
Abstract
Ovarian hormones significantly influence dysregulated eating in females. However, most women do not develop appreciable disordered eating, which suggests that ovarian hormones may not affect all women equally. We examined whether individual differences in trait negative affect (NA) moderate ovarian hormone–dysregulated eating associations in 446 women who provided saliva samples for hormone measurements and ratings of NA and emotional eating daily for 45 consecutive days. Women were at greatest risk for emotional eating when they had high trait NA and experienced a hormonal milieu characterized by low estradiol or high progesterone. Although effects were evident in all women, the combination of high trait NA and high progesterone was particularly risky for women with a history of clinically significant binge-eating episodes. These findings provide initial evidence that affective and hormonal risk interact to promote emotional eating and that effects may be amplified in women with clinically significant binge eating.
Ovarian hormones (i.e., estradiol, progesterone) play an important role in risk for dysregulated eating in females during puberty and adulthood (Culbert, Sisk, & Klump, 2018; Klump, Culbert, & Sisk, 2017; Mikhail, Culbert, Sisk, & Klump, 2019). At puberty, low estradiol appears to potentiate latent genetic risk for binge eating in girls (Klump et al., 2018). Relatedly, female animals that have undergone prepubertal ovariectomy (which removes the body’s primary source of estradiol) have shown increased rates of binge eating in adulthood (Klump et al., 2020). In adult women, natural fluctuations in ovarian hormones across the menstrual cycle are associated with phenotypic changes in binge eating and related behaviors, such as emotional eating (i.e., the tendency to eat in response to negative emotions, which strongly correlates with binge eating and is considered a core mechanism; Haedt-Matt & Keel, 2011; Ricca et al., 2009). Understanding these hormonal effects may therefore be critical to understanding why some women experience clinically significant dysregulated eating.
Extensive animal research suggests that estrogen protects against binge eating and related phenotypes (e.g., excessive food intake) in adult females, whereas progesterone antagonizes the protective effect of estrogen and increases binge eating (Asarian & Geary, 2006; Kemnitz, Gibber, Lindsay, & Eisele, 1989). Results across the menstrual cycle in women are consistent with these findings. Emotional eating tends to be lowest just before ovulation, when estradiol is at its peak, and highest during the midluteal phase, when progesterone is high and estradiol is moderate (Klump, Keel, Racine, et al., 2013). Correspondingly, the interaction between estradiol and progesterone is the strongest predictor of emotional eating in models with direct measures of ovarian hormones in community samples (Klump, Keel, Racine, et al., 2013). In other words, women are most susceptible to emotional eating during the midluteal phase of the cycle, when both estradiol and progesterone are relatively elevated (relative to the follicular phase). These effects are evident even after controlling for within-persons changes in negative affect across the cycle (Klump, Keel, Racine, et al., 2013), and a similar pattern has been observed for binge-eating frequency among women with clinically significant binge eating (Edler, Lipson, & Keel, 2007; Klump et al., 2014).
Although nearly all women are exposed to ovarian hormones, not all women experience binge eating or related forms of dysregulated eating (e.g., emotional eating). It is possible that only women who are already at elevated biological/genetic or psychosocial risk experience increased dysregulated eating during risky hormonal milieus. If so, the strength of associations between ovarian hormones and dysregulated eating would be expected to vary along with individual differences in these risk factors. To date, however, few moderators of ovarian hormone–dysregulated eating associations have been identified. Women with a history of clinically significant binge eating have shown stronger associations between ovarian hormone levels and emotional eating than women without a history of binge eating (Klump et al., 2014), but the source of these differences is unclear. Body mass index, dietary restraint, and impulsive personality dimensions (e.g., negative urgency) do not appear to moderate hormone–emotional eating associations across the menstrual cycle (Klump, Keel, Burt, et al., 2013; Racine et al., 2013b). Genetic influences on emotional eating vary across different hormonal milieus (Klump et al., 2015, 2016), indicating that individual differences in genetic risk play a role. However, specific genes or molecular pathways have yet to be identified. Additional research is therefore needed to uncover factors that predict the strength of ovarian hormone–dysregulated eating associations, which may help identify the women at greatest risk.
One promising but as yet unexplored potential moderator is the general intensity of negative affect (e.g., anger, sadness, fear) a woman experiences across situations. Women who engage in binge eating tend to experience higher trait negative affect and increased rates of disorders characterized by elevated negative affect (e.g., depression) than other women (e.g., Welch et al., 2016; Wolff, Crosby, Roberts, & Wittrock, 2000). Likewise, women who experience higher negative affect across time report more emotional eating than those who typically experience less negative affect (Racine et al., 2013a). Negative affect also correlates with other risk factors for dysregulated eating, such as stress and emotion regulation difficulties (Bolger, DeLongis, Kessler, & Schilling, 1989; Gross, 2015). This suggests that the general intensity of negative affect a woman experiences is a significant indicator of her overall risk for dysregulated eating.
Factors that make women more generally prone to intense negative affect (e.g., difficulties with emotion regulation, neurobiological vulnerabilities) could also potentially amplify the influence of ovarian hormones on binge eating and emotional eating. Persistent elevations in negative affect may therefore serve as a proxy for a range of underlying biological and/or psychosocial vulnerabilities that could render a woman more susceptible to dysregulated eating associated with shifts in ovarian hormones. Although a prior study failed to find a moderating effect of negative emotionality (i.e., a personality dimension encompassing emotional lability and general negative affect; Patrick, Curtin, & Tellegen, 2002) on hormone–emotional eating associations (Racine et al., 2013b), the inclusion of other traits in this construct (e.g., persecutory beliefs, antisocial tendencies) may have obscured the effect of negative affect per se.
Elucidating whether individual differences in negative affect moderate relationships between ovarian hormones and dysregulated eating could help researchers better understand why some women are more vulnerable to hormone effects than others. It would also significantly enhance understanding of the role of affective experience in risk for dysregulated eating. Although prior research has shown that within-persons fluctuations in negative affect are associated with within-persons changes in emotional eating and binge eating (Haedt-Matt et al., 2014; Haedt-Matt & Keel, 2011), it is also important to understand how between-persons differences in affective experience contribute to heightened risk across situations. In other words, although it is clear that daily fluctuations in negative affect predict increases in emotional eating and binge eating, it is not yet fully evident why women who are generally higher in negative affect experience more dysregulated eating. Identifying potential mechanisms underlying these individual differences (including the possibility of increased sensitivity to biological factors, e.g., changing hormone levels) is important to deepen the field’s understanding of etiology and risk. Although prior research on ovarian hormone–dysregulated eating associations has statistically controlled for within-persons changes in negative affect across the cycle (e.g., Klump, Keel, Racine, et al., 2013), it has yet to examine how trait negative affect may interact with ovarian hormones to influence risk.
The primary aim of the current study was therefore to examine whether ovarian hormone–emotional eating associations across the menstrual cycle differ as a function of individual differences in trait negative affect. Ovarian hormone–emotional eating associations were expected to be stronger among women with higher trait negative affect. A secondary aim was to investigate whether these moderating effects are similar across the full spectrum of binge eating-related pathology. To do so, the moderating effects of trait negative affect on ovarian hormone–emotional eating associations were first compared between women with and without clinically significant binge eating. Moderation of associations between ovarian hormone and binge-eating frequency was then examined in the subsample of women with a history of clinically significant binge eating.
