
Abstract
Organoids, as three-dimensional (3D) self-organizing tissue models, provide physiologically relevant platforms for disease modeling, drug discovery, and regenerative medicine. Effective cryopreservation is essential for their long-term storage, large-scale distribution, and standardized application. This review summarizes the fundamental principles of organoid cryopreservation, including slow-freezing and vitrification strategies, cryoinjury mechanisms, and permeability-based theoretical models, as well as the recent advances in cryoprotective agents. Further progress in organoid preservation is highlighted, focusing on how integrated approaches, such as in situ cryopreservation, hydrogels, and nanotechnology, enhance cryopreservation outcomes. In addition, methods for evaluating the post-thaw structural and functional integrity of organoids are discussed. Finally, we present emerging engineering innovations—including microfluidic control, 3D bioprinting, and biomaterial-assisted systems—that enable precise manipulation and efficient preservation of organoids. Although challenges persist due to the structural complexity and heterogeneity of organoids, the convergence of biophysical principles and engineering technologies offers promising directions toward high-quality and high-throughput organoid preservation.
Introduction
Organoids are derived from stem cells or organ progenitor cells. They have the ability to self-organize into miniature organs with specific physiological functions. Organoids closely mimic the structure and function of the corresponding organs in vivo. 1 Compared with two-dimensional (2D) cell cultures and animal models, organoids provide a more physiologically relevant environment. They also reduce the reliance on animal experimentation. 2 They have become ideal platforms for studying disease mechanisms, testing drug efficacy, and evaluating drug toxicity.
The construction of organoids usually takes several weeks to months, depending on their complexity. 3 The long construction time not only extends the experimental cycle but also increases the overall cost. This poses certain limitations for high-throughput screening and urgent research needs. Cryopreserved organoids address these issues by providing ready-to-use samples without the need for reculturing, thereby significantly reducing the experimental timeline. For clinical and translational applications, organoids need to be preserved long-term without loss of function. 4 In addition, cryopreserved organoids are easy to transport and store, supporting resource sharing and preparing for the establishment of biobanks.
However, the cryopreservation of organoids remains challenging due to ice crystal formation and their inherent structural complexity.5,6 Ice crystal formation can damage the cell membrane and internal structures of organoids. 7 Compared with single cells, organoids as multicellular structures require more considerations. Their structural and size variations, ranging from micrometers to millimeters, make it difficult to apply a uniform cryopreservation strategy. Moreover, hollow organoids (e.g., small intestine-type) and solid organoids (e.g., neural-type) differ in cryoprotectant (CPA) penetration, affecting incubation time and concentration. Additionally, in solid organoids with relatively large sizes (on the millimeter scale), high cellular density can lead to slow heat conduction and uneven cooling during the freezing process. Combined with insufficient penetration of CPAs, this may increase the likelihood of intracellular ice formation (IIF). Even within a single type of organoid, cellular heterogeneity complicates the requirements for effective cryoprotection. Currently, efficient and high-quality methods for organoid preservation remain lacking.
Cryopreservation research began when Polge et al. 8 demonstrated that spermatozoa could survive glycerol-based freezing, introducing the concept of CPAs. Whittingham et al. 9 reported that mouse embryos could survive cryopreservation at −196°C and develop into viable offspring, marking a major breakthrough in multicellular biology. Despite progress in optimizing CPA formulations, cooling and warming rates, ice crystal formation remained a major obstacle. Rall and Fahy 10 introduced vitrification in 1985, enabling the first ice-free cryopreservation of mouse embryos and advancing the field significantly. With the advancement of bioengineering and medical fields, numerous biotechnological tools have emerged, including warming techniques, hydrogels, and nano-assisted technologies.11,12 These developments have also facilitated deeper investigations into CPA toxicity and ice crystal formation mechanisms. 13 Building on these advances, cryopreservation technology has made important breakthroughs in reproductive medicine (such as oocytes, sperm, and embryos), biobanking, and endangered species conservation. As stem cell and regenerative medicine technologies progressed, cryopreservation techniques expanded from cell preservation to the preservation of complex three-dimensional (3D) tissue structures, 14 including tissues, spheroids, and organoids. This evolution enables long-term preservation of high-quality biological samples and supports personalized medicine, disease modeling, and drug discovery.
In this review, organoid cryopreservation is discussed based on the principles of slow freezing and vitrification, which provide frameworks for balancing ice formation and osmotic stress and guiding protective strategies. Building on these foundations, recent advances focus on optimizing cryoprotective agents, improving warming techniques, and developing novel approaches such as in situ and nanotechnology-assisted strategies. Finally, we explore emerging engineering innovations—including microfluidic platforms, 3D bioprinting, and biomaterial-assisted designs—that hold potential to overcome current limitations and enable high-throughput, high-quality, efficient preservation of complex organoid systems.
Fundamental Principles of Organoid Cryopreservation
Cryopreservation preserves biological samples by lowering the temperature to suppress metabolic activity, thereby maintaining cellular viability and function. Ice crystal formation is a major cause of cellular damage, so strategies focus on controlling or avoiding ice formation, primarily through two approaches: slow freezing and vitrification. Meanwhile, permeability models are widely used to describe water and CPA transport across cell membranes, enabling predictions of cell volume changes, dehydration, and intracellular ice risk, and thus guiding the optimization of cryopreservation parameters. Despite these advances, ice formation and other physicochemical stresses remain unresolved challenges and continue to drive research in cryopreservation science.
Slow freezing
Cooling rate
Slow freezing involves gradually lowering the temperature of a solution containing biological samples and CPAs at a controlled rate, thereby reducing biological activity to enable long-term preservation. According to Mazur’s two-factor hypothesis,15,16 cell damage arises from ice crystal formation and solution effects (dehydration-induced injury), both dependent on cooling rate.
Figure 1 illustrates the relationship between cell survival rate and cooling rate during cryopreservation. During slow freezing, extracellular ice draws water out of cells, causing dehydration and solute concentration. Too slow a cooling rate prolongs hyperosmotic stress, while too fast a cooling rate traps intracellular water and forms ice, damaging cell structures. Thus, an optimal cooling rate is critical: slow enough to avoid intracellular ice, yet fast enough to limit solute-induced injury.17,18

Relationship between cell survival rate and cooling rate during cryopreservation.
Currently, organoids are typically slow-frozen in a 10% Dimethyl sulfoxide (DMSO) cryoprotective solution using a controlled-rate freezing container. Although conventional slow-freezing strategies allow organoids to survive after thawing and to partially regain their proliferative and differentiation capacities, significant variations in evaluation methods and assessment criteria across different laboratories make it scientifically difficult to directly compare survival or recovery rates among different studies (e.g., 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) 4 vs. live/dead staining 19 ) nor have they comprehensively addressed the preservation of organoid functionality.
Since organoids are far more complex than single cells in terms of 3D structure, cellular composition, and microenvironment, the existing cryopreservation parameters may not be fully applicable. First, in terms of size and heat/mass transfer, organoids have diameters much larger than single cells, resulting in significant differences in cooling rates between the interior and exterior. This leads to the coexistence of temperature and solute concentration gradients, thereby increasing the risk of IIF or localized supercooling. 5 Second, the diffusion and penetration of CPAs are limited; during rapid cooling, CPA concentrations in the core of organoids may not reach effective levels, increasing the likelihood of ice-induced damage, whereas excessively slow cooling can prolong CPA exposure and amplify solution effects and cytotoxicity. 20 Finally, organoids consist of multiple cell types, each exhibiting distinct tolerances to osmotic stress, CPA toxicity, and cooling rate. A uniform cooling profile often causes selective damage among different cell populations, thereby compromising the overall structure and function of the organoid. 5
Therefore, exploration of organoid cryopreservation can refer to existing experience with the cryopreservation of cell aggregates and spheroids. These structures share certain similarities with organoids in spatial organization and cell–cell interactions, thus providing valuable references to some extent. In terms of CPA design, strategies have included protein-free formulations, 21 macromolecular polyampholytes 22 or low-molecular-weight zwitterions, 23 proline preconditioning, 24 and low-molecular-weight polyethylene glycols (PEGs), 25 with some systems even developed entirely without DMSO. 26 Regarding cooling rates, differences in the responses of aggregates versus single cells under DMSO-based conditions highlight the need to optimize cooling parameters for 3D structures. 27
Ice nucleation
During the freezing process, the solution often experiences supercooling, meaning it remains liquid below its equilibrium freezing point. However, excessive supercooling may cause IIF, which can suddenly occur and grow rapidly, leading to severe damage to cells.
