
Abstract
In-group favoritism has often been conceptualized as the flip side of out-group derogation. Whereas research has dissociated these attitudes at the phenotypic level, it is currently unknown whether such dissociation is also evident at the biological level. Here, using an adult German twin sample, which provided ratings on patriotism, nationalism, and prejudice, we tested whether common or distinct heritable influences best explained variation in in-group love and out-group derogation. Results indicated that independent genetic effects accounted for individual differences in in-group love (i.e., patriotism) and out-group derogation (i.e., prejudice). In addition, we observed that nationalism showed common genetic links to both patriotism and prejudice, albeit through distinct pathways. These findings suggest that in-group sentiment is complex at the genetic level as well as at the behavioral level. Future work is recommended to further delineate the specific biological processes underlying these heritable effects.
Introduction
Research has typically treated in-group favoritism as the flip side of out-group derogation (Brewer, 1999); however, numerous studies indicate that this conflation of constructs may conceal important mechanistic distinctions between sentiment toward members of one’s in-group and, in turn, toward individuals belonging to out-groups (e.g., Aboud, 2003; de Figueiredo & Elkins, 2003). Nonetheless, important similarities between in-group and out-group attitudes have also been noted, such as common links to ethnocentrism (Esses, Dovidio, Semenya, & Jackson, 2005; Yakushko, 2009), as well as theoretical support for a common basis (e.g., Tajfel & Turner, 1979). Here, we extended behavioral work in this literature by examining whether national in-group favoritism (i.e., patriotism) is empirically linked or distinct from nationalism (e.g., attitudes concerning out-groups behaving in accordance with local customs) and out-group derogation (e.g., general prejudice toward foreign nationalities) using a genetically informative study design.
In-Group Love and Out-Group Hate: Reciprocal Relations or Distinct Bases?
Much research has operated under the assumption that in-group love reflects the reverse of out-group derogation; that is, those individuals who show preferences for individuals of their in-group will show enhanced out-group dislike (Brewer, 1999). This view can be traced at least as far back as Sumner (1906), who noted that “Loyalty to the group, sacrifice for it, hatred and contempt for outsiders, brotherhood within, warlikeness without—all grow together, common products of the same situation” (p. 12). Later, this perspective found apparent support from work stemming from social identity theory (Tajfel & Turner, 1979), which posits that as personal identity is linked to one’s social identity—with positive self-esteem in turn being a fundamental human need—active derogation can emerge, so as to enhance positive differentiation in favor of one’s in-group versus appropriate out-groups. It is noteworthy, however, that Tajfel (1981) warned against this interpretation with regard to favoritism inevitably leading to derogation; yet, this view is still broadly evident in the literature (Viki & Calitri, 2008).
In contrast, several authors have suggested that in-group love need not mirror out-group derogation. In one early such statement, Allport (1954) argued that “Although we [do] not perceive our own in-groups excepting as they contrast to out-groups, still the in-groups are psychologically primary … Hostility toward out-groups helps strengthen our sense of belonging, but it is not required” (p. 42). Brewer (1999) chiefly revived this position, noting that several studies had provided evidence of a fractionation between in-group and out-group sentiment. For example, Brewer and Campbell (1976) reported no association between positive in-group regard and social distance toward out-groups across 30 ethnic groups in East Africa. Subsequent work has indicated no systematic correlation between in-group positivity and out-group negativity (Licata & Klein, 2002; Struch & Schwartz, 1989).
In addition to the debate detailed above is the distinction that needs to be drawn between patriotism and nationalism: The former defined as love for one’s country; the latter as sentiment concerning the importance of one’s nation and its values and one’s nation’s place in the world. Because both of these attitudinal constructs reflect holding one’s country in high esteem, it is perhaps not surprising to find correlations between these variables (Kosterman & Feshbach, 1989). Of importance, though, Yakushko (2009) argued that some aspects of in-group favoritism (i.e., nationalism) are linked to xenophobia (e.g., prejudice and discrimination), but xenophobia is not linked to in-group commitment (i.e., patriotism). In line with this hypothesis, research has shown moderate links between xenophobic tendencies and both ethnocentrism (i.e., the priority of one’s own group or culture over others) and nationalism (i.e., the priority of one’s own nation over others), although low or negligible links with in-group identification (i.e., commitment to own group) and patriotism (commitment to one’s own nation; Esses et al., 2005). There is good reason, then, to believe that a mixed model—that is, a model positing both common and distinct elements underpin patriotism, nationalism, and prejudice—is the most accurate reflection of human in-group/out-group sentiment.