Method
Participants
Analyses were conducted with data from 446 female twins (age range = 15–24 years; M = 17.84, SD = 1.79) from the Twin Study of Hormones and Behavior Across the Menstrual Cycle (Klump, Keel, Racine, et al., 2013). The current sample includes participants who were also included in prior reports analyzing data from the parent study (43.9% were included in the first report of the initial study participants—before completion of data collection—in Klump, Keel, Racine, et al., 2013; 99.8% were included in Klump et al., 2014). Participants were recruited through the Michigan State University Twin Registry, which identifies twins through birth records using methods described previously (Burt & Klump, 2013, 2019; Klump & Burt, 2006). Twins had to meet the following eligibility criteria to be included in the study: (a) menstruation every 22 to 32 days for the past 6 months, (b) no hormonal contraceptive use in the past 3 months, (c) no psychotropic or steroid medications in the past 4 weeks, (d) no pregnancy or lactation in the past 6 months, and (e) no history of genetic or medical conditions known to influence hormone functioning or appetite/weight (Klump, Keel, Racine, et al., 2013).
This sample was demographically representative of the recruitment region with regard to race and ethnicity (Klump, Keel, Racine, et al., 2013). The majority of participants identified as White (n = 368; 82.5%), and smaller numbers identified as Black or African American (n = 51; 11.4%), Asian American (n = 2; 0.4%), American Indian or Alaska Native (n = 2; 0.4%), or more than one race (n = 22; 4.9%). The remaining participant (n = 1; 0.2%) did not identify her race. In addition, 9.4% (n = 42) of participants across races identified as Latina. The average participant body mass index (BMI) was 23.69 (SD = 5.47, range = 15.30–47.59). With respect to zygosity, 56.5% (n = 252) of twins were monozygotic, and 43.5% (n = 194) were dizygotic.
Procedure
As described by Klump, Keel, Racine, et al. (2013), participants provided daily saliva samples for ovarian hormone measurements and ratings of affect, emotional eating, and binge-eating frequency for 45 consecutive days. Saliva samples were collected in the morning, whereas affect, emotional eating, and binge-eating ratings were acquired in the evening, ensuring that hormone measures for a given day preceded behavioral ratings. Three additional in-person assessments were conducted at the beginning, midpoint, and end of data collection. At each of these assessments, saliva samples were collected, BMI was measured, and study eligibility was reconfirmed. Between assessments, staff contacted participants once per week to answer questions and confirm protocol adherence. Dropout over the course of the study (3%) and missing data (≤ 6%) were minimal, and only a small number (3%) of participants became ineligible because of pregnancy or medication use (Klump, Keel, Racine, et al., 2013).
Measures
Emotional eating
Daily emotional eating was assessed using a version of the Emotional Eating scale from the Dutch Eating Behavior Questionnaire (DEBQ; van Strien, Frijters, Bergers, & Defares, 1986). Although the Emotional Eating scale of the DEBQ is intended to refer to emotional eating in general, we modified the scale so that it referred to emotional eating on the day the participant completed the questionnaire. We made this modification with the permission of van Strien et al. Participants rated how often 13 items assessing the desire to eat in response to negative emotions (e.g., “Did you have the desire to eat when you were irritated?”) were true for them on that day on a scale from 1 (not at all) to 5 (very often). This version of the DEBQ Emotional Eating scale has been used in past studies investigating ovarian hormone-emotional eating associations in community samples (Klump, Keel, Culbert, & Edler, 2008) and correlates strongly with measures of binge eating (e.g., the Bulimia scale of the Eating Disorders Inventory; van Strien, 1996) and palatable food consumption in laboratory settings (van Strien, 2000). It has also been shown to differentiate between obese women with and without binge-eating disorder (Schulz & Laessle, 2010), which suggests a specific association with binge-eating pathology.
Lifetime history of binge eating
History of binge-eating episodes (BEs; representing current or past clinically significant binge eating) was assessed via interview using a version of the Eating Disorders Module from the Structured Clinical Interview for DSM-IV Axis 1 Disorders (SCID; First, Spitzer, Gibbon, & Williams, 1996) modified to include additional symptom probes tailored to a community sample. There was good interrater reliability for the presence of BEs (κ = .82; Klump et al., 2014).
Twenty-seven women (6.1% of the sample) reported current or past BEs on the SCID. To maximize power with a community sample and maintain consistency with prior studies (Klump et al., 2014), this includes both participants with threshold objective binge-eating episodes (n = 15; ≥ 1,000 calories consumed and both behavioral (e.g., eating more than intended, being unable to stop eating, going back repeatedly to find more food) and psychological (e.g., feeling sad, guilty, or disgusted with oneself after binge eating) indicators of severe loss of control) as well as participants who may have consumed somewhat fewer calories (i.e., 600–999 calories) or reported less severe loss of control during BEs (i.e., endorsed some but not all behavioral indicators of loss of control along with psychological indicators). This approach is consistent with evidence that loss of control over eating is clinically significant even if the amount of food consumed falls short of “objectively large” thresholds (i.e., ≥ 1,000 calories; Forney, Haedt-Matt, & Keel, 2014). In analyses, the presence of BEs was identified using a dichotomous indicator (0 = no BEs; 1 = current or past BEs).
Daily binge eating
Participants also reported the number of times they engaged in BEs (0–9 or more episodes) each day during the 45 days of the study. To ensure that participants provided valid reports of BEs over the course of the study, they were given a detailed definition of a BE during the first study session and quizzed on their understanding of BEs with four case examples (for additional details, see Klump et al., 2014). These steps were shown to increase the accuracy of self-reported BE episodes in prior research (Celio, Wilfley, Crow, Mitchell, & Walsh, 2004). Although daily BE data were collected for all participants, they were analyzed only in participants with a history of BEs confirmed through the SCID (described above) to provide the highest likelihood that reported BE episodes aligned with our study definition and represented clinically significant BE. This approach is consistent with our previous article examining daily binge eating in this sample (Klump et al., 2014).
Ovarian hormones
Estradiol and progesterone levels were assayed using saliva samples, which promote greater compliance and show stronger hormone-behavior associations than more invasive methods (e.g., bloodspots; Edler et al., 2007). Salivary samples were analyzed for hormone levels using specialized enzyme immunoassay kits from Salimetrics, LLC (State College, PA) that show excellent reliability (intra- and interassay coefficients of variation: estradiol = 7.1% and 7.5%; progesterone = 6.2% and 7.6%), specificity (determined by interpolating the mean optical density −2 SD of 10–20 replicates at the 0 pg/ml level; estradiol = .10 pg/ml; progesterone = 5 pg/ml), and method accuracy (measured via spike recovery and linearity; estradiol = 104.2% and 99.4%; progesterone = 99.6% and 91.8%; Klump, Keel, Racine, et al., 2013). To optimize use of resources, samples were assayed daily during key periods of hormonal change (i.e., the midfollicular through premenstrual phases) and every other day when hormone levels were expected to be low and stable (i.e., during menstrual bleeding and the early follicular phase).
Menstrual phases
Trained raters coded menstrual phase on the basis of estradiol and progesterone hormone plots and recorded days of menstrual bleeding. Two raters coded menstrual phases for each participant, and discrepancies were resolved at weekly meetings (for more detailed descriptions of the coding methods, see Klump et al., 2015). In analyses, phase is identified using a dichotomous indicator (0 = before ovulation; 1 = after ovulation). Observations were grouped into before and after ovulation rather than more finely segmented phases of the menstrual cycle to conserve power to detect interaction effects.