Ice nucleation is a critical process in slow freezing. To control the formation of ice crystals and to ensure that cells are not damaged by the disordered growth of ice crystals during cryopreservation, seeding is performed. Seeding introduces one or more nuclei into the sample, thus controlling ice crystal growth and avoiding the formation of harmful ice crystals inside the cells. The methods include physical and chemical approaches.
The Physical seeding approach is carried out at a specific temperature, such as liquid nitrogen spray, immersion in liquid nitrogen, 28 contact with precooled forceps, or programmed rapid temperature drops,26,29 to induce nucleation and promote more uniform ice crystal formation. Meanwhile, engineering approaches such as ultrasound and lasers can also be employed to induce ice nucleation. 30 Chemical nucleation methods generally involve adding ice nucleating agents (INAs) to the solution. INAs facilitate the formation of ice crystals at relatively higher subzero temperatures, thus reducing uncontrolled or uneven crystallization caused by excessive supercooling. Studies have used cholesterol and pollen washing water as INAs in the cryopreservation of cell spheroids, reducing IIF and thereby improving post-thaw viability.31,32
At present, these nucleation methods have been successfully applied to the cryopreservation of cells, aggregates, or spheroids and have shown great advantages. The ability to precisely control ice crystal formation makes nucleation technology highly promising for the cryopreservation of organoids. Introducing ice nucleation technology into organoid cryopreservation may improve post-thaw viability and preservation quality.
Vitrification
Vitrification is the process during cooling in which the water in the sample forms a glassy state instead of ice crystals, thereby avoiding damage to the cellular structure caused by ice crystals.
Two strategies for achieving vitrification
Figure 2 is a representative supplementary phase diagram of a CPA solution, depicting the thermodynamic state of the solution as a function of temperature and CPA concentration. From this figure, two approaches for achieving vitrification can be identified. 33
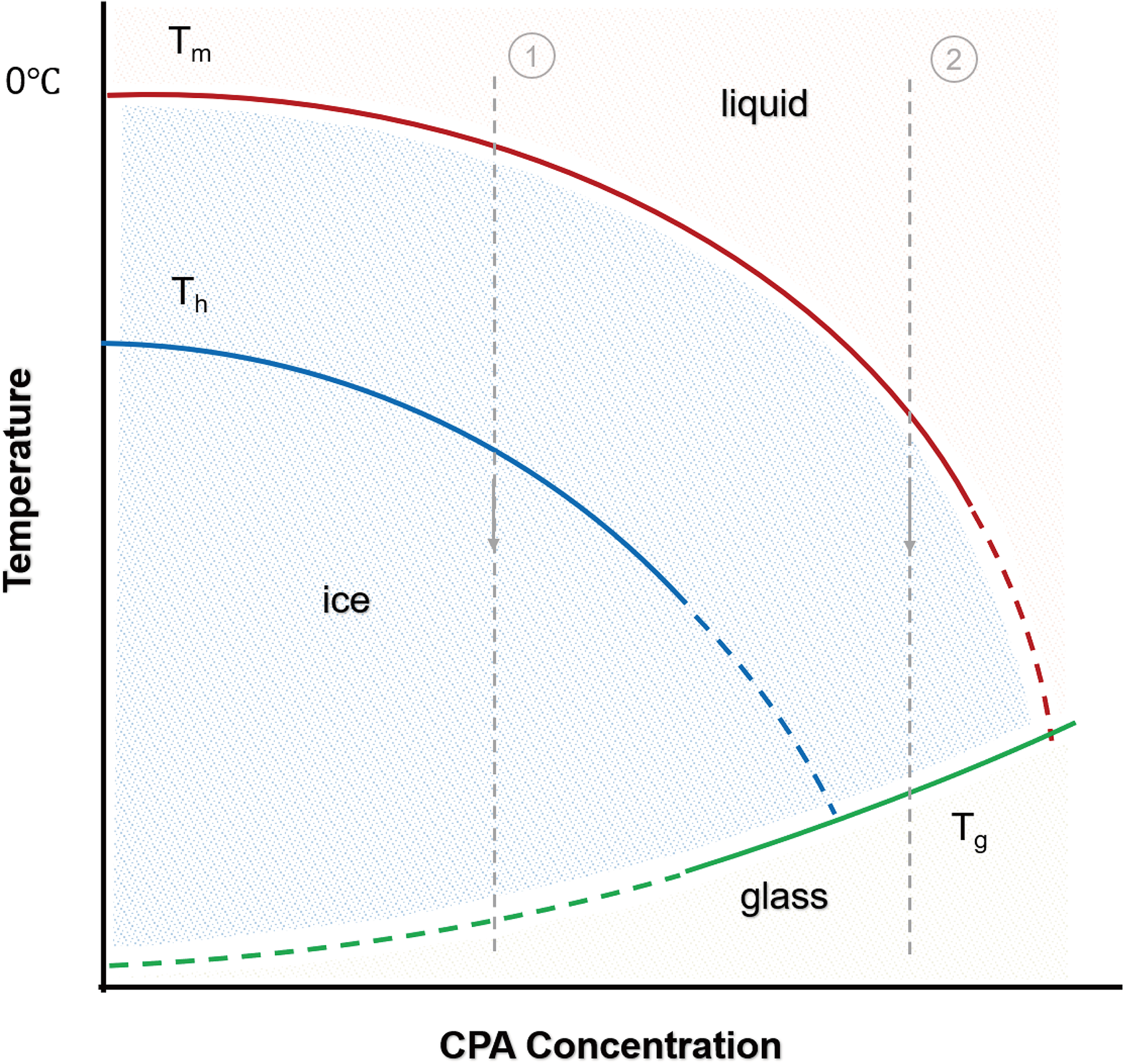
A representative supplementary phase diagram of a CPA solution. Tm is the melting point of ice crystals, Th is the homogeneous nucleation temperature, and Tg is the glass transition temperature. CPA, cryoprotectant.
The first is ultra-rapid cooling of low-concentration protectant solutions. 34 This rapid cooling rate far exceeds the crystallization rate of solute molecules in the solution, so the solute molecules have no time to form an ordered crystal structure but are “frozen” in a disordered liquid structure, forming a glassy state. Further, by dispersing the sample into very small volumes (e.g., using quartz microcapillaries)35,36 and employing materials with high thermal conductivity, 37 heat dissipation efficiency can be significantly enhanced.
Another approach is the use of a higher concentration of CPA solution with a relatively slow cooling rate.10,38 High concentrations of CPAs increase solution viscosity and restrict molecular mobility, effectively suppressing ice formation. As cooling progresses, the solution gradually loses fluidity and eventually vitrifies.
Research on organoid vitrification
Recently, some studies have been conducted on the vitrification of organoids. Ryan et al. 39 reported that using a high-concentration CPA solution and a controlled cooling protocol improved the post-thaw survival of mouse and human intestinal organoid units (OUs) compared with conventional freezing. However, OUs are multicellular aggregates resembling crypt-like structures. Although they contain mucosal and mesenchymal components, they lack the 3D architecture of small intestinal organoids. Moreover, the study did not explicitly clarify whether the method truly achieved an ideal vitrification state free of ice crystal formation.
In 2024, Parham et al. 20 investigated cryopreservation methods for human kidney organoids, comparing the effectiveness of slow freezing and vitrification in maintaining the structural and functional integrity of kidney organoids. After slow freezing, kidney organoids retained a certain level of cell viability (79% and 83% with two different CPAs) but exhibited some degree of proximal tubule dilation and podocyte damage. In contrast, vitrification (using 20% DMSO, 20% ethylene glycol [EG], and 0.75 M sucrose) preserved up to 91% cell viability and was more effective in protecting podocytes and renal tubules.
Permeability model
Fick’s first law.
Due to the complex intercellular connections and tissue architecture of organoids, the application of permeability models during cryopreservation serves as a valuable theoretical tool, offering practical guidance for optimizing the preservation of 3D structures. Therefore, for the cryopreservation of organoids, some researchers have introduced mathematical models to predict permeation behavior. Lee et al. 40 used Fick’s first law to describe the diffusion of DMSO through organoid samples.
Figure 3A simulates the trans-organoid transport of DMSO, revealing how temperature-dependent membrane permeability regulates solute penetration and volume changes. By calculating the diffusion of DMSO across organoid cell membranes over time, the optimal treatment conditions can be estimated to achieve uniform CPA distribution, thereby minimizing cellular damage during cryopreservation and thawing.