Genetically Informative Research: Unraveling Etiological Bases
Whereas the debate highlighted above can be addressed through the use of behavioral data, such an approach overlooks the fact that the origins of complex social attitudes, such as intergroup sentiment, are comprised of both genetic and environmental components. For example, in an early genetically informative study of social attitudes, Eaves and Eysenck (1974) found that self-reported radicalism (vs. conservatism) and tough-mindedness (vs. tender-mindedness) were both substantially influenced by heritable factors. Subsequently, Martin et al. (1986) extended the scope of heritable influences on social attitudes, including issues as diverse as sentiment toward gay rights, the death penalty, and abortion: Almost all of the social attitudes examined by Martin et al showed evidence of heritable effects. More recently, work has replicated and extended these initial findings. For instance, Olson, Vernon, Harris, and Jang (2001) obtained similar evidence for heritable effects on a range of social attitudes in an independent, Canadian sample. Research has also found that prosocial sentiment (Lewis & Bates, 2011; Rushton, Fulker, Neale, Nias, & Eysenck, 1986), religiosity (Lewis & Bates, 2013b; Waller, Kojetin, Bouchard, Lykken, & Tellegen, 1990), and attitudes to homosexuality (Verweij et al., 2008) all contain heritable effects. For almost all of these observations, heritable effects accounted for between 25% and 50% of phenotypic variance, indicating that moderate-to-large genetic influences are common for social attitudes. The residual variance is broadly accounted for by environmental factors. Accordingly, because such attitudes can vary in substantial ways due to both genetic and environmental factors, a full understanding of the relations between positivity toward one’s own group (e.g., patriotism) and negativity (e.g., prejudice) toward out-groups will be impoverished without the use of a genetically informative study design.
Of specific relevance to the current study aims, etiological bases underlying intergroup sentiment have also been shown to reflect both genetic and environmental factors (Lewis & Bates, 2010, 2013a; Orey & Park, 2012). Lewis and Bates (2010) demonstrated that religious, ethnic, and race favoritism (preferences for members of one’s own group in these domains) were each individually heritable, but also shared a common heritable basis. Orey and Park (2012) extended this work confirming a heritable influence for race favoritism (using a thermometer measure to tap warmth to members of one’s racial in-group vs. racial out-groups). Subsequently, Lewis and Bates (2013a) established that the two measures of intergroup sentiment noted above shared heritable influences with both traditionalism/authoritarianism and openness. To date, however, no work has explicitly assessed whether genetic bases extend beyond in-group favoritism (as broadly captured in the studies by Lewis & Bates, 2010, 2013a; Orey & Park, 2012) and also underpin out-group derogation. Moreover, no work to date has examined whether in-group love and out-group derogation show common or dissociable genetic elements (assuming that out-group derogation is itself heritable).
The Current Study
With the above in mind, the current study utilizes the classical twin design in order to investigate the sources of the links between in-group love and out-group derogation. We use a multivariate, multigroup structural equation modeling framework in order to decompose behavioral variation and covariation within and between measures of patriotism, nationalism, and generalized prejudice toward foreign nationalities into genetic and environmental components common and specific for in-group love and out-group derogation.
