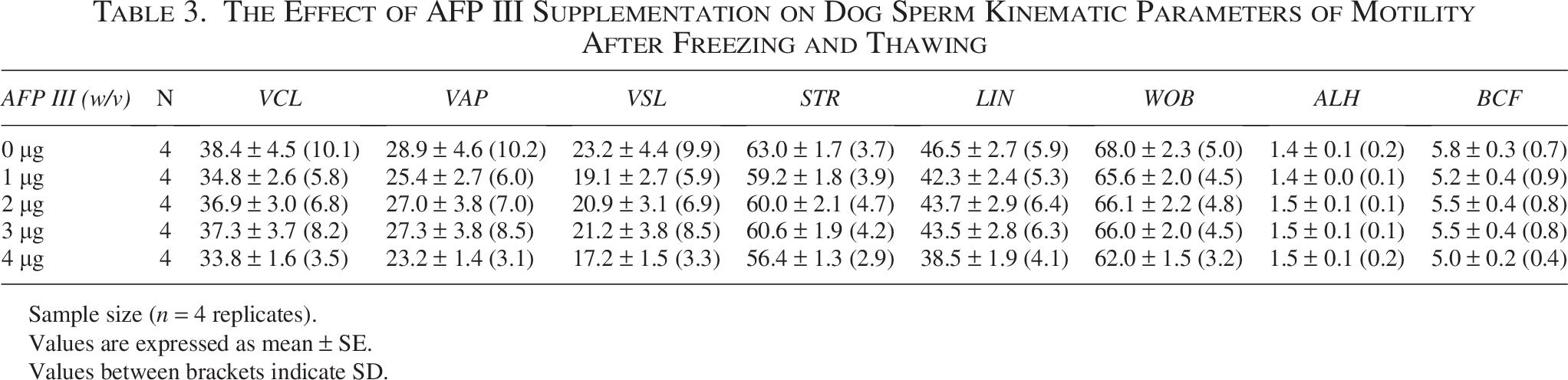
Sample size (n = 4 replicates).
Values are expressed as mean ± SE.
Values between brackets indicate SD.
Discussion
The characteristics and mechanisms of action of AFPs have gradually been elucidated, leading to a growing variety of applications. Currently, AFPs are used in cryomedicine, agriculture, the food industry, environmental management, and anti-icing materials. 45 At low temperatures, these proteins induce thermal hysteresis, inhibit ice crystal growth by binding ice nuclei, alter the ice crystal formation process, and interact with the plasma membrane.46–49
To our knowledge, this is the first study investigating the effects of AFP III in the freezing extender on protein expression levels in frozen–thawed dog spermatozoa. The concentrations of AFP III tested in this study were selected based on a literature review and considering the variation in cryopreservation results attributed to differing AFP concentrations among species. The goal was to determine the optimal concentration of AFP III to be added to semen extender to best preserve sperm and their activity.
In the present study, the total number of collected ejaculates was enough to support the experimental design, but the sample size is still relatively small, which may limit the generalizability of the obtained results. Additionally, the use of pooled ejaculates, which was necessary to standardize the experimental conditions, may mask potential interindividual variation among dogs. We recognize these limitations; thus, it is obvious that future research with larger sample sizes and a focus on individual ejaculates is essential to confirm and expand upon our findings.