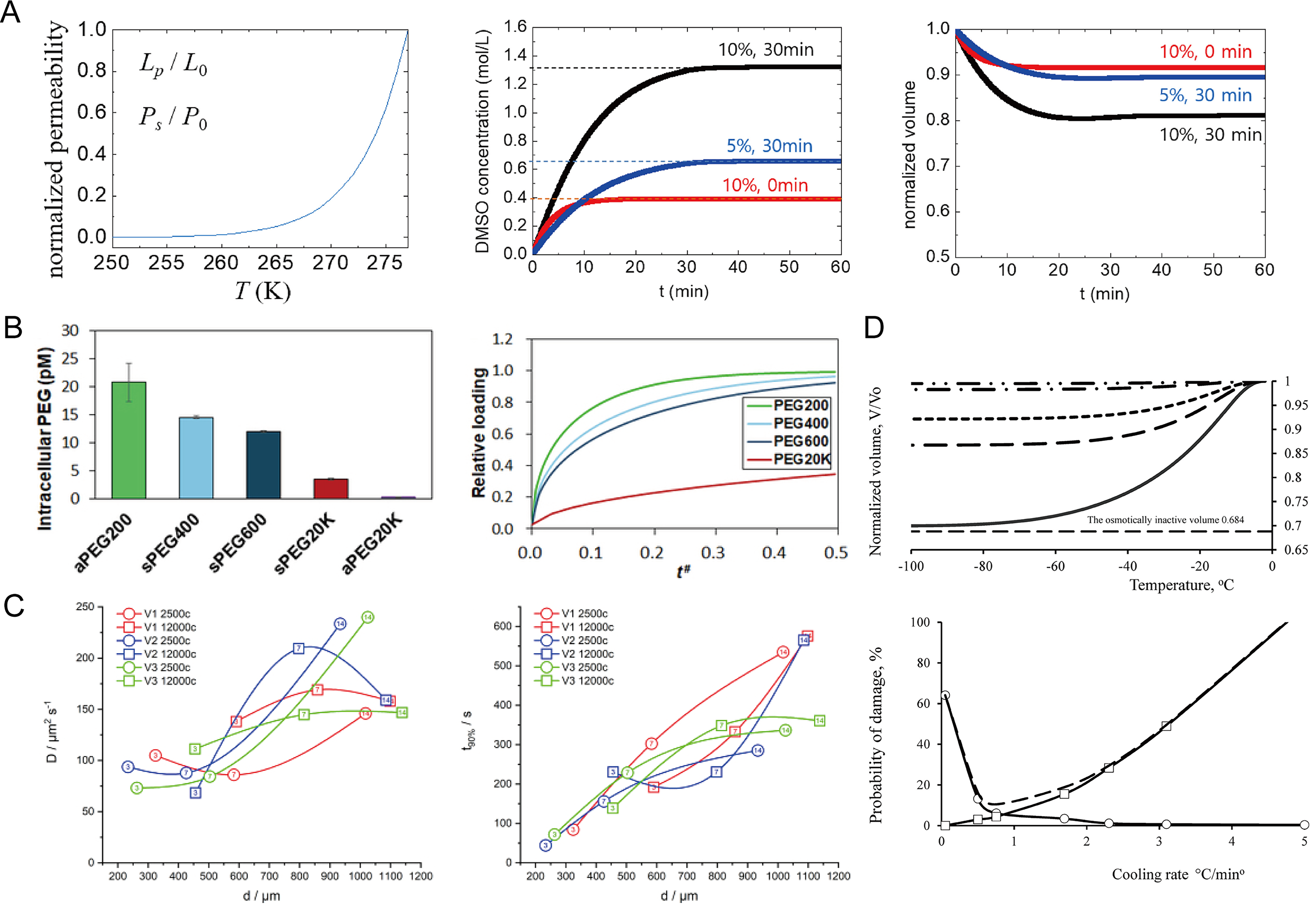
Modeling and simulation of CPA diffusion and cryoinjury in organoids and spheroids.
Fick’s second law
Fick’s first law applies to steady-state diffusion, whereas cryopreservation typically involves unsteady-state diffusion, requiring Fick’s second law for time-dependent modeling. The size of organoids is an important physical parameter affecting CPA permeation behavior. As the diameter increases, the diffusion distance from the external solution to the core region becomes longer, which can significantly prolong CPA penetration time and result in heterogeneous CPA distribution, potentially increasing freezing-induced cellular stress.
Patel et al. 25 used Fick’s second law to model PEG diffusion in stem cell spheres (SCSs). The model predicted PEG distribution and permeation rates for different molecular weights, as shown in Figure 3B. These analyses evaluated the impact of PEG molecular weight on SCS cryopreservation and helped determine suitable pre-incubation times and PEG concentrations.
Similarly, Altmaier et al. 41 used bioorthogonal Raman microspectroscopy (BRM) combined with a spherical diffusion model to study the diffusion kinetics and perfusion time of DMSO in neural stem cell spheroids. Using Raman tracking of DMSO-d6, they monitored concentration changes at the spheroid center. Integrated with the diffusion model, these data yielded the diffusion coefficient and perfusion time, as shown in Figure 3C. These parameters guide CPA incubation optimization and highlight the utility of BRM for improving cryopreservation of 3D cell aggregates.
Kedem–Katchalsky equation
Some researchers have also predicted the optimal cooling rate for cryopreserving spheroids through physical and mathematical modeling. The Kedem–Katchalsky (K-K) equation 42 is commonly used to describe the volume and solute flow of nonelectrolyte solutions across membranes. Gordiyenko et al. 43 used the K-K equations to model the permeation behavior of water and permeable solutes (DMSO) in mesenchymal stem cell–based spheroids during cryopreservation. The article used the K-K equations to calculate the filtration and permeation coefficients of DMSO solutions at different concentrations. These coefficients were then applied to differential equations describing the changes in relative volume and the concentration dynamics of permeating solutes within the spheroid during extracellular crystallization. The model calculated the volume changes of the spheroid at different cooling rates to determine the appropriate cooling rate. Furthermore, two types of damage probability models were established to determine the optimal cooling rate, as shown in Figure 3D. Subsequent experiments showed that the results predicted by the model were largely consistent with the experimental findings.
Although Fick’s second law and the K-K equations are currently applied primarily in the cryopreservation of spheroids, with a deeper understanding of organoid permeation characteristics, these theoretical approaches hold promise for guiding low-temperature preservation of organoids.
Damage
In addition to ice crystal formation, dehydration, and osmotic imbalance, other factors contributing to cellular damage include oxidative stress, protein denaturation, and alterations in the epigenetic landscape. Some of this damage is not solely caused by low-temperature conditions but is also associated with the toxicity of CPAs.
Oxidative stress refers to a state in which the balance between oxidation and antioxidation processes within the organism is disrupted. During the cryopreservation of cells, although metabolic activities are suppressed, the mitochondrial electron transport chain continues to produce reactive oxygen species. At the same time, under low-temperature conditions, the activities of intracellular antioxidant enzymes such as superoxide dismutase and catalase decrease, leading to a weakened cellular antioxidant defense system. Due to insufficient antioxidant capacity, the generated reactive oxygen species cannot be effectively eliminated, resulting in their accumulation within the cells. This accumulation triggers lipid peroxidation, protein oxidation, DNA damage, and other types of cellular injury, ultimately causing cell death. 44 To prevent oxidative stress, antioxidants need to be added to the cryopreservation system. Antioxidants can neutralize free radicals and inhibit the activity of enzymes responsible for free radical generation, thereby interrupting the chain reactions initiated by free radicals and preventing the accumulation of reactive oxygen species. 45
During the freezing and thawing processes, drastic temperature changes can alter the thermodynamic properties of proteins, leading to the disruption of their secondary and tertiary structures. 46 Consequently, proteins lose their normal biological functions, which adversely affect cellular metabolism and physiological activities. Meanwhile, high concentrations of CPAs can cause protein denaturation within cells.47,48 Additionally, changes in the intracellular environment during cryopreservation, such as variations in pH and ionic strength, may also affect the structure and stability of proteins, 49 leading to protein denaturation and inactivation.
The cryopreservation process itself may also induce alterations in the cellular epigenetic landscape. Factors such as low temperature and oxidative stress can affect epigenetic modifications, including DNA methylation and histone modifications, 50 leading to abnormal regulation of gene expression. Moreover, studies have found that treatment with EG and DMSO can significantly alter the epigenetic profile of cells, affecting gene expression and normal cellular functions. 51
Selection and Application of CPAs
Common CPAs
Permeable CPAs
Permeable CPAs can penetrate the cell membrane and enter the intracellular space. They lower the freezing point of both intracellular and extracellular solutions, thereby reducing ice crystal formation and preventing excessive cellular dehydration. DMSO possesses strong membrane permeability and can rapidly penetrate into the interior of cells. DMSO, the most widely used CPA, inhibits ice formation through strong hydrogen bonding with water 52 and enhances membrane permeability to regulate osmotic balance. 53 Although DMSO is the most widely used CPA, it is associated with issues such as cytotoxicity and genetic damage. 54 Similar to DMSO, glycerol also suppresses ice formation and reduces osmotic stress. 55 However, its slower membrane permeability and the difficulty of removing it after thawing make the cryopreservation process more complex.