Negative affect
Daily negative affect was measured using the negative emotion items from the Positive and Negative Affect Schedule (PANAS; Watson, Clark, & Tellegen, 1988). Participants rated the extent to which they experienced 10 negative emotions (distressed, upset, guilty, scared, hostile, irritable, ashamed, nervous, jittery, and afraid) each day on a scale from 1 (very slightly or not at all) to 5 (extremely). Daily negative affect composites were created by summing across negative emotion ratings on a given day.
As in a prior study of trait negative affect in this sample (Racine et al., 2013a), trait negative affect was calculated by averaging the daily negative affect composite scores for each woman over the study. Research using other brief affect scales found that negative affect is relatively stable across time (r = .60 for global ratings of negative affect across 2 years; Hudson, Lucas, & Donnellan, 2017), which suggests that negative affect measured over a discrete period provides a reasonable indication of trait affect. With respect to convergent validity, prior research found significant correlations between trait negative affect derived from daily ratings on the PANAS and measures of stress, anxiety, depression, and self-esteem (inversely) in nonclinical participants (rs = .23–.51; Merz & Roesch, 2011). In our own sample, we found that trait negative affect was significantly correlated with negative emotionality on the Multidimensional Personality Questionnaire-Brief Form (r = .39, p < .001; Patrick et al., 2002) and was significantly higher in participants with a history of major depressive disorder on the SCID (17.03 vs. 14.65, p = .001). An average of 41.97 days of PANAS ratings per participant were available to compute trait negative affect (range = 26–45 days), providing a strong indication of participants’ negative affect over time.
Statistical analyses
Data preparation for time-varying predictors
As described by Klump, Keel, Racine, et al. (2013), 5-day rolling averages were calculated for both estradiol and progesterone. Rolling averages for ovarian hormones are standard in the field (Kassam et al., 1996; Waller et al., 1998) because they reduce “noise” introduced through intermittent hormone release (Gladis & Walsh, 1987) and help to account for potential delays in hormone effects on behavior (Eckel, 2011).
As in past studies of ovarian hormone–dysregulated eating associations (Klump et al., 2014; Klump, Keel, Burt, et al., 2013; Klump, Keel, Racine, 2013; Racine et al., 2013b), all time-varying measures were standardized within persons (i.e., the difference between each observation and a woman’s mean was divided by the standard deviation of all observations for that woman). This allowed for modeling of how changes in hormone levels from a woman’s average predicted changes in emotional eating and binge eating relative to her average. We standardized the data within persons (vs. person mean centering without standardization) because trait negative affect was correlated with average levels of emotional eating; thus, the same qualitative change in emotional eating across the cycle (e.g., a 50% increase) would lead to a greater raw change in emotional eating scores among women with high trait negative affect. Standardizing emotional eating within persons allowed us to examine whether there was a qualitative difference in effects after accounting for expected differences in the absolute magnitude of changes across the cycle. More complex methods of centering the data (e.g., person-specific time trend centering) were not necessary because there was little evidence of meaningful linear time trends in any time-varying predictor (Curran & Bauer, 2011).
Emotional eating scores were nonnormally distributed even after within-persons standardization. Log-transforming values before within-persons standardizing did not meaningfully improve normality (kurtosis = 8.29 for the transformed values vs. 8.82 for the untransformed values), and thus untransformed values were retained in analyses. Simulation studies have shown that estimates and standard errors for fixed effects in mixed linear models are very robust to violations of normality, particularly when sample sizes are large (e.g., 50 or more; Jacqmin-Gadda, Sibillot, Proust, Molina, & Thiébaut, 2007; Maas & Hox, 2004). It is therefore unlikely that nonnormality in emotional eating scores would have affected the inferences drawn from our analyses.
Data preparation for trait-level predictors
Trait negative affect was standardized across all participants in the sample (for analyses involving the full sample) or a given subsample (for analyses that examined effects separately in women with and without BEs). In other words, the difference between a woman’s value and the mean value for all participants in the sample/subsample was divided by the standard deviation across participants. This provided a measure of a woman’s trait negative affect relative to other women.
Missing data
As described by Klump, Keel, Racine, et al. (2013), in the case of missing data for PANAS or DEBQ Emotional Eating ratings, raw scores were prorated if no more than 10% of items were missing and marked as missing otherwise. Rolling averages were calculated if there were at least 3 days of data within the 5-day window and counted as missing if there were fewer than 3 days of data. Observations from anovulatory cycles were excluded from analyses, which is in line with past studies (Klump et al., 2015; Klump et al., 2016) and evidence that changes in eating associated with ovarian hormone fluctuations are altered during anovulatory cycles (Asarian & Geary, 2006). Five women (1.1% of the sample) had missing data for presence of BEs on the SCID and so were excluded from analyses involving BE status.
Statistical models
Overall modeling approach
All statistical analyses were conducted using Stata (Version 15; StataCorp, 2017). Mixed linear models (MLMs) were used for analyses given the nonindependence of the repeated measures twin data. Models involving the full sample used maximum likelihood estimation (MLE), whereas models involving only the subset of women with BEs used restricted maximum likelihood (REML) to account for small sample size. Three-level models were used. Observations (Level 1) were nested within participants (Level 2), and participants were nested within twin pairs (Level 3). A random slope was placed on all time-varying predictors at the participant level. Covariance structures for the random effects and residuals were determined by optimizing model fit measures (e.g., Akaike information criterion [AIC]; Bayesian information criterion [BIC]) to the extent possible while maintaining convergence; final models used a diagonal covariance structure for random effects and a Toeplitz structure for residuals to account for autocorrelation (with the correlation between residuals set to 0 after a lag of 5 days).
It is theoretically possible that women who generally experience more intense negative affect also experience more variability in negative affect across the cycle, confounding between-persons and within-persons effects. Daily negative affect was therefore included as a covariate in all models to account for this possibility and maintain consistency with prior studies (Klump et al., 2014; Klump, Keel, Racine, et al., 2013). Note that daily within-persons standardized negative affect is by definition linearly independent from a woman’s trait negative affect (i.e., the correlation between them is 0). This is true because the mean of every participant’s within-persons standardized daily negative affect across the study is 0 regardless of her absolute average level of negative affect.
Note that all models represent cross-level interactions, in which a factor that differs among people (i.e., trait negative affect) modifies the strength of within-persons associations (e.g., between ovarian hormone levels and emotional eating on a given day). In these models, the main effect of trait negative affect can be conceptualized as an “adjustment factor” on the intercept (i.e., it modifies the prediction of a woman’s level of emotional eating relative to her average when her levels of estradiol, progesterone, and daily negative affect are at their average). It does not represent the strength of the association between trait negative affect and average level of emotional eating, which is best captured by the correlation between these variables (presented in descriptive statistics below).
We set our threshold for statistical significance to p < .01 for all analyses to account for multiple comparisons.
Associations between menstrual phase and emotional eating
We first examined whether trait negative affect moderated menstrual phase–emotional eating associations. Menstrual phase was analyzed in addition to ovarian hormone levels to allow for comparisons with previous studies that have examined menstrual phase alone (e.g., Lester, Keel, & Lipson, 2003) or both menstrual phase and hormones (e.g., Klump et al., 2014). In these analyses, emotional eating was predicted from menstrual phase (i.e., before or after ovulation), trait negative affect, and the interaction between menstrual phase and trait negative affect.
Associations between ovarian hormones and emotional eating
We then examined whether trait negative affect moderated associations between fluctuations in ovarian hormones themselves (i.e., within-persons standardized estradiol and progesterone) and emotional eating. Emotional eating was initially predicted from estradiol, progesterone, trait negative affect, the Estradiol × Trait Negative Affect interaction, and the Progesterone × Trait Negative Affect interaction. Results are also reported for models including the Estradiol × Progesterone interaction and three-way Estradiol × Progesterone × Trait Negative Affect interaction.