To test the competing perspectives detailed above, we examined a set of genetically informative models reflecting these theoretical positions. Model 1 tested the position that patriotism, nationalism, and prejudice share common genetic and environmental bases, although we also allowed individual genetic and environmental influences to affect each of these variables. Moreover, we tested two variants of this “common factor” model: Model 1a tested whether genetic and environmental influences influencing a single common factor fitted the data well; Model 1b tested a similar principle but relaxed the restriction on genetic and environmental influences acting on a single common latent factor, instead serving to explain variation in the measured variables through a common genetic factor, and common shared and nonshared environmental factors; Model 2 posited that patriotism is a distinct construct from nationalism and out-group derogation, in line with work from Brewer (1999), and only allowed genetic and environmental influences on patriotism that were distinct from nationalism and out-group derogation; Model 3 posited that patriotism and nationalism, and nationalism and out-group derogation are linked, but not patriotism and out-group prejudice, in line with work showing both commonalities and distinctions between these variables (see Yakushko, 2009). Conceptual representations of each the theoretical models are detailed in Figure 1 (also see Analyses section for more details).
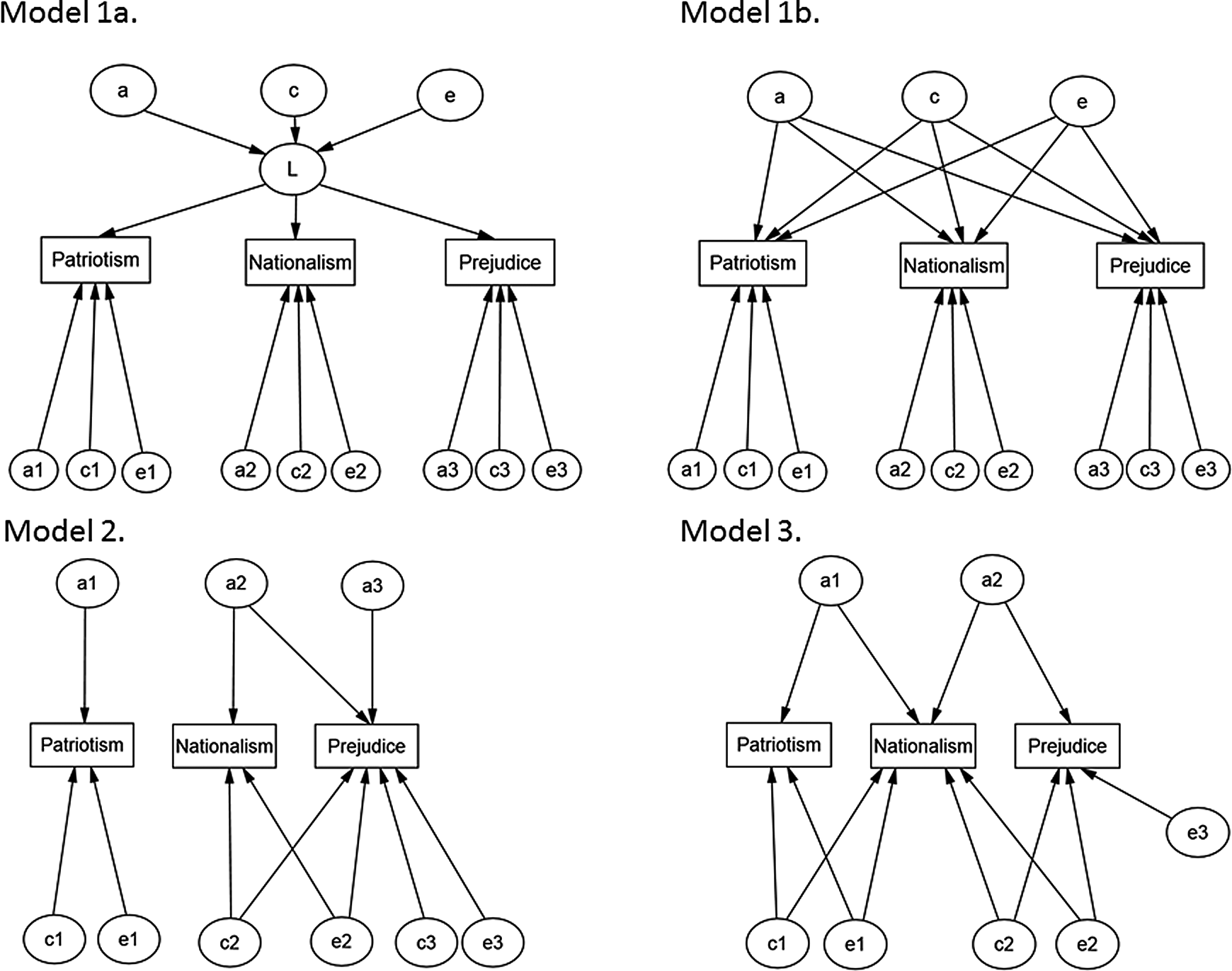
Graphical representation of the theoretical models.