Nonpermeable CPAs
Nonpermeable CPAs primarily act extracellularly. They increase the osmotic pressure outside the cell, promoting the efflux of intracellular water and enhancing cellular dehydration, thereby reducing the risk of intracellular ice crystal formation. These agents mainly include sugars, polymers, and protein-based substances.
Sugars such as sucrose, trehalose, and dextran stabilize cell membranes, increase extracellular osmotic pressure, and support vitrification.56,57 However, due to their inability to permeate cell membranes, their protective efficacy is limited when used alone, and thus they are typically combined with permeable cryoprotective agents to achieve more effective preservation.
Polymeric compounds such as PEG and polyvinyl alcohol can suppress ice nucleation58,59 and crystal growth while providing surface protection to cell membranes. 60 These materials have been explored as adjunct CPAs to improve structural preservation in 3D tissues and organoids.
Antifreeze proteins (AFPs) inhibit ice crystal growth by binding to ice surfaces and modifying ice crystal morphology.61,62 Due to their strong ice-modulating ability, AFPs have attracted increasing attention for improving the cryopreservation of complex biological systems, including organoids.
Natural deep eutectic solvents (NADESs) are a novel class of green solvents formed by natural components—such as hydrogen bond donors and acceptors—through extensive hydrogen bonding interactions. Typically composed of sugars, amino acids, or organic acids, NADESs exhibit low toxicity and low melting points. 63 In terms of antifreeze mechanisms, NADESs reduce ice nucleation and crystal growth and protect cells from dehydration and membrane damage during cryopreservation through robust hydrogen bond networks, high viscosity, and vitrification potential. 64
Exploration of CPAs for cryopreservation of organoids
Researchers have explored different CPA formulations to improve organoid survival and functional recovery. Studies have validated the effects of DMSO on various organoids. Lee et al. 40 found that although 5% DMSO slightly improved survival, there was no significant difference among the different DMSO concentrations in mouse intestinal organoids. Similarly, Riley et al.65,66 reported that 10% DMSO preserved equine endometrial and feline oviductal organoids effectively. However, most studies focused on DMSO, with limited research on other protective components.
Single-component CPAs often fail to meet the demands of organoid cryopreservation, while composite CPAs can enhance protective efficacy through synergistic effects of multiple components. 67 Han et al. 4 showed that adding Y-27632, a ROCK inhibitor, during cryopreservation significantly reduces apoptosis and improves recovery of intact small intestinal organoids. Xue et al. 68 proposed a novel CPA technology called MEDY, which was used for the cryopreservation of human brain tissue and neural organoids. The formulation included 1% methylcellulose, 10% EG, 10% DMSO, and 10 μM Y-27632. After thawing, the appearance, growth, and function of the organoids were highly similar to those of the fresh, age-matched brain organoids. The MEDY technology can efficiently preserve neural organoids, but its applicability to other types of organoids still requires further investigation.
Commercial cell cryopreservation media are used in the cryopreservation of organoids. However, due to the structural and cellular complexity of organoids, their cryopreservation requirements differ from those of single cells. Specialized cryopreservation media for organoids have emerged69,70; however, their compositions remain undisclosed. CPAs must be customized for each organoid type to maximize survival and functional integrity. Moreover, research on vitrification solutions for organoids is limited, as current formulations are often adapted from embryos or other samples, 19 highlighting the need for organoid-specific development.
Advances in Organoid Cryopreservation Technologies
In situ cryopreservation of organoids
Cryopreservation using cryovials is the most common method, but in situ cryopreservation allows organoids to be frozen directly within porous plates and used immediately after thawing. Elfi et al. 70 showed that bovine colonic organoids preserved in porous plates maintained growth and staurosporine sensitivity similar to nonfrozen controls. The research group subsequently performed in situ cryopreservation on human liver organoids, colonic organoids, and colorectal tumor organoids. 69 A high proportion of viable cells was observed after thawing, and gene expression as well as drug sensitivity was comparable to those in the fresh group. However, extracellular matrix (ECM) behavior and thermal transfer limitations in large 3D structures may cause uneven preservation effects.
In situ cryopreservation has certain advantages compared with the traditional methods involving centrifugation and transfer before cryopreservation. Liu et al. 71 developed a superhydrophobic microwell array chip-based in situ technique for lung cancer organoids, achieving cell survival and drug responses comparable to vitrification in cryovials. This method avoids centrifugation and resuspension during thawing, although CPA residue may still cause damage. Additionally, CPA permeability may be limited, and vessel materials (e.g., plastic or glass) can affect cooling rates, leading to uneven protection.
It is important to note that the impact of the ECM on organoid cryopreservation efficacy is highly dependent on the specific procedural workflow. In conventional protocols, organoids are dissociated from the matrix prior to freezing, rendering the ECM’s influence negligible. However, when organoids are preserved in situ within the ECM, the physical properties of the matrix emerge as decisive biophysical factors for cryopreservation efficacy.
First, the density and porosity of the ECM restrict molecular diffusion, 72 thereby increasing the mass transfer resistance for CPA permeation. Furthermore, the biochemical composition and water-binding capacity of the ECM significantly modulate ice nucleation and propagation. The confined microenvironment within hydrogel pores physically constrains the growth pathways of ice crystals, potentially mitigating mechanical damage to the embedded cells. 73 Crucially, the stiffness (mechanical modulus) of the ECM is essential for maintaining the structural integrity of organoids post-thaw. Evidence suggests that cryogenic-induced degradation of matrix fibers compromises the mechanical scaffolding provided to the organoids, 74 which ultimately leads to the loss of apical–basal polarity. Future research should prioritize tailoring cryopreservation parameters to the specific biophysical profiles of the supporting hydrogels.
Hydrogel encapsulation
Organoids may experience structural fragmentation during cryopreservation. Ice crystal formation mechanically damages the cells and structures of organoids, and volume changes cause stress concentration leading to rupture. Encapsulation of organoids within hydrogel materials reduces direct impact from ice crystals, provides mechanical cushioning, and ensures uniform distribution of CPAs to alleviate localized stress. Hydrogel technology represents an important and effective strategy for organoid cryopreservation.
Hydrogels can inhibit ice crystal formation and serve as sustained-release platforms for CPAs, reducing cellular damage and osmotic stress.75,76 By further integrating microfluidics, samples can be encapsulated within hydrogel microbeads 77 (Fig. 4A–D) or hydrogel fibers78–80 (Fig. 4E). Some studies have encapsulated stem cells, red blood cells, and other samples in microcapsules for rapid freezing or vitrification. The results showed that, compared with nonencapsulated groups, the encapsulated samples exhibited significantly higher viability and required lower concentrations of CPAs, thereby reducing the cytotoxic stress caused by high CPA concentrations. 81
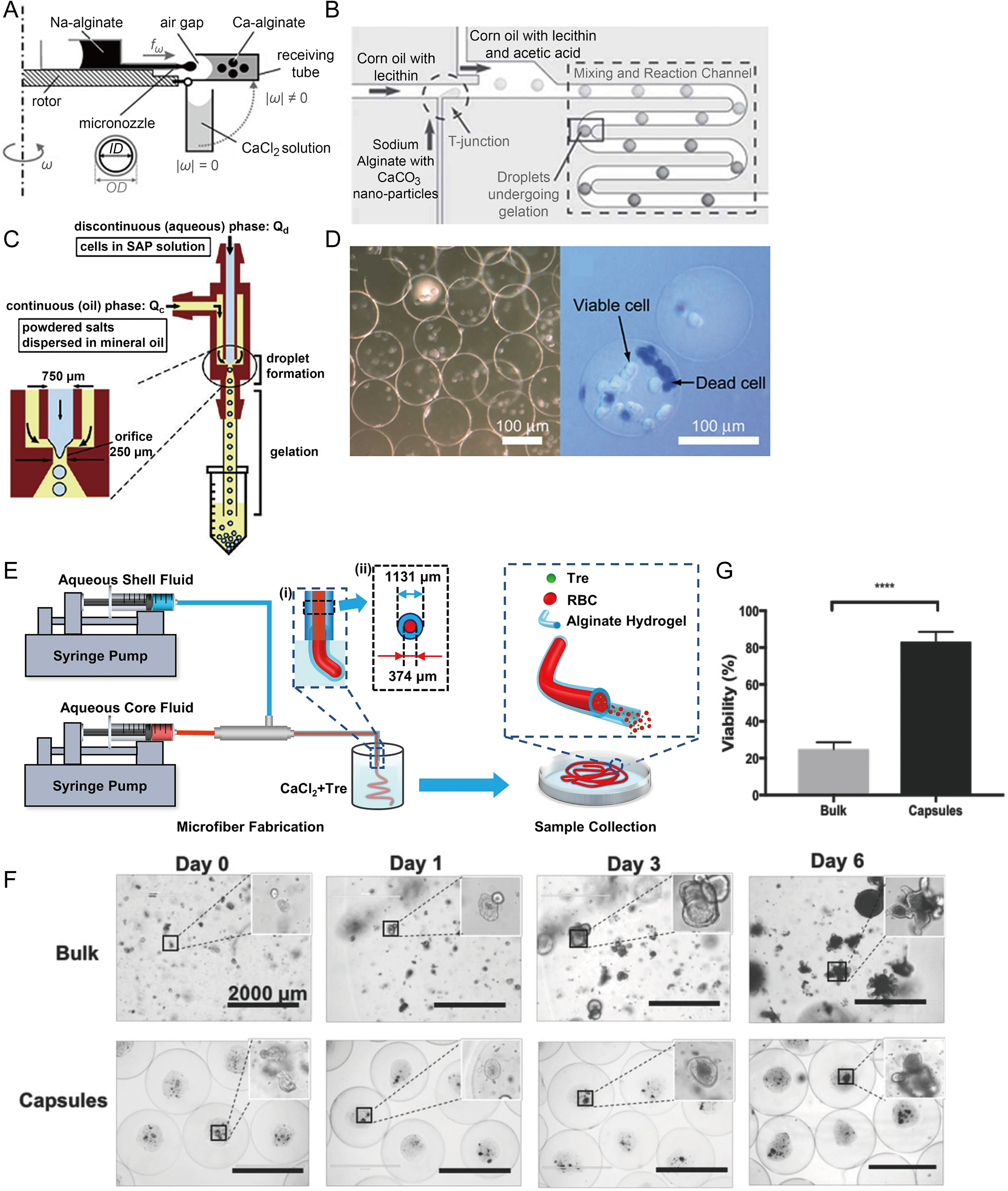
Preparation methods of microbeads and microfibers for cell encapsulation, and their applications in organoid culture and preservation.