Differences across binge-eating status
Next, we examined whether the moderating effects of trait negative affect on ovarian hormone–emotional eating associations differed between women with and without BEs (i.e., a double moderation model). We approached this question in two steps. First, we modeled three-way interactions between menstrual phase, BE status, and trait negative affect. We then modeled three-way interactions between BE status, trait negative affect, and estradiol/progesterone. The Estradiol × Progesterone interaction and higher order interactions involving this term were not included in this model so that a four-way interaction was not required. If analyses in the full sample indicated a significant Estradiol × Progesterone × Trait Negative Affect interaction, we modeled this interaction in women with and without BEs separately to explore whether effect sizes differed between groups. Finally, we tested whether trait negative affect moderated menstrual phase or ovarian hormone influences on daily binge-eating frequency only in the subsample of women with BEs. These analyses were identical to those previously described for emotional eating in the full sample but with daily binge-eating frequency as the outcome.
Comparison with past statistical modeling approaches
The statistical modeling approach used in the current analyses differed somewhat from that used in prior research (e.g., Klump, Keel, Racine, et al., 2013). Because of advances in MLM techniques, we were able to explicitly model autocorrelation in the current analyses. Although our group has used rolling averages for emotional eating/binge eating in some past analyses (Klump et al., 2014; Klump, Keel, Racine, et al., 2013), while constructing an updated modeling approach that accounted for autocorrelation, we realized that using rolling averages for the dependent variables induced an undesirable level of autocorrelation. More recent research has also shown that daily affect is most strongly associated with emotional eating on the same day (Haedt-Matt et al., 2014), indicating that rolling averages were not needed to account for delayed effects of negative affect on emotional eating. Single-day values were therefore used for emotional eating, binge eating, and negative affect in all models. We also chose not to include BMI as a covariate given minimal changes across the study period (see Klump et al., 2014; Klump, Keel, Racine, et al., 2013) and minimal effects on results (for models including BMI, see Tables S3–S5 in the Supplemental Material available online).
Although it is not possible to empirically examine the impact of all of changes on model fit (because models are not completely nested), we observed that including a Toeplitz residual structure to account for autocorrelation improved fit to the data relative to the same model without this structure (e.g., for the model examining phase in the full sample, AIC improved from 41,750.61 to 40,946.47, and BIC improved from 41,788.72 to 40,984.58; for the model examining estradiol and progesterone separately in the full sample, AIC improved from 33,372.61 to 32,968.78, and BIC improved from 33,469.11 to 33,102.39). As shown in Table S1 in the Supplemental Material, ovarian hormone-emotional eating associations were broadly consistent across statistical modeling approaches, although the models used in the current article might produce slightly more conservative estimates than models used in prior articles.
Results
Descriptive statistics
Descriptive statistics for person-level and daily variables are presented in Table 1. Both raw scores and z scores are included to provide an indication of variability across the sample and (for daily variables) within each participant over the study. A range of trait negative affect was represented in the sample (M = 14.95; range = 10.04–30.49; possible range = 10–50), and nearly the full possible range of daily negative affect was reported (range = 10–46; possible range = 10–50). The distribution of negative affect aggregated across days in this sample was comparable with that previously observed in other nonclinical samples (e.g., Watson & Clark, 1999). There was also good variability in daily emotional eating scores (range = 1–4.69; possible range = 1–5). Similar to prior research, higher trait negative affect was associated with more emotional eating (r = .47, p < .001) and a greater likelihood of having current or past BEs (OR = 1.95, p = .002). In contrast, trait negative affect was unrelated to average levels of estradiol (r < .001, p = .998) or progesterone (r = −.01, p = .824). As has been observed in prior research (e.g., Klump et al., 2016; Klump et al., 2018), average levels of emotional eating were also not significantly related to average estradiol (r = −.02, p = .661) or progesterone (r = −.05, p = .292) levels; however, this does not preclude within-persons associations between fluctuations in hormone levels and fluctuations in emotional eating from day to day that have been found in previous work (e.g., Klump et al., 2014; Klump, Keel, Burt, et al., 2013; Klump, Keel, Racine, et al., 2013; Racine et al., 2013b).
Descriptive Statistics for Person-Level and Daily Variables
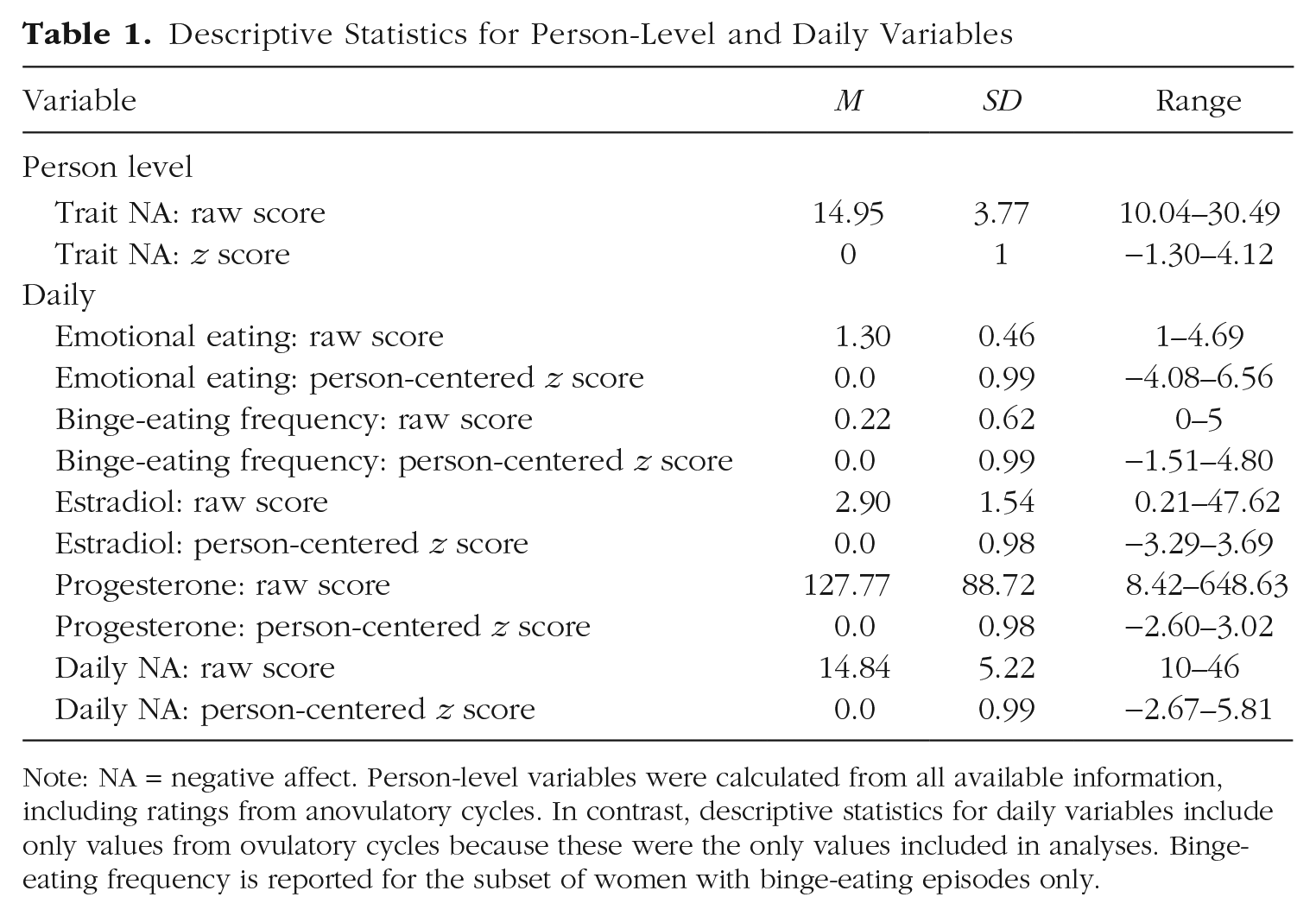
Note: NA = negative affect. Person-level variables were calculated from all available information, including ratings from anovulatory cycles. In contrast, descriptive statistics for daily variables include only values from ovulatory cycles because these were the only values included in analyses. Binge-eating frequency is reported for the subset of women with binge-eating episodes only.