Method
Participants
The sample was drawn from the Jena Twin Study of Social Attitudes (Stößel, Kämpfe, & Riemann, 2006). This sample consisted of data from 875 individuals including 452 monozygotic (MZ), 336 dizygotic (DZ), and 87 unmatched twins reared together. About 74% were females. Mean age was 34 years (SD = 13.6). All twins were German by birth. The sample was heterogeneous with regard to education and occupational status.
The sample was recruited by contacting multiple-birth registries of hospitals, registration offices, twin clubs, and through media calls for participation. Data were collected via paper questionnaires sent to participants in order to estimate genetic and environmental effects on a range of social attitudes and personality traits (Kandler, Riemann, Spinath, & Angleitner, 2010), including the variables analyzed in the current study. Detailed descriptions of the sample and data collection are presented by Stößel, Kämpfe, and Riemann (2006).
Measures
German patriotism (example items: “I love Germany;” “I am not proud to be German” [reversed scored]) and nationalism (example items: “People who do not dearly support Germany and do not respect the culture should live somewhere else;” “Germans should encourage rejecting foreign citizens who do not like Germany”) were measured each by 4 items using a 5-point scale. Internal consistency was α = .61 for nationalism and α = .57 for patriotism. Prejudice toward four national out-groups—Turks, Poles, Italians, and Swedes—was measured by seven self-constructed prejudicial adjective pairs (e.g., “modest–arrogant;” “likeable–unlikeable;” “friendly–unfriendly;” and “trustworthy–backstabbing,” for each nationality) followed by a 5-point scale. Internal consistency was α = .86.
Because of the modest Cronbach’s αs for patriotism and nationalism, in order to assess whether these constructs reflected a coherent underlying basis we submitted the items of each of these scales to confirmatory factor analysis, examining whether a model containing two correlated latent factors—latent patriotism defined by the 4 patriotism items, and latent nationalism defined by the 4 nationalism items, with no additional covariances specified in the model—provided a good fit to the data. The fit statistics for this model indicated that the hypothesized structure was a good representation of the data: χ2(19) = 102.37, p < .01; Comparative Fit Index = .91; Root Mean Square Error of Approximation = .068. Path loadings from latent factors to their indicators were broadly moderate-to-large (for latent patriotism: .27, .34, .64, and .74 and for latent nationalism: .21, .53, .62, and .73).
Analyses
The classical twin design partitions observed variation into three latent components: Additive genetic influences (A), shared environmental influences (C; environmental influences fostering similarities within twin pairs), and nonshared environmental influences (E; environmental influences serving to make individuals within a twin pair less similar). Genetic effects are inferred when MZ twins are more similar than DZ twins, whereas shared environment effects are inferred when MZ twin correlations are less than twice that of the DZ twins. Nonshared environmental effects are inferred when MZ twins are correlated less than at unity for a given trait, and this variance component thus also contains measurement error. Whereas these heuristics provide an instructive guide to the pattern of relative genetic and environmental effects, modern approaches typically utilize a multigroup structural equation modeling framework, which facilitates formal tests of parameter significance, as well as for the estimation of parameters in multivariate models (Neale & Maes, 2004), such as in the current analyses.
Prior to conducting the analyses, we controlled for the effects of age and sex, and standardized residuals were used in all subsequent analyses in line with standard practice (McGue & Bouchard, 1984). The models were estimated by full information maximum likelihood analysis using OpenMx (Boker et al., 2010a, 2010b) and R (R Development Core Team, 2009). Models were compared both against other theoretical models, as well as a Cholesky/baseline saturated model (Neale & Maes, 2004), using the −2 log likelihood difference (Δ−2LL) test for nested model comparisons and the Akaike information criterion (AIC) for descriptive comparisons of non-nested models.