Researchers have applied hydrogels for the cryopreservation of organoids. Sakai et al. 82 developed a method using spherical alginate microcapsules templated with gelatin microparticles to culture and cryopreserve PANC-1 cell spheroids. Building upon this approach, Lu et al. 83 created a microcapsule-based scalable platform for organoid production and cryopreservation (Fig. 4F). Microcapsules showed higher cell recovery (∼20%–80%) while maintaining stem cell properties (Fig. 4G). Despite these improvements, mechanical stress such as capsule deformation and the relatively thick shell may limit cell viability and mass transfer efficiency.
Warming
Warming refers to the process of bringing biological samples preserved at ultra-low temperatures back to physiological temperature. The water bath is commonly used for small-volume samples due to water’s high thermal conductivity. However, for large-volume samples, water bath warming is slow, leading to ice recrystallization, temperature gradients, and potential sample damage.
As a result, various advanced warming techniques have been developed, including electromagnetic warming, 84 nanowarming, 85 laser warming, 86 Joule heating-based warming, 87 and ultrasonic warming 88 (Fig. 5A–D). Among these, electromagnetic, nanowarming, and laser-based methods utilize the thermal effects of external physical fields on specific materials, achieving rapid and uniform heating and showing promise for larger-sized samples. 89
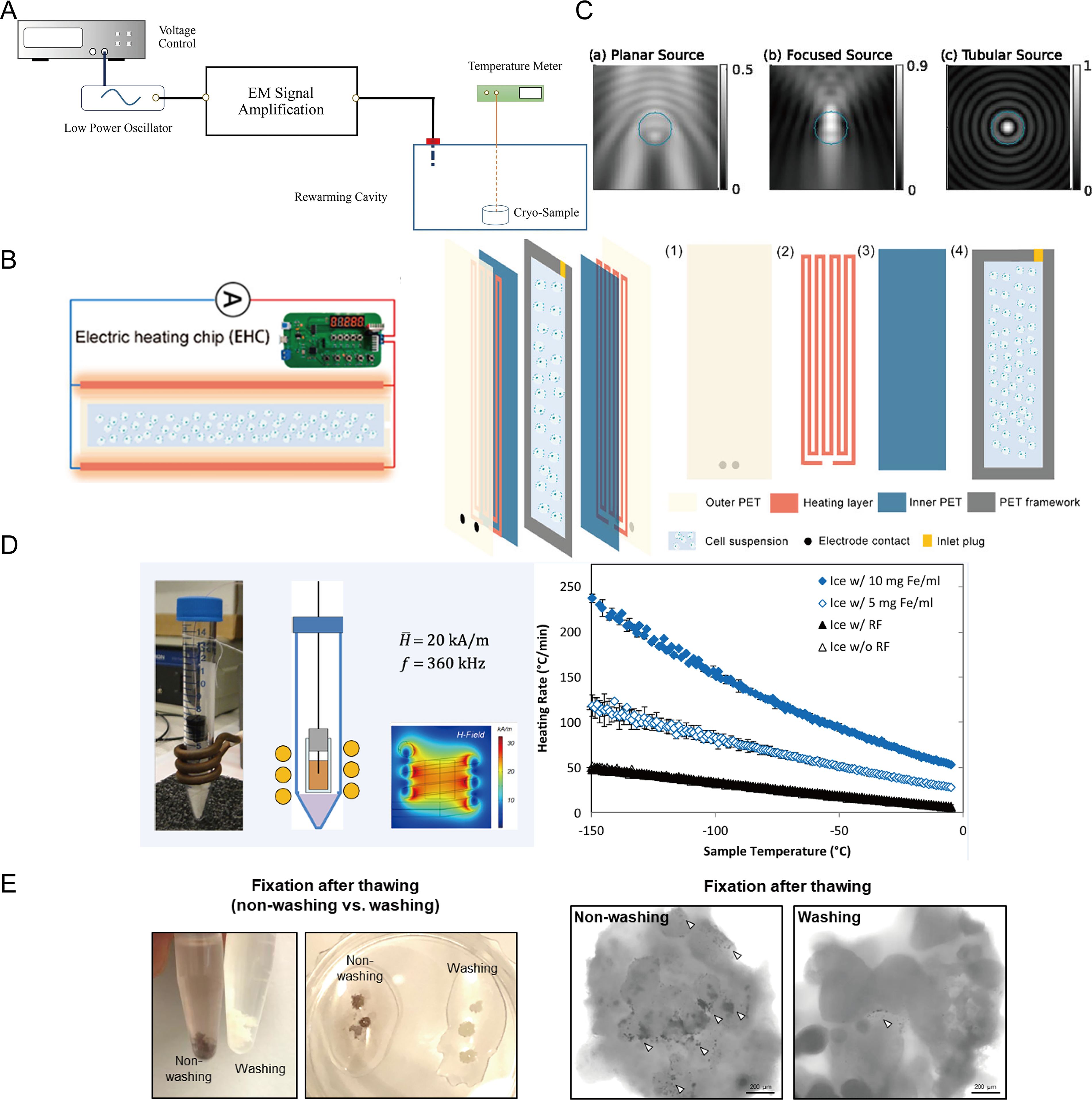
Rewarming techniques and their applications in organoid preservation.
The rewarming of large organoids is often hindered by nonuniform heat conduction. In traditional water bath warming, a temperature gradient exists where the core remains at subzero temperatures while the surface melts, providing a “window” for ice recrystallization, which can mechanically disrupt the 3D architecture of the organoids.
To address this issue, Lee et al. 90 demonstrated a heart organoid rewarming method based on Fe3O4 nanoparticles. Fe3O4 nanoparticles were diluted in a high-concentration CPA solution and incubated to permeate the organoid. The sample was then vitrified in liquid nitrogen and later rewarmed using a high-frequency induction heating device. Within the alternating magnetic field generated by the induction heating coil, Fe3O4 nanoparticles rapidly produce heat. This nanoscale rapid heating facilitates uniform and fast rewarming of the heart organoid. Compared with samples without nanoparticles, this method significantly improved the functional recovery of heart organoids after thawing. Additionally, to mitigate the negative effects of nanoparticles, they were removed through shaking combined with gradient sucrose dilution and elution. However, the nanoparticles were not completely removed after cleaning, which could impact subsequent clinical translational applications (Fig. 5E). Therefore, new methods are needed to achieve efficient removal of nanoparticles or entirely novel warming techniques.
