
Abstract
Background
The hallmark pathologies of neurodegenerative diseases include neuroinflammation, oxidative stress, mitochondrial dysfunction, protein misfolding, and dysregulated cell-survival signaling. These biochemical processes lead to neuronal death in disorders such as Alzheimer's disease, Parkinson's disease, and others. The complexities involved in neuronal cell death still need to be unraveled.
Objective
To study the role of curcumin and its mono- and diglucoside derivatives in IMR32 cells, focused on cytotoxicity, cell cycle analysis, apoptosis, reactive oxygen species (ROS), antioxidant potential, 8-hydroxy-guanosine (8-OH-G)* production, and cell and nuclear morphology.
Methods
In the present study, we synthesized a novel molecule, curcumin mono- and diglucoside, to improve the water solubility and partition coefficient, making the molecule highly bioavailable. We assessed cytotoxicity using the MTT assay and trypan blue, with a Nikon Eclipse E200 microscope. cell cycle and apoptosis by FACS, cell morphology by a Nikon microscope, nuclear morphology by confocal imaging, and 8-OH-G*, ROS, and antioxidant activity by ELISA kits.
Results
Our findings show that (1) curcumin and the positive control compound doxorubicin have lower IC50 values than curcumin monoglucoside and diglucoside; (2) The three compounds induce distinct cell-cycle arrests (curcumin: S-phase; curcumin-monoglucoside: G0–G1; curcumin diglucoside: G2). Curc-glucoside also reduced ROS production and DNA damage marker 8-OH-G*, while activating antioxidant potential; (3) Curcumin mainly causes necrosis; Curcumin mono- and diglucosides did not induce any apoptosis or necrosis; and doxorubicin promotes apoptosis only.
Conclusions
Curcumin mono- and diglucoside efficiently reduced ROS, increased antioxidant activity, and decreased 8-OH-G* production. Curcumin is more prooxidant and proapoptotic, whereas glycosylated derivatives are more antioxidant and cytostatic.
Keywords
Introduction
Neurodegenerative diseases and central nervous system (CNS) injuries share hallmark pathologies, including neuroinflammation, oxidative stress, mitochondrial dysfunction, protein misfolding, and dysregulated cell-survival signaling. These events collectively lead to neuronal death across disorders, namely Alzheimer's disease, Parkinson's disease, and trauma-induced brain injury.1–3 There are limited drugs to treat or manage many of these disorders, and future therapies will require novel drug targets. Polyphenols such as curcumin (diferuloylmethane), a bioactive component of Curcuma longa, have attracted preclinical interest due to their multiple pharmacological activities, including antioxidant and anti-inflammatory effects, modulation of proteostasis, and regulation of cell signaling pathways involved in neuronal survival and/or cell death. Several studies have indicated curcumin's multimodal neuroprotective effects and its capacity to modulate neuroinflammatory cascades and redox balance in CNS models.4–8
Curcumin, with low water solubility, limited blood-brain barrier permeability, and poor cellular uptake, posed challenges for further development as a drug. Alternatively, the preparation of different curcumin derivatives has attracted attention. Glycosylation (the attachment of glucose or oligosaccharides) is one such curcumin derivatization that has gained biological importance because it markedly increases aqueous solubility, improves chemical stability, and modulates cellular uptake and metabolic fate (Figure 1). Glucosylation also tends to reduce susceptibility to rapid phase II conjugation (glucuronidation/sulfation) at the phenolic sites. Still, it may introduce susceptibility to β-glucosidases in the gut or tissues (prodrug behavior).6–8

Structures of curcumin and its novel derivatives.