In this study, after freezing and thawing, the group supplemented with 1 µg/mL AFP III showed the highest total motility and moderate-progressive motility compared with the control and other groups. The results for post-thawed dog sperm motility in this study are comparable to those previously found in dog,33,34 human, 25 goat, 50 and rooster spermatozoa. 28 The previous researchers found that adding AFP III at 1 µg/mL to the freezing extender improved post-thawed sperm motility in those mammalian species. On the other hand, improvements in motility results were observed when AFP III was used at higher concentrations (100 µg/mL) in chimpanzees 18 and lower concentrations (0.1 µg/mL) in buffalo and miniature pigs.23,24 In the current study, the enhancement in total motility is corroborated by the reduction in the percentage of immotile sperm observed at the 1 µg/mL concentration. Even though there was no significant difference in progressive motility, the overall decrease in immotile sperm may still be advantageous, particularly in situations where it is crucial to maintain a higher proportion of motile sperm. According to previous studies, in other species, such an improvement in sperm motility was induced by the stability of the plasma membrane and reduced calcium influx, which can be explained by the blockage of ion channels by AFP to stabilize transmembrane electrolyte gradients.23,51 Conversely, the present results showed exposure to 15 µg/mL AFP III to have the lowest total and progressive motility percentages. In other animals, such as mice, AFP III supplementation at 1, 10, and 100 µg/mL reduced motility and sperm survival rate after freezing and thawing, 52 but it did not affect total motility in rabbit 22 or goat (4 µg/mL) spermatozoa. 29 Additionally, adding AFP III to the freezing extender did not improve the preservation of epididymal goat spermatozoa. 31 These results indicate that several factors, including animal species, AFP concentration, and sample resources, affect the use of AFPs in the freeze–thaw process.
In the current study, sperm viability, acrosome integrity, and apoptosis differed non-significantly between the control and AFP III concentrations. Similarly, other research reported that the viability and acrosome integrity of frozen cow, 20 buffalo, 23 ram, 27 and goat29,31 sperm were not improved by the addition of AFP III. A nonsignificant difference in the sperm apoptosis rate was previously described in dogs. 33 On the other hand, AFP III added to the extender enhanced post-thawed viability and acrosome integrity in humans, 25 boars, 24 roosters, 28 and fish. 21 Acrosome integrity in chimpanzees 18 and goats 50 was improved by the addition of 100 µg/mL and 1 µg/mL AFP III, respectively. In dogs, the addition of 0.75 and 1 µg/mL AFP III significantly increased post-thawed sperm viability and acrosome integrity.33,34 The differences in cooling and freezing rates and the higher glycerol concentration (5%) used in the previous dog experiments,33,34 compared with the present study, might have provided stronger protection for sperm plasma and acrosome membrane. Moreover, in rooster sperm, the percentage of apoptotic sperm decreased significantly in the 0.1- and 1 µg/mL-treated groups compared with the control group. 28 The differences between the obtained findings and previous research into the effects of AFP III on dog sperm can be explained by the differing characteristics of the membrane components among species, impacting the response to freeze–thawing. 53 AFP III primarily affects membranes and ice crystal formation by linking to the membrane phospholipid bilayer, which increases the membrane phase transition temperature and its stability.47,54 Thus, the variations among species and lipid arrangements, interactions with membrane components, and phase transition temperatures might explain the differences in results.
AFP III works through two complementary mechanisms. First, a concentration-dependent mechanism, it adsorbs to the surface of the ice, which inhibits growth and recrystallization through thermal hysteresis. 34 The second mechanism involves the interaction with the plasma membrane directly, which raises the phase transition temperature, tightens the packing of lipids, and reduces ionic leakage, including a decrease in Ca2+ influx. 51 The membrane protection by AFP III depends on membrane lipid composition and the AFP:lipid ratio.47,53 At very high concentrations, however, the shape of the ice crystals becomes spicular or needle-like due to the action of AFP III and may eventually be detrimental to cell survival. These biophysical factors may help explain why a 1 µg/mL concentration of AFP III is often found to be optimal in sperm cryopreservation. At this level, it may provide sufficient ice-surface and membrane coverage to prevent recrystallization while maintaining ionic balance. In addition, previous research indicates a distinct species-specific variability in AFP III efficacy. For example, beneficial effects have been observed at minimal concentrations in buffalo and miniature pigs,23,24 at significantly higher levels in chimpanzees 18 ; however, mice exhibit reduced sperm motility and survival rates at concentrations ranging from 1 to 100 µg/mL. 52 These contrasting outcomes suggest that AFP III efficacy is not general but instead depends on species-dependent differences in membrane lipid organization, resulting in a different optimal concentration range for each species.