Trait negative affect as a moderator of menstrual phase-emotional eating associations
As shown in Table 2, people with higher trait negative affect tended to show a stronger association between menstrual phase and daily emotional eating, although this relationship did not quite reach our threshold for statistical significance (β = 0.05, p = .017). The overall pattern indicated that the postovulatory period was riskier for emotional eating in women with higher trait negative affect. A simple-slopes analysis was conducted to examine effects in women 1 SD above the mean, at the mean, and 1 SD below the mean in trait negative affect. Results indicated that the change in emotional eating from before ovulation to after ovulation was greater in women with high trait negative affect (β = 0.06, p = .026) than in women with moderate (β = 0.01, p = .446) or low (β = −0.03, p = .233) trait negative affect.
Interactions Between Trait Negative Affect and Menstrual Phase (Before vs. After Ovulation) or Estradiol/Progesterone in Predicting Emotional Eating in the Full Sample
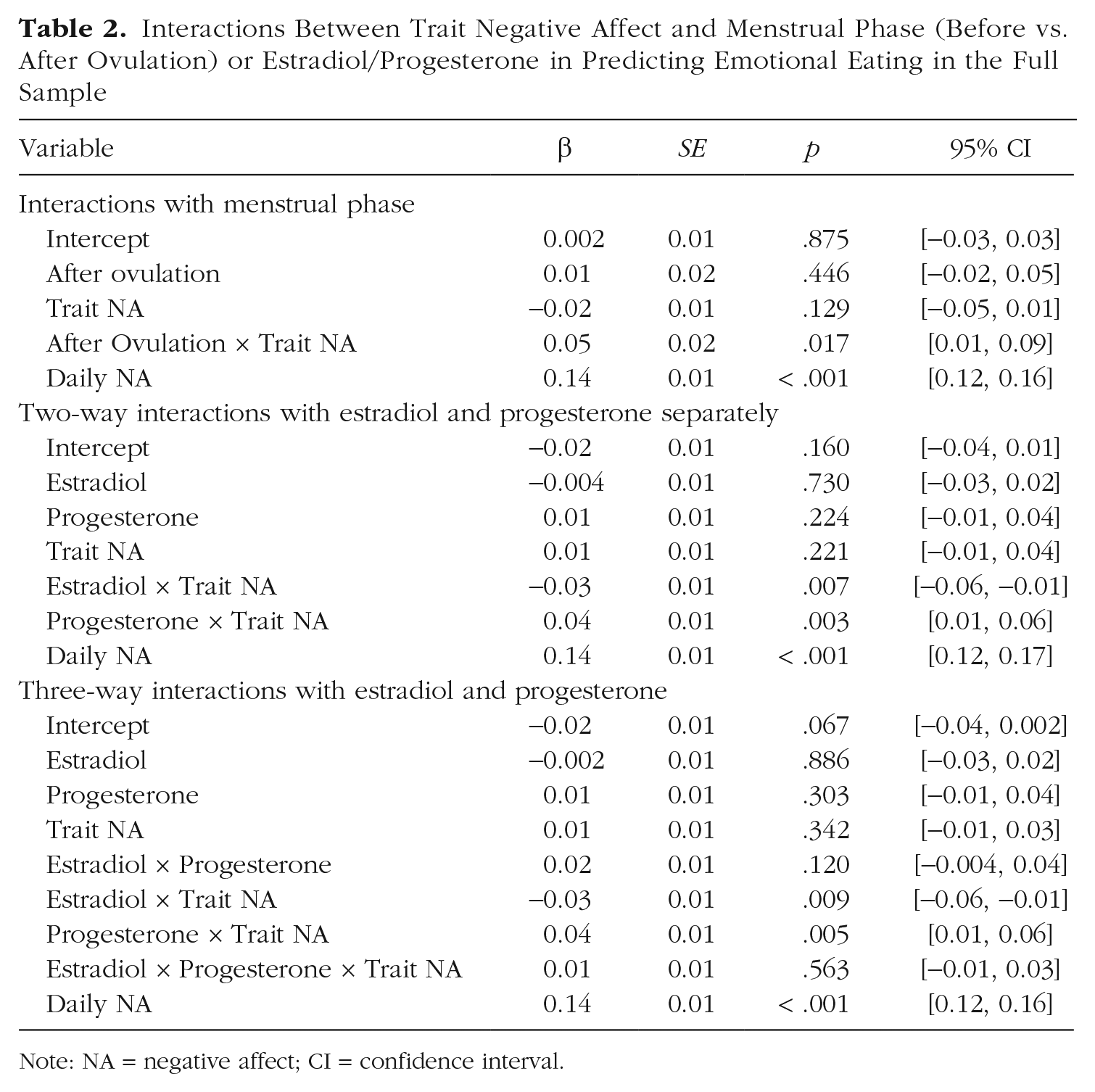
Note: NA = negative affect; CI = confidence interval.
Trait negative affect as a moderator of ovarian hormone–emotional eating associations
In the model with direct measures of ovarian hormones, trait negative affect significantly moderated associations between emotional eating and both estradiol (β = −0.03, p = .007) and progesterone (β = 0.04, p = .003; see Table 2 and Fig. S1 in the Supplemental Material). The pattern of moderation indicated that higher estradiol was more protective against and higher progesterone riskier for emotional eating in women with higher trait negative affect. No main effects of hormones were observed. In this context, the presence of significant interactions without significant hormone main effects indicates that the influence of ovarian hormones on emotional eating depends on a woman’s level of trait negative affect (rather than suggesting that hormones do not affect emotional eating). Indeed, simple-slopes analyses indicated that progesterone was significantly positively associated with emotional eating only for women high in trait negative affect (high trait negative affect: β = 0.05, p = .003; moderate trait negative affect: β = 0.01, p = .224; low trait negative affect: β = −0.02, p = .196). Likewise, estradiol showed the strongest (negative) association with emotional eating for women high in trait negative affect, with a trend toward the opposite effect in women with low trait negative affect (high trait negative affect: β = −0.04, p = .032; moderate trait negative affect: β = −0.004, p = .730; low trait negative affect: β = 0.03, p = .083).
When the Estradiol × Progesterone × Trait Negative Affect interaction was added to the model, it was nonsignificant (β = 0.01, p = .563), and the Estradiol × Trait Negative Affect and Progesterone × Trait Negative Affect interactions remained significant (see Table 2). These findings suggest that the strength of the interactions between trait negative affect and estradiol/progesterone do not depend on the level of the other hormone (e.g., the Estradiol × Trait Negative Affect interaction is equally strong when progesterone is high as when progesterone is low). To examine whether the lack of a significant three-way interaction might be due to use of a slightly different statistical modeling approach, we reran analyses examining trait negative affect as a moderator of hormone-emotional eating associations using the original Klump, Keel, Racine, et al. (2013) statistical approach. As shown in Tables S6 and S7 in the Supplemental Material, the Estradiol × Progesterone × Trait Negative Affect interaction was not significant in either approach.