Results
We examined phenotypic correlations between our measures using one twin at random from each pair. Nationalism showed positive links to both patriotism (r = .34, p > .01) and generalized prejudice (r = .28, p < .01), but patriotism and generalized prejudice were not significantly interrelated (r = .01, p > .05).
For each variable, MZ twins were more highly correlated than the DZ twins indicating a role for genetic influences (see Table 1). In addition, DZ twins were correlated at more than half of the correlation of the MZ twins, indicating a role for shared environmental effects. Finally, the less than unity correlations between the MZ twins indicated a role for nonshared environmental effects. Of interest, DZ opposite
Twin Correlations for In-Group and Out-Group Measures and Variance Component Estimates.
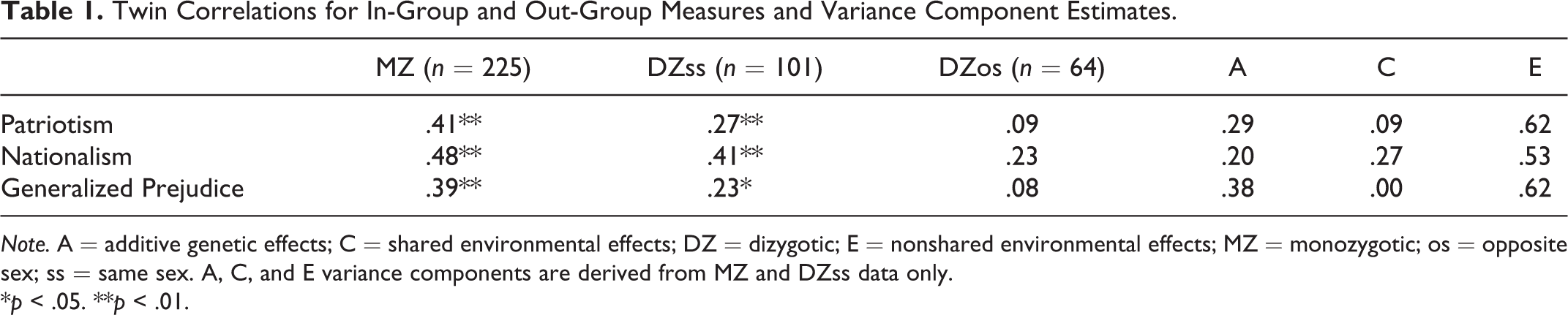
Note. A = additive genetic effects; C = shared environmental effects; DZ = dizygotic; E = nonshared environmental effects; MZ = monozygotic; os = opposite sex; ss = same sex. A, C, and E variance components are derived from MZ and DZss data only.
*p < .05. **p < .01.
We next moved to tests of our core hypotheses. As detailed above, we tested four models reflecting competing theoretical positions regarding the relationship between in-group love and out-group derogation. Fit statistics for each of the models are detailed in Table 2. Our model comparisons for Models 1–3 were adjudicated against the full Cholesky/baseline model. Model 1a, reflecting a single common influence to in-group love and out-group derogation, and Model 1b, specifying common genetic and environmental influences to in-group love and out-group derogation, provided a worse fit compared to the Cholesky/baseline model (i.e., larger AIC values). Model 2, specifying no overlap between patriotism and both nationalism and prejudice (although allowing common paths between nationalism and prejudice), provided a poorer fit to the data compared to the Cholesky/baseline model, Δ−2LL(df = 6) = 77.39, p < .001. Model 3, reflecting common links between patriotism and nationalism, and between nationalism and prejudice, but not between patriotism and prejudice, fitted not significantly worse compared to the baseline model, Δ−2LL(df = 5) = 1.33, p = .93, and so was retained as our preferred model for subsequent tests. Of additional note, we specifically examined the path from a1 to prejudice in a modified version of Model 3 to formally test if this parameter was redundant: This path was estimated at close to zero (.02) and was not significant, Δ−2LL(df = 1) = .08, p = .78, confirming our assessment that no genetic links were present between patriotism and prejudice.
Fit Statistics for All Theoretical Models.