To address the challenge of removing magnetic nanoparticles after nanowarming, several engineering strategies have been proposed. First, during the washing stage, a controlled osmotic gradient can be generated by gradually reducing the concentration of CPAs in the washing solution. This process promotes water exchange and facilitates the outward migration of nanoparticles trapped within the tissue interstitial space.91,92 Second, surface modification of nanoparticles (e.g., PEGylation or phosphate coatings) can reduce particle aggregation in the ECM and minimize nonspecific interactions with biological tissues, thereby improving washout efficiency. 93 In addition, because magnetic nanoparticles respond to external magnetic fields, applying a high-gradient magnetic field during the washing process can help guide particles toward tissue boundaries and accelerate their removal. 94 Furthermore, optimizing nanoparticle size or developing degradable coatings may also reduce the risk of particle retention in tissue. 95
Quality Assessment
Morphological assessment
Morphological changes can reflect the effects of low temperatures on organoids. Bright-field imaging, often combined with image analysis, assesses organoid size and quantity. 96 Transmission electron microscopy (TEM) can reveal ultrastructural damage caused by cryopreservation, for example, Riley et al. 97 used TEM to examine equine oviductal organoids, highlighting potential freeze–thaw effects.
Viability
Viability is a necessary metric for evaluating the effectiveness of cryopreservation. In 3D structures, traditional 2D assays may be inaccurate. 98 Currently, most researchers measure cellular activity in organoids using live/dead fluorescence staining for qualitative analysis. Yet, due to the limited penetration ability of live/dead dyes in 3D structures, it is important to note that this value may not fully represent the viability of all cells within the organoid.
Cell viability in organoids after cryopreservation can be assessed by dissociation followed by trypan blue staining or flow cytometry, although dissociation may reduce viability and requires operator-dependent optimization. There are many methods for measuring cell viability in 3D spheroids.99,100 These methods indirectly assess cell metabolic activity and viability by measuring certain products or changes that occur during cellular metabolism. With the same 3D structure, these methods are also suitable for the determination of cell viability in organoids. However, the viability assays provide only relative values, and their accuracy relies on maintaining consistent cell numbers across experimental groups.
During the growth and development of organoids, cell death is a common occurrence. For instance, in intestinal organoids, a substantial number of dead cells may accumulate within the lumen as the culture period progresses. The accumulation of dead cells may adversely affect the overall viability of organoids and compromise the outcomes of downstream experiments. Hence, it is essential to determine an optimal culture duration for viability assessment and cryopreservation to mitigate the interference caused by excessive cell death.
Molecular biology assessment
In organoids, the maintenance of stem cell pluripotency is the foundation for their continuous growth and expansion. However, during the cryopreservation and thawing process, low temperatures, ice crystal formation, and osmotic stress may damage stem cells, causing apoptosis, altered gene expression, or disrupted signaling pathways, thereby affecting pluripotency and proliferation. Therefore, molecular-level assessments of organoids are necessary.
Common detection methods include gene expression analysis and protein detection. Through Quantitative polymerase chain reaction (qPCR) or RNA-seq technologies, the expression levels of pluripotency-related genes, differentiation markers, and metabolism-related genes can be assessed.69,101 Western blot or immunofluorescence staining can be used to directly evaluate the expression and localization of pluripotency-related proteins. These assessments determine whether organoids maintain their original biological characteristics and storage stability after cryopreservation.
To address the lack of unified evaluation criteria in organoid biobanking, we propose a comprehensive Standardized Quality Assessment Checklist, as presented in Table 1. This framework emphasizes that post-thaw validation must transcend simple cell viability, focusing instead on the preservation of the organoid’s complex 3D structural integrity and lineage-specific functionalities across multiple biological scales. Furthermore, we incorporate the capacity for post-thaw self-assembly and structural reorganization—such as the active reestablishment of apical–basal polarity, functional tight junction formation, and lumen reformation—as a critical functional benchmark. These dynamic morphogenetic processes serve as definitive indicators that the cryopreservation process has preserved the delicate cell–cell signaling and cytoskeletal coordination required for organoids to faithfully regain their physiological architecture and specialized functions during the recovery phase.
Proposed Standardized Quality Assessment Checklist for Post-Thaw Organoids.

Emerging Technologies with Potential for Organoid Cryopreservation
In cryopreservation studies of other biological samples, several emerging technologies have demonstrated significant advantages, not only improving cell survival but also optimizing the preservation process. These approaches have yielded positive outcomes in the handling of oocytes, embryos, and stem cells and thus hold promise for broader application in the cryopreservation of organoids. The following sections will highlight several categories of techniques with notable potential.
Microfluidic technology
In traditional cryopreservation methods, biological samples such as oocytes and embryos are often transferred directly from low-concentration to high-concentration CPA solutions. This sudden change forces cells to undergo drastic osmotic pressure fluctuations, which may result in membrane rupture, rapid volume changes, and severe osmotic injury. To address this issue, researchers have employed microfluidic technology.102,103
Microfluidic technology, through precise designs such as laminar flow mixing or concentration gradients, enables controlled regulation of CPA exposure curves, allowing cells to gradually adapt to extracellular osmotic changes and thereby significantly reducing the risk of osmotic shock.104,105 In addition, while conventional approaches usually require cells to remain in high-concentration CPA solutions for extended periods to achieve permeation, microfluidic platforms can shorten the overall exposure time by performing rapid, stepwise operations in small volumes, thus effectively reducing the chemical toxicity of CPAs. 106
As early as 2009, Song et al. 104 designed a horizontal three-stream microfluidic device (Fig. 6A), introducing cells through the middle channel and CPAs or buffer through the sides. Laminar diffusion enabled the gradual loading and removal of CPAs. Using human hepatocellular carcinoma cells, the microfluidic group showed >80% post-thaw survival, higher than the one-step (∼55%) and two-step (∼70%) methods, reducing osmotic damage and enhancing metabolic activity (Fig. 6B). Heo et al. 107 developed a microfluidic device comprising solution-mixing channels, an oocyte chamber, and cell in/out channels to enable precise, controlled CPA loading of mouse oocytes (Fig. 6C). Compared with traditional stepwise methods, a 10-minute linear gradient reduced oocyte volume changes from ∼25%–30% to below 10%, as shown in Figure 6D–E. An optimized composite gradient achieved equilibration in 15 minutes with only ∼8% volume change and halved total CPA exposure. Zheng et al. 108 compared one-step, multistep, and microfluidic methods for cryopreserving Human umbilical vein endothelial cells (HUVECs) under different CPA concentrations. The microfluidic approach outperformed traditional methods: at 3 M DMSO, cell viability reached 85%, higher than 61% for the multistep method, with proliferation comparable to fresh cells. A two-parameter model showed that the microfluidic method, using a gentle concentration gradient, minimized water flux peaks and toxicity exposure at 3 M DMSO, significantly reducing osmotic, toxic, and centrifugation damage—key factors for improved recovery with high-concentration CPAs (Fig. 6F). Microfluidic technology has shown favorable outcomes in CPA loading and removal across various types of samples, providing important references and promising prospects for the efficient and low-damage preservation of organoids.