Moreover, the overall pattern of effects was broadly consistent across statistical modeling approaches, with somewhat stronger interactions between trait negative affect and phase and somewhat weaker interactions between trait negative affect and direct measures of hormones, using the Klump, Keel, Racine, et al. (2013) approach. Note that the lack of a significant Estradiol × Progesterone interaction (independent of trait negative affect) using either approach may reflect the presence of higher-order interaction terms (i.e., interactions with trait negative affect) that were not present in prior studies and that alter the interpretation of lower-order effects.
Comparing the strength of trait negative affect-hormone interactions in women with and without binge eating
BE status did not significantly moderate the Estradiol × Trait Negative Affect interaction in predicting emotional eating (β = −0.01, p = .890, for the three-way Trait Negative Affect × Estradiol × BE Status interaction; see Table 3). This suggests that low estradiol is equally risky for emotional eating in women with high trait negative affect with and without a history of BEs. However, BE status did significantly moderate the association between progesterone levels and emotional eating (β = 0.14, p = .004 for the Trait Negative Affect × Progesterone × BE Status interaction), which likely contributed to a qualitatively stronger Trait Negative Affect × Phase interaction in women with BEs (β = 0.04 in women without BEs vs. 0.23 in women with BEs). Although the Progesterone × Trait Negative Affect interaction was in the same direction in both women with and without BEs, it was amplified among women with BEs (β = 0.03 for women without BEs and β = 0.16 for women with BEs; see Table S2 and Fig. S2 in the Supplemental Material). Simple-slopes analyses indicated that higher progesterone was associated with increased emotional eating in women with BEs who had high trait negative affect (β = 0.17, p = .003) but a trend toward decreased emotional eating in women with BEs who had low trait negative affect (β = −0.15, p = .017). Exploratory analyses showed that the Progesterone × Trait Negative Affect interaction remained significant if the sample was restricted to women with current BEs (n = 21; β = 0.14, p = .003) or threshold objective binge-eating episodes characterized by consumption of at least 1,000 calories and severe loss of control (n = 15; β = 0.14, p = .002). This suggests that results are robust and effects in the BE subsample were not simply due to higher negative affect among women with current BEs compared with women with past BEs only or different effects in women with more severe BEs.
Interactions Between Trait Negative Affect and Menstrual Phase (Before vs. After Ovulation) or Estradiol/Progesterone in Predicting Emotional Eating in the Full Sample, Moderated by BE Status
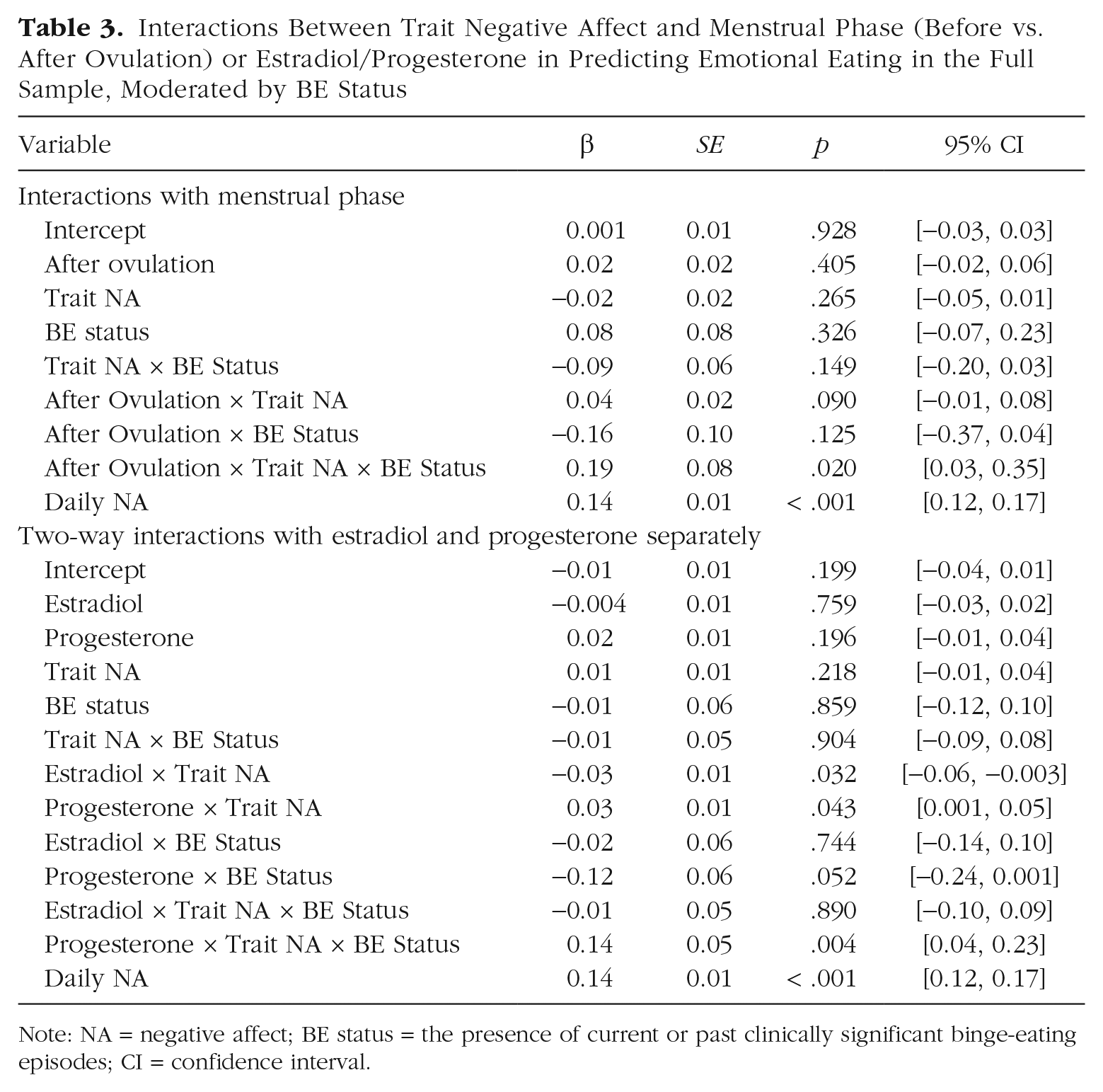
Note: NA = negative affect; BE status = the presence of current or past clinically significant binge-eating episodes; CI = confidence interval.
Trait negative affect as a moderator of hormone–binge eating associations in women with clinically significant binge eating
Daily negative affect was the only significant predictor of binge-eating frequency in the subsample of women with BEs (and this effect reached our threshold for statistical significance in only one model). The main effects of menstrual phase/ovarian hormones and the Menstrual Phase/Ovarian Hormone × Trait Negative Affect interactions were not significant. However, the pattern of moderation was in the same direction as previously described for emotional eating. Specifically, the postovulatory period (β = 0.11, p = .274) and higher progesterone levels (β = 0.06, p = .505) were riskier for binge eating in women with higher trait negative affect (see Table 4). Results were consistent in showing no significant Hormone × Trait Negative Affect interactions when binge frequency was examined using a count-based negative binomial regression model (see Table S8 in the Supplemental Material). The lack of significant effects may be due in part to reduced power (a particular concern with cross-level interaction models), given that only 16 (59.3%) participants in the BE subsample reported a binge frequency of 1 or greater during the study period. This possibility is consistent with the fact that the association between daily negative affect and daily binge frequency was only weakly significant despite similar effect sizes in models examining emotional eating.