Note. −2LL = −2 log likelihood; AIC = Akaike information criterion; CP = common pathway; EP = estimated parameters; IP = independent pathway; NA = not applicable; p = p value for the χ2 ratio test between the theoretical model and the Cholesky/baseline saturated model.Author Biographies
Next we examined whether the common genetic and environmental influences on patriotism and nationalism (a1 and c1), and nationalism and prejudice (a2 and c2), respectively, were significant. The paths from a1 and c1 could be individually removed from the model without significantly worsening fit, Δ−2LL(df = 2) = 2.42, p = .30; Δ−2LL(df = 2) = 0.30, p = .86, respectively; however, these influences could not be removed simultaneously, Δ−2LL(df = 4) = 39.01, p < .01: As such, we retained both of these common genetic and shared environmental influences on patriotism and nationalism. In contrast, the paths from a2 were significant, Δ−2LL(df = 2) = 9.21, p < .01, although for c2 the paths were nonsignificant, Δ−2LL(df = 2) = 0.81, p = .67, and so were removed from our final model. No other paths were tested. The final model is displayed in Figure 2.
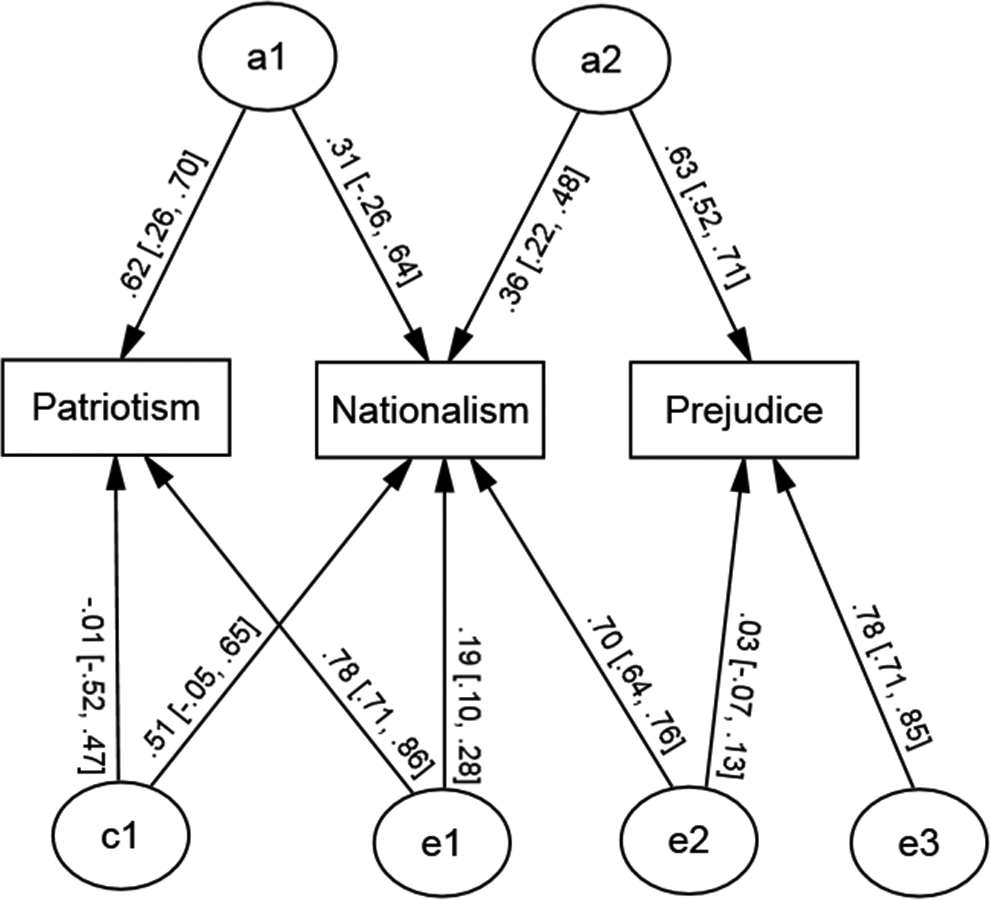
Final model of genetic and environmental influences on patriotism, nationalism, and prejudice.