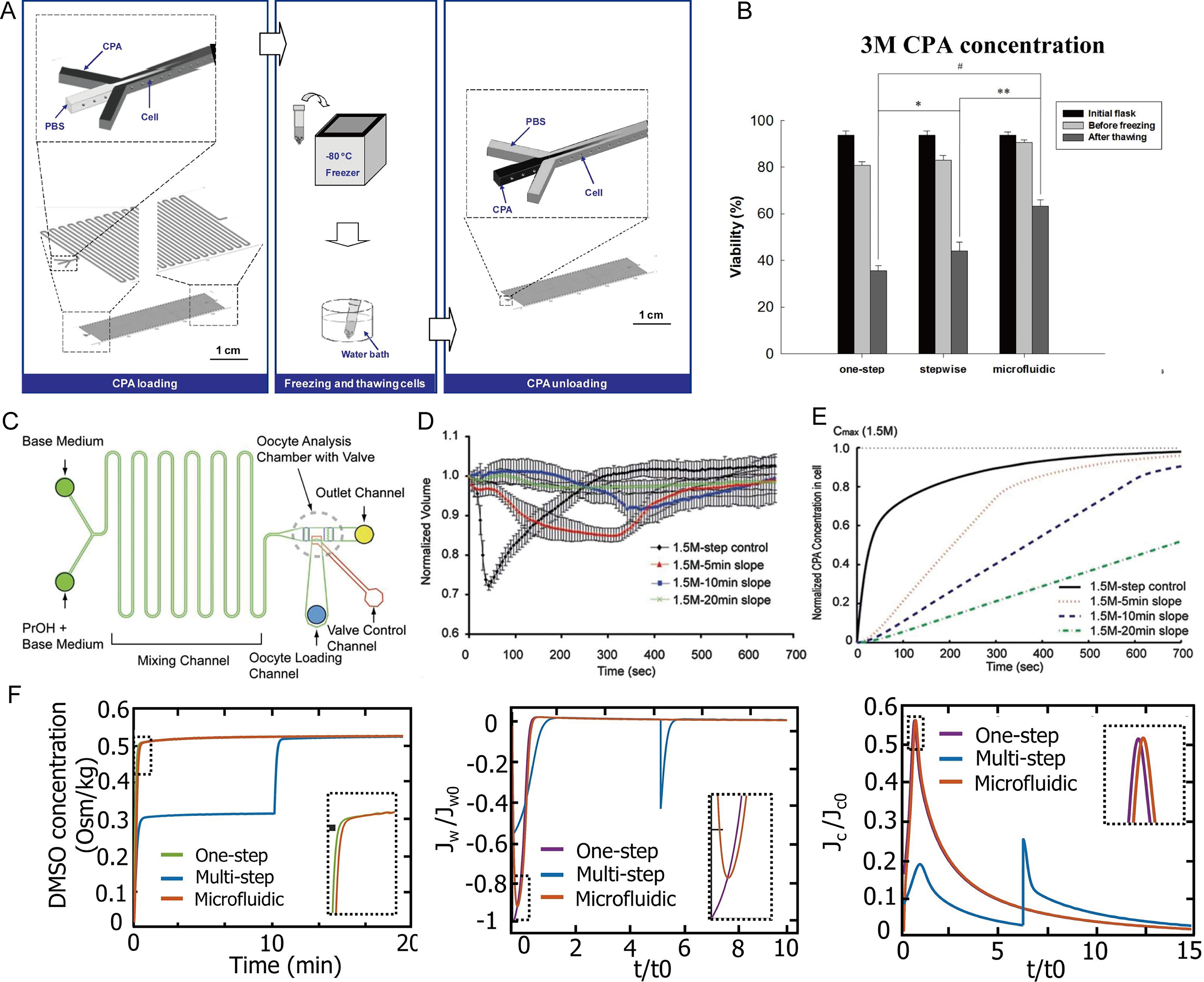
Microfluidic technology for cryoprotectant loading and removal.
The efficacy of microfluidic platforms in mitigating osmotic shock depends heavily on the specific geometric architecture and associated mass transfer characteristics of the organoids.
For hollow organoids, the primary challenge resides in the transepithelial osmotic pressure gradient and the subsequent risk of luminal collapse. This biophysical process is highly analogous to the cryopreservation of oocytes or blastocysts, both of which are exquisitely sensitive to abrupt fluctuations in water flux. 109 Drawing from these established models, microfluidic technology can generate near-linear CPA concentration gradients to smooth water flux peaks, 104 thereby maintaining structural integrity and preventing the disruption of barrier functions.
In contrast, the bottleneck for solid organoids is the interstitial diffusion lag of CPAs into the dense tissue core, a constraint similar to those encountered in engineered tissues or tissue slices. 110 In these scenarios, the advantage of microfluidics extends beyond osmotic regulation; it leverages continuous perfusion to improve solute distribution, potentially overcoming diffusion limits. This facilitates more uniform CPA permeation throughout the tissue matrix while minimizing the risk of hyperosmotic injury to peripheral cells. 111
Consequently, while microfluidics serves as a versatile hardware solution, future research must tailor operational parameters—such as flow rates and gradient steepness—to the specific physical properties of the organoids (e.g., surface-area-to-volume ratio and hydraulic conductivity) to achieve optimized preservation across diverse structural archetypes.
Antifreeze hydrogel
The hydrogel encapsulation strategy has proven effective for preserving biological samples, prompting researchers to investigate hydrogel materials and their antifreeze properties. Among antifreeze hydrogel materials, 112 alginate is the most commonly used hydrogel due to its simple gelation properties. It forms a gel through crosslinking reactions with divalent metal ions, such as calcium ions, and exhibits excellent biocompatibility and biodegradability. With advancements in materials science, novel antifreeze hydrogels have emerged.
GelMA, due to its excellent mechanical properties and biocompatibility, is widely used in tissue engineering. Crosslinking GelMA after soaking in DMSO enables effective inhibition of ice crystal formation and growth during the cryopreservation of cells and tissues.73,113,114 Figure 7A and B shows that in the presence of GelMA, the heat flow curves exhibit no melting peak, indicating its ice-inhibiting capability. Li et al.
115
incorporated
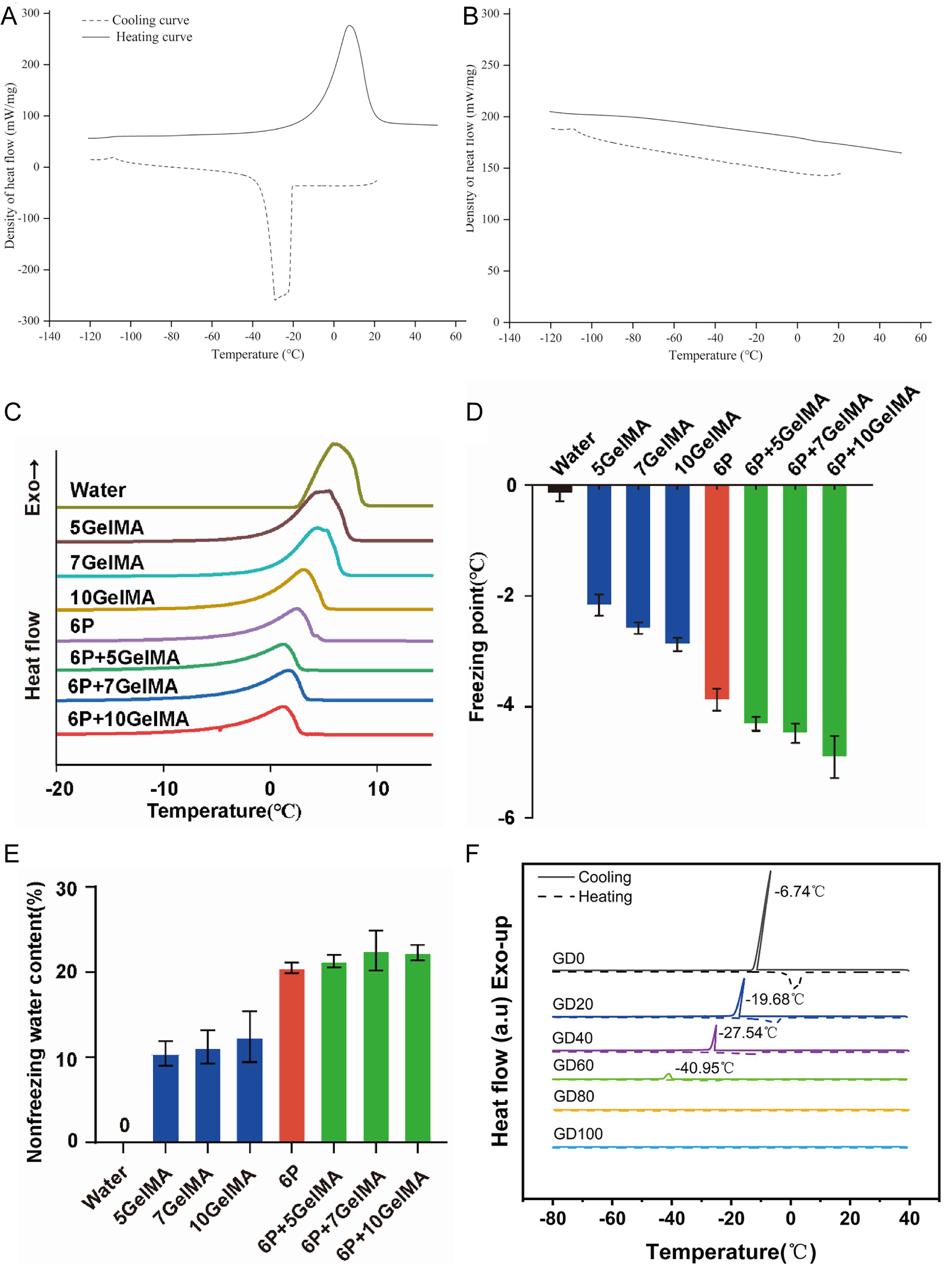
Antifreeze properties of different hydrogel systems.
Inspired by Nagao et al., 117 a multifunctional hydrogel can be developed that serves both as a 3D culture scaffold for organoids and as a cryopreservation matrix, integrating in situ freezing to combine culture and preservation processes. Such a system could maintain the spatial organization and cell–cell interactions of organoids during freezing, potentially reducing ice-induced mechanical damage and osmotic stress. By preserving the structural and functional integrity of organoids, this approach may facilitate long-term storage, high-throughput experimentation, and downstream applications such as drug screening or disease modeling. However, the efficacy of these hydrogels specifically in organoid cryopreservation remains to be experimentally validated, and future studies are needed to optimize hydrogel composition, crosslinking conditions, and freezing protocols for different organoid types.