Interactions Between Trait Negative Affect and Menstrual Phase (Before vs. After Ovulation) or Estradiol/Progesterone in Predicting Daily Binge-Eating Frequency in the BE Subsample Only
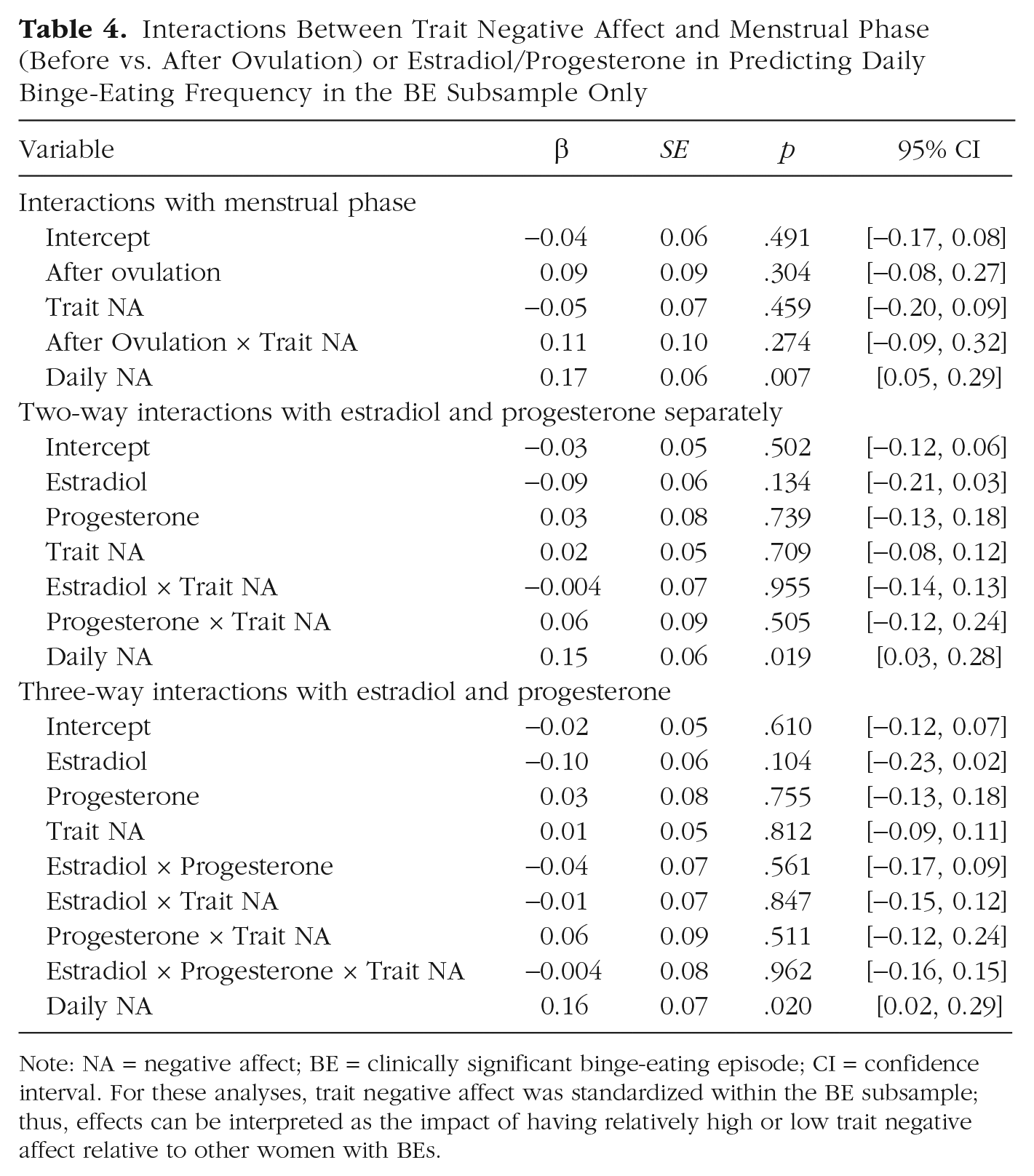
Note: NA = negative affect; BE = clinically significant binge-eating episode; CI = confidence interval. For these analyses, trait negative affect was standardized within the BE subsample; thus, effects can be interpreted as the impact of having relatively high or low trait negative affect relative to other women with BEs.
Exploratory analyses
To determine whether effects were specific to trait negative affect, analyses were repeated using a measure of trait positive affect calculated from the PANAS positive emotion items. There were no significant interactions between trait positive affect and menstrual phase or estradiol or progesterone (with or without the Estradiol × Progesterone interaction; ps > .05), which suggests a unique effect of trait negative affect.
We also wondered whether the impact of ovarian hormones on emotional eating might differ depending on a woman’s daily level of negative affect (i.e., if she were experiencing more negative affect than was typical for her on a given day independent of overall levels of negative affect). We therefore added interactions between daily negative affect and phase/ovarian hormones to our models. As shown in Tables S9 and S10 in the Supplemental Material, there were no significant Daily Negative Affect × Phase/Hormone interactions, and the magnitude of the Trait Negative Affect × Phase/Hormone interactions were largely unchanged.
Discussion
This study was the first to examine how affective and hormonal risk interact to influence dysregulated eating in women. The results indicate that individual differences in trait negative affect are an important moderator of hormone effects on emotional eating, particularly in women with a history of clinically significant binge eating. Findings may help to resolve discrepancies in prior research showing significant independent effects of ovarian hormones on dysregulated eating in women with bulimia nervosa (who would be expected to have elevated negative affect; Edler et al., 2007) but not in women from a community sample (Klump, Keel, Racine, et al., 2013). Overall, these findings advance the understanding of individual differences in ovarian hormone influences on eating behavior as well as the ability to predict who is at greatest risk for increases in dysregulated eating during risky hormonal milieus.
Although daily negative affect was the strongest independent predictor of dysregulated eating across the cycle, trait negative affect showed unique interactions with hormonal influences on emotional eating that were not present for daily negative affect or trait positive affect. In the full sample, women with high trait negative affect were at greatest risk for emotional eating when estradiol was low or progesterone was high, as occurs during the early follicular (i.e., low estradiol) and midluteal (i.e., high progesterone) phases of the menstrual cycle. The same overall pattern of effects was observed in women with and without BEs, although some effects were stronger in the BE subsample (see below). These findings suggest that hormonal factors may interact with risk factors for sustained high negative affect to promote increases in emotional eating. Note that women with high trait negative affect also had higher average levels of emotional eating (i.e., independent of hormone levels). Thus, women reported the highest absolute levels of emotional eating when they were high in trait negative affect and experienced a risky hormonal milieu characterized by low estradiol or high progesterone. These findings are broadly consistent with results from animal studies showing that female rats engage in the highest levels of binge eating when they have undergone ovariectomy (which removes the primary source of estradiol) and have been exposed to other sources of risk for dysregulated eating (i.e., food restriction and stress; Micioni Di Bonaventura et al., 2017).