Regarding apoptosis, in the current study, treatment with 1 µg/mL AFP III significantly improved sperm motility and reduced immotile sperm, suggesting better preservation of membrane and mitochondrial function. While not directly confirming reduced apoptosis, these findings point to a possible indirect protective effect of AFP III against early cryodamage. Further research using additional apoptosis-specific markers is needed to clarify its potential anti-apoptotic role in canine sperm.
To investigate the effects of AFP III on dog sperm freezing at the proteomic level, the three proteins AKAP4, ATP1B1, and HSP70 were considered in the current study. A slight increase in AKAP4 protein expression was observed in the 1 µg/mL group compared with the other groups (Fig. 4). AKAP4 is localized in the fibrous sheath of dog sperm, 55 as well as in the sperm of mice, 56 stallions, 57 boars, 37 bulls, 58 and humans. 59 This protein is an indicator of the quality and fertility of dog semen 55 as it is a crucial structural protein of the flagellum that regulates sperm motility, hypermotility, capacitation, and fertility.36,37 Moreover, the AKAP4 protein regulates cyclic AMP and calcium-dependent protein kinase A (PKA) and protein kinase C (PKC) signaling. Additionally, AKAP4 is essential for synchronizing the signal transduction of PKA with motor proteins and promotes the sliding of the microtubule doublets of the axoneme. 37
The ATP1B1 protein was also studied. The 1 µg/mL group had the lowest ATP1B1 protein expression, whereas the 10 µg/mL and 2 µg/mL groups showed the highest ATP1B1 expression in experiments 1 and 2, respectively (Figs. 4 and 5). The ATP1B1 protein is a subunit of ATP synthase that must undergo a structural rearrangement to produce energy. ATP1B1 is a potentially useful biomarker for evaluating sperm freezability. 38 According to Nijtmans et al. 60 and Lapaille et al., 61 the absence of ATP1B1 can prohibit ATP synthase from producing intracellular ATP, decrease the respiratory rate, and limit spermatozoa motility. 62 In addition, Ryu et al. 38 found that higher levels of ATP1B1 maintained sperm motility and viability during freezing and thawing. However, in the current study, the ATP1B1 protein did not directly impact sperm motility.
The third studied protein was HSP70, which is detected in sperm during capacitation and the acrosome reaction and is predominantly localized in the acrosome and postacrosome areas. 41 Although the differences were not statistically significant, we observed a numerical increase in HSP70 protein expression in the 1 µg/mL group compared with the other groups (Fig. 4), which may suggest a potential trend worth exploring in future studies. HSP70 is associated with high sperm quality and with viability, motility, and fertility.39–41 HSP70 expression was discovered to be low in infertile men, 63 and Pardede et al. 41 reported a positive correlation among gene expression levels, abundance of HSP70 protein, and sperm motility, suggesting that an increase in HSP70 production can protect sperm cells from the damaging effects of cryopreservation. These results offer initial exploratory insights into potential relationships between protein expression and sperm quality. However, the lack of significance in protein expression highlights the need for further investigation with greater statistical power and refined study designs. These findings identify that those proteins are potential candidates for further evaluation as sperm quality markers.
Conclusion
This study showed that the addition of 1 µg/mL AFP III to freeze extender could enhance the total motility of dog sperm during the freeze–thaw process; however, progressive motility and other kinematic parameters remained unchanged. Moreover, AFPIII within the concentration range applied in this study had no notable impact on dog sperm viability and acrosomal integrity. Additionally, the protein analysis indicated the possible use of protein expression levels as biomarkers to predict post-thawed semen quality and freezability. However, detailed investigations are needed to better understand the proteomic effects of AFP III on freeze–thawing of dog sperm.
Authors’ Contributions
S.I.: Methodology and formal analysis (lead). M.A.: Investigation and writing—original draft (equal). I.-J.Y.: Supervision and writing-review and editing (lead).
Footnotes
Author Disclosure Statement
No conflicting interests exist.
Funding Information
This research was supported by the “Research Base Construction Fund Support Program” funded by Jeonbuk National University in 2025.
Supplemental Material
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.