3D bioprinting technology
The application of bioprinting technology in organoid construction is gradually becoming a frontier direction in the field of biomedical engineering. 118 With the rapid advancement of 3D bioprinting technology, researchers have been able to construct organoids with highly biomimetic structures and functions by precisely controlling the spatial distribution of cells and biomaterials. 119 Several related studies have been conducted, including the construction of bone/cartilage organoids, kidney organoids, 120 droplet-based bioprinting of bladder organoids, 121 and centimeter-scale intestinal tissue fabrication using bioprinting-assisted tissue engineering technology. 122 The integration of 3D bioprinting technology with organoid culture enables precise spatial arrangement of cellular or tissue units, facilitating high-throughput and efficient construction processes while improving organoid reproducibility, structural uniformity, and standardization of culture protocols.
Cryobioprinting integrates the precise fabrication capabilities of bioprinting with the long-term stability of cryopreservation, enabling the production and prolonged storage of complex biological structures. Compared with conventional bioprinting techniques, cryobioprinting primarily addresses the challenges associated with printing low-viscosity bioinks. 123 In cryobioprinting, bioinks are rapidly frozen during the printing process using a chilled plate or low-temperature environment to enhance shape fidelity, followed by immediate cryopreservation to maintain structural stability,124,125 as shown in Figure 8A. While conventional bioprinting excels at constructing complex structures, cryobioprinting enables not only fabrication but also “print-and-preserve,” as shown in Figure 8B. For organoid biobanking, this “print-and-preserve” workflow is a critical engineering solution that minimizes the mechanical stress and loss of cellular polarity typically caused by repeated manual pipetting and centrifugation during traditional cryopreservation protocols.
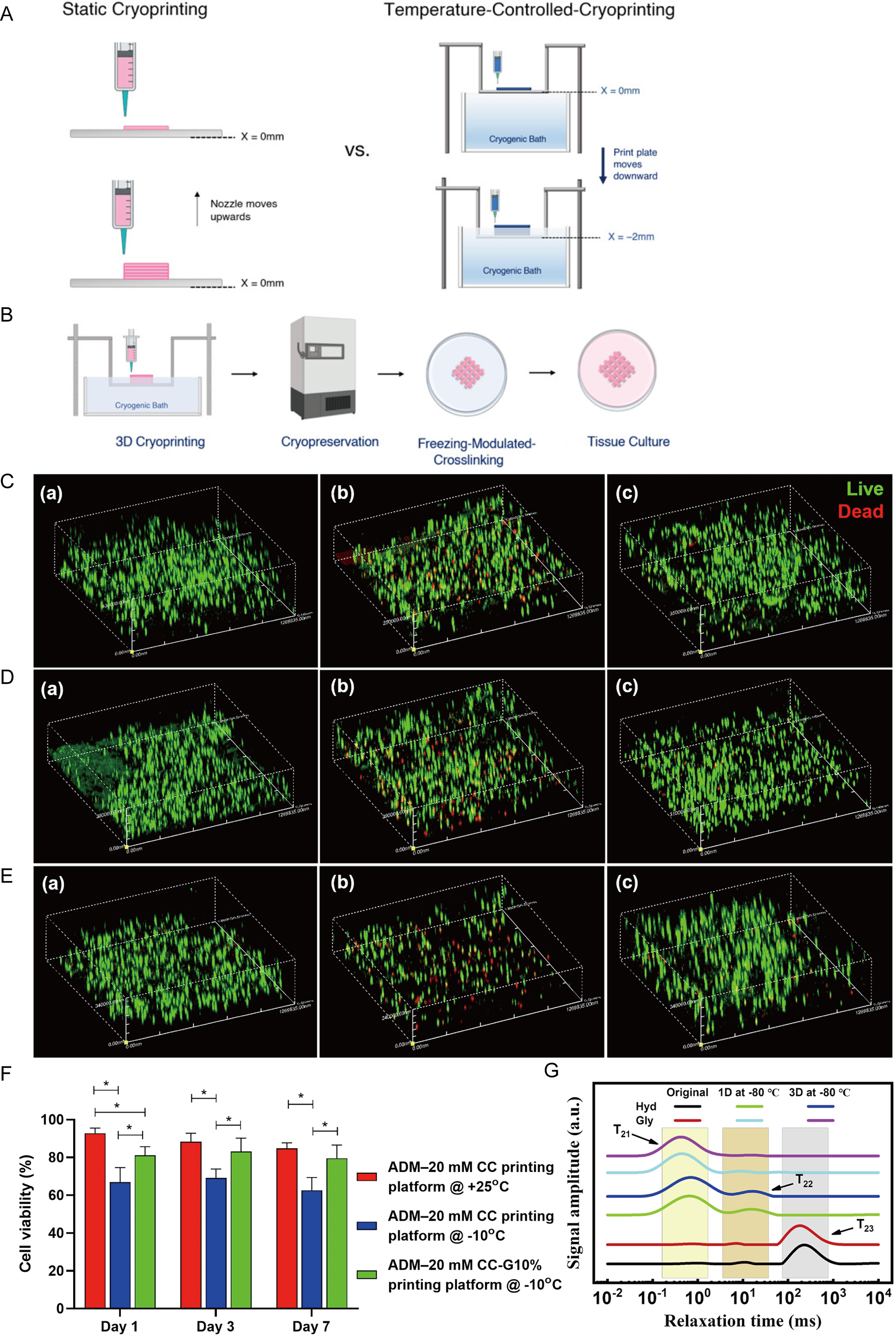
Cryogenic bioprinting technology and its applications in low-temperature preservation.
The core of cryobioprinting lies in the printing process, freezing parameters, and bioink design. Regarding the printing process, the printing speed and pressure must be adjusted according to the rheological properties of the bioink and the tolerance of the cells to ensure structural accuracy and cell viability. 126 Freezing parameters directly affect cell survival and the functional performance of the constructed tissue, requiring precise control of chilled plate temperature, cooling rate, and duration to prevent mechanical damage to cells caused by ice crystal formation. 127 Furthermore, the ability to print sacrificial microchannels within large-scale organoid constructs can significantly enhance mass transfer, allowing for more efficient loading and removal of CPAs, thereby reducing the risk of core recrystallization in centimeter-scale tissues.
The design of antifreeze bioinks is central to the entire system and typically consists of a hydrogel matrix combined with appropriate concentrations of CPAs. 128 These bioinks must not only exhibit good biocompatibility, printability, and mechanical properties but also provide effective cellular protection during cryopreservation. Bioinks such as GelMA incorporated with DMSO and sugars 129 and alginate combined with DMSO or glycerol 130 (Fig. 8C–F) have been developed to enable the fabrication and long-term preservation of cell-laden tissue constructs. In addition, novel hydrogels such as gelatin combined with glycerol 131 and antifreeze starch-based hydrogels 132 have demonstrated promising antifreeze properties (Fig. 8G). Although not yet applied in tissue engineering, they offer potential for expanding applications in the bioprinting field. Meanwhile, to optimize the design of antifreeze bioinks, some researchers have employed computational approaches to predict optimal formulations. Qiao et al. 133 developed various machine learning models to predict the effects of different CPAs (EG, glycerol, and three types of sugars) on post-thaw cell viability in GelMA-based cryobioprinting bioinks. Compared with conventional trial-and-error screening methods, machine learning enables efficient pattern discovery under multivariable and complex conditions, allowing rapid prediction of formulation outcomes and significantly reducing experimental cost and time. This represents a highly promising direction.
Conclusion
Organoid cryopreservation supports personalized medicine, regenerative medicine, and the establishment of biobanks.1,134 However, current preservation strategies are still largely adapted from conventional cell cryopreservation, without systematic optimization for the unique structural and functional characteristics of organoids. Major challenges remain, including the heterogeneity of organoid architecture, limited permeability of CPAs, insufficient understanding of post-thaw functional recovery, and the absence of standardized evaluation criteria. Future progress in this field will depend on the integration of emerging technologies—such as microfluidic control, biomaterial-assisted preservation, and 3D bioprinting—to enable efficient, reproducible, and high-quality cryopreservation of complex organoid systems.
Authors’ Contributions
S.J.: Writing—original draft (lead), formal analysis (lead), conceptualization (supporting), and writing—review and editing (equal). X.Z.: Conceptualization (lead), writing—review and editing (equal), and supervision (lead).
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.