One initially surprising finding was the lack of a three-way Estradiol × Progesterone × Trait Negative Affect interaction. Although initial data in a community sample showed that modulation of estradiol effects by progesterone was most influential in predicting emotional eating (Klump, Keel, Racine, et al., 2013), analyses in subpopulations have suggested that individual hormones may also be important for some women (particularly those with higher levels of eating pathology or who are at elevated risk). For example, estradiol on its own has been found to be a significant predictor of emotional eating in women with binge-eating episodes but not in women without binge eating (Klump et al., 2014). As mentioned above, estradiol and progesterone have also previously been found to be independently associated with binge eating in women with bulimia nervosa (Edler et al., 2007). These findings align with our current results showing stronger independent influences of ovarian hormones among women with greater trait negative affect and suggest that independent hormone effects may be evident only in populations with other vulnerability factors (e.g., a propensity for high negative affect) and could operate through separate neural pathways in these individuals.
The interaction between progesterone and trait negative affect in predicting emotional eating was particularly pronounced among women with a history of clinically significant binge eating. Indeed, the effect of menstrual phase on emotional eating was equivalent to that of daily negative affect in women with BEs and high trait negative affect (β = 0.26 for both predictors), who are arguably at highest risk for dysregulated eating. This suggests that the effect of simply being in the luteal phase of their menstrual cycle may be about as impactful as having an exceptionally bad day emotionally for these women. The presence of stronger hormone effects in the BE subsample in this study parallels results from a previous study that did not examine individual differences in negative affect (Klump et al., 2014) and suggests that women with a history of clinically significant binge eating may be particularly sensitive to ovarian hormone influences. Although prior studies across the menstrual cycle have suggested that progesterone increases emotional eating by counteracting the protective effects of estradiol (Klump, Keel, Racine, et al., 2013), findings from this study are consistent with recent evidence that progesterone may also strengthen the influence of psychosocial risk factors for disordered eating (e.g., weight-based teasing; Forney et al., 2019). The combination of sustained high negative affect and a within-persons peak in progesterone may therefore be a “perfect storm” for dysregulated eating in women with a history of BEs, who may also be more likely to have other sources of biological or behavioral risk (e.g., a history of dieting) that potentiate these effects. However, a close inspection of the Progesterone × Trait Negative Affect interaction in women with BEs (see Table S2 and Fig. S2 in the Supplemental Material) also shows that progesterone was negatively associated with emotional eating in a subset of women with a history of clinically significant binge eating but relatively low trait negative affect. This suggests that ovarian hormones could affect dysregulated eating differently for women whose binge eating may be driven primarily by factors other than negative affect, a finding that is consistent with prior research showing individual differences in ovarian hormone influences on binge eating in women with bulimia nervosa (Edler et al., 2007).
Mechanistically, trait negative affect may serve as a proxy for neurobiological or psychosocial risk factors that could amplify the influence of ovarian hormones on dysregulated eating. One possibility is that women who experience consistently high negative affect have underlying vulnerabilities in serotonergic or dopaminergic neural circuits that affect both mood and eating behavior and that these vulnerabilities become more pronounced during periods of hormonal risk. Estradiol has been shown to enhance dopaminergic and serotonergic signaling (Kuhn et al., 2010; McEwen & Alves, 1999; Morissette et al., 2008), whereas progesterone is associated with increased neural responsivity to negative stimuli (e.g., through increased amygdala reactivity; Andreano, Touroutoglou, Dickerson, & Barrett, 2018). Hormonal milieus characterized by low estradiol or high progesterone may therefore potentiate existing tendencies toward negative mood or emotion dysregulation, pushing women who are already at risk “over the edge” to engage in emotional eating. However, other possibilities also exist; for example, high trait negative affect could indicate the presence of chronic environmental stressors rather than biological vulnerabilities innate to an individual. Research incorporating neuroimaging and psychophysiological data or additional self-report measures of constructs such as emotion regulation and stress are therefore needed to determine the key mechanisms underlying the effects observed in this study.
Some limitations of this study should be noted. Because there is likely a reciprocal relationship between negative affect and emotional eating at the daily level (e.g., Haedt-Matt & Keel, 2011), it is conceivable that greater increases in emotional eating during the luteal phase could have driven higher average levels of negative affect. We examined this possibility and found that negative affect was very stable across the cycle in terms of average levels (follicular phase = 14.87; luteal phase = 14.81; p = .432) and individual differences (r = .90 between negative affect during the follicular phase and luteal phase for a given woman), making it unlikely that our findings were confounded by increases in emotional eating during risky phases of the cycle. Nonetheless, given our study design, it is difficult to completely disentangle the effects of emotional eating and negative affect. Additional studies that can better tease apart the temporal relationship between negative affect and emotional eating (e.g., examining these constructs multiple times per day) are therefore needed.
In addition, although a subset of women reported a history of clinically significant binge eating, this group was relatively small (n = 27), and it was not possible to examine potential differences across eating disorder diagnoses (i.e., bulimia nervosa vs. binge-eating disorder). Some caution is also warranted in interpreting interaction effects in this subsample because of small sample size. Replication in larger samples of women with clinically significant binge eating is needed, although it is worth noting that the direction of effects was consistent across women with and without BEs. Although the sample was demographically representative of the recruitment region, it had limited representation of women from some racial-ethnic groups (e.g., Asian/Asian American and Native American women); thus, it will be important to ensure results replicate in more diverse samples. As we advocated for in past work (e.g., Klump et al., 2017), replication of hormone effects and interactions by independent groups is also vital to ensure that findings are consistent across settings.
Finally, although the PANAS is a well-validated and widely used measure of negative and positive affect, it primarily captures high arousal emotions. Alternative measures of negative/positive affect that incorporate more low arousal emotions (e.g., bored, calm) may show different patterns of moderation. It would also be ideal to measure negative affect across a longer time span to ensure that a “trait-like” construct is being captured.
In conclusion, this study provides initial evidence that individual differences in negative affect moderate hormonal risk for emotional eating. High trait negative affect may not only be a risk factor for dysregulated eating in itself but may also be associated with stronger ovarian hormone influences on emotional eating. Among women with a history of clinically significant binge eating, the combination of sustained high negative affect and a within-persons peak in progesterone may be associated with particularly high risk for dysregulated eating. These findings are important given the high comorbidity rates between disorders characterized by binge eating and those characterized by elevated negative affect (e.g., mood and anxiety disorders, Ulfvebrand, Birgegård, Norring, Högdahl, & von Hausswolff-Juhlin, 2015). Clinicians should be aware that women with persistently high negative affect (e.g., those with depression) and a current or past eating disorder characterized by binge eating may be at elevated risk for dysregulated eating during the midluteal phase of their menstrual cycle (when progesterone levels are naturally higher). Additional research is needed to identify the biological and psychosocial factors that may underlie these effects.
Supplemental Material
Mikhail_Supplemental_Material – Supplemental material for Trait Negative Affect Interacts With Ovarian Hormones to Predict Risk for Emotional Eating
Supplemental material, Mikhail_Supplemental_Material for Trait Negative Affect Interacts With Ovarian Hormones to Predict Risk for Emotional Eating by Megan E. Mikhail, Pamela K. Keel, S. Alexandra Burt, Cheryl L. Sisk, Michael Neale, Steven Boker and Kelly L. Klump in Clinical Psychological Science
Footnotes
Transparency
Action Editor: Stephen Wonderlich
Editor: Kenneth J. Sher
Author Contributions
M. E. Mikhail and K. L. Klump developed the study concept. All of the authors contributed to the study design. Testing and data collection were performed by K. L. Klump. Data analysis and interpretation were performed by M. E. Mikhail under the supervision of K. L. Klump. M. E. Mikhail drafted the manuscript, and all of the authors provided feedback and approved the final manuscript for submission.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.