Discussion
The current study sought to determine whether in-group love and out-group derogation reflect common or distinct constructs using a genetically informative design. The key result was that whereas in-group love was influenced by heritable factors—consistent with previous work (Lewis & Bates, 2010, 2013a)—these heritable influences were not shared with out-group derogation. That is, independent genetic effects accounted for individual differences in in-group love and out-group derogation. Of further interest, though, was the observation that nationalism showed common genetic links to both patriotism and prejudice, albeit through distinct pathways. The current results are consistent with a range of behavioral studies emphasizing both the common (Yakushko, 2009) and the unique (Brewer, 1999) bases underlying in-group and out-group sentiment, and now extend these observations to demonstrate that the complex architecture underlying these phenotypes is also present at the genetic level.
These findings open new questions concerning the underlying biological substrates reflecting these heritable effects on in-group love and out-group derogation. One possible substrate of in-group love may be variation in oxytocin production and reception: Oxytocin has been linked to affiliative behaviors (MacDonald & MacDonald, 2010) and recent work has demonstrated that administration of oxytocin increases favoritism for members of one’s in-group, although does not enhance out-group derogation (De Dreu et al., 2010, 2011). Out-group derogation may, in part, reflect variability in amygdala functioning: For instance, in both Black and White individuals, increases in amygdala activation have been reported in response to viewing faces of out-group versus in-group members (Hart et al., 2000; see also Cunningham et al., 2004). Of importance, however, Schreiber and Iacoboni (2012) report that amygdala activation may not represent the in-group/out-group distinction per se, with amygdala activation also enhanced when viewing photographs of norm-violating in-group members. Prejudice, then, may reflect (at least in part) concerns about appropriate social conduct rather than group membership per se (Duckitt & Sibley, 2010), with amygdala function (or neural circuits functionally linked to the amygdala), reflecting this sensitivity to violations of social norms.
Of importance, shared genetic effects, particularly for nationalism and prejudice, were observed, although this pattern was not observed for nonshared environment effects, which acted independently, showing no overlapping influences across the measures. These nonshared environmental effects were large, however, consistent with other research in this field (Lewis & Bates, 2013a; Orey & Park, 2012), and greater in magnitude than the heritable influences observed here. These effects reflect the importance of environmental forces for generating individual differences in intergroup sentiment, but this variance component also contains measurement error and thus may be an overestimate of nonshared environmental effects. Finally, we observed some, albeit limited, evidence for shared environment effects on nationalism: These effects were not formally significant in either univariate or multivariate models, but were indistinguishable from additive genetic effects and also moderate in magnitude. This observation, although tentative, is of some interest as evidence of shared environment effects is rare in behavior genetic research (Turkheimer, 2000) and suggests that rearing and/or social environments may impact on nationalistic sentiment. This observation may reflect more general social and cultural features concerning the origins of nationalism or population-specific effects. Assortative mating may also account for this effect, however: If parents assort on the basis of social attitudes, such as nationalism, and these traits are heritable, then DZ twins will (on average) share more than 50% of variable DNA and additive genetic effects will, in turn, be underestimated and shared environment effects overestimated. As such, further work is needed to establish the generalizability and robustness of these findings.
Certain limitations in the current study require mention. First, although significant familial influences were observed to be common for patriotism and nationalism, these additive genetic and shared environment influences were not able to be formally distinguished. As such, further work is recommended that incorporates larger samples to provide greater statistical power for parsing such effects. Second, scale internal reliabilities were modest for patriotism and nationalism (Cronbach’s α < .65); future work utilizing more reliable measures will be valuable. Third, the findings reported here were based on a sample of German twins and may not generalize to other populations. Research seeking to extend these findings to broader populations will be of importance.
In summary, we demonstrate that heritable effects underlying in-group love and out-group derogation are dissociable. Of some interest, however, was the observation that nationalism showed common genetic links to both patriotism and prejudice, albeit through distinct pathways. These findings emphasize the need to consider group sentiment as a complex phenomenon, with multiple underlying biological factors.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.