Beyond neuroprotection, curcumin and its derivatives exhibit concentration-dependent cytotoxic and anti-proliferative effects against neural tumor cell lines (neuroblastoma) by targeting cell-cycle regulators, mitochondrial pathways, and apoptotic machinery. In neuroblastoma models, curcumin triggers cell-cycle arrest and apoptosis by downregulating pro-survival pathways (e.g., PI3 K/AKT, NF-κB), upregulating the PTEN/p53 axis, and activating caspases, with concomitant changes in Bcl-2 family proteins and mitochondrial membrane potential. These mechanisms provide insight into the use of human neuroblastoma lines as experimental proxies for neuronal responses to stress and pharmacological agents.9–10 Further, curcumin glucosides are derivatives of curcumin, a compound found in turmeric, in which one or more sugar groups, for example, glucose, are chemically attached to the molecule. These modifications have been extensively investigated due to the poor water solubility, low absorption in the digestive system, rapid breakdown in the body, and limited overall bioavailability (amount available for use by the body) of native curcumin. The addition of sugar groups improves water solubility, chemical stability, and the ability of cells to absorb curcumin, positioning curcumin glucosides as strong candidates for use in medicines and nutritional supplements. Curcumin glucosides can be made using chemical reactions, special enzymes (glycosyltransferases), or natural processes carried out by certain microorganisms. Enzymatic glycosylation, which uses these enzymes, is especially useful for producing molecules with one or two glucose units, thereby improving water solubility. These types of curcumin glucosides often act as prodrugs—compounds given in an inactive or less active form, which release the active ingredient, curcumin, after being broken down by enzymes in the body, especially in tissues or the digestive system. Laboratory (in vitro) studies show that curcumin glucosides maintain antioxidant, anti-inflammatory, and anticancer activity while providing improved pharmacokinetics, meaning the way the body absorbs, distributes, and eliminates the compound. Researchers have found that these derivatives scavenge free radicals more effectively, are more stable under conditions found in the body, and are taken up by cells more efficiently than regular curcumin. Studies in animals show higher blood levels and longer circulation times for these sugar-attached derivatives. Recent research has focused on formulations, such as nanoparticles (tiny particles), liposomes (tiny fat-like bubbles), and phytosomal systems (plant-based delivery systems), to boost the oral uptake and effectiveness of curcumin glucosides as treatments. These methods are intended to overcome the challenges of native curcumin and to enhance its potential to manage inflammatory, nerve-related, metabolic diseases, and cancer11–14
The human neuroblastoma cell line IMR-32 is widely used as an in vitro model to study neuronal phenotypes and neuroprotective responses. In this study, we investigated the effects of curcumin and its derivatives (curcumin mono- and diglucoside) on IMR-32 neuroblastoma cells, with an emphasis on cell and nuclear morphology, MTT assay, trypan blue, cell cycle, reactive oxygen species (ROS), antioxidant activity, 8-hydroxy-guanosine (8-OH-G)* levels, and apoptosis.
Methods
Synthesis of curcumin mono- and diglucosides as cited earlier
In summary, curcumin mono- and diglucosides are produced through a two-step process involving phase-transfer-assisted glycosylation followed by deacetylation. Initially, curcumin is converted to its potassium salt and reacts with -D-acetobromoglucose in the presence of benzyltributylammonium chloride under ultrasound irradiation, yielding mono- and di- glucoside tetraacetates. Thin-layer chromatography (TLC) revealed two products (Rf values of 0.75 and 0.60), and electrospray ionization mass spectrometry (ESI-MS) confirmed molecular ions at m/z 721.34 [M + Na] and 1051.48 [M + Na], corresponding to the mono- and diglucoside tetracetates, respectively, with an overall yield of approximately 65%. In the second phase, deacetylation is performed with sodium methoxide in dry methanol, yielding the free curcumin mono- and diglucosides, which are validated by ESI-MS, showing molecular ions at m/z 553.28 ([M + Na]+) and 715. 35 ([M + Na]+), respectively. The final products are produced in a 0.6:1 ratio, with about 70% yield, are further purified by HPLC, and are subsequently used for experimental studies. 15
Bioavailability studies
IMR 32 cells are exposed to curcumin and its derivatives for 24 h, and the presence of curcumin and derivatives is estimated by liquid chromatography–mass spectrometry (LC-MS, Agilent model). In brief, cells are exposed to curcumin and its derivatives for 24 h. Cells are trypsinized and centrifuged (e.g., at 3000 rpm for 5 min). Cell pellets are lysed with cold methanol or acetonitrile (protein-precipitation method). Samples are vortexed and sonicated to ensure complete extraction. Lysates are centrifuged at 12,000–14,000 rpm for 10–15 min at 4°C. Supernatants are collected and filtered through 0.22 µm syringe filters before LC-MS analysis. An internal standard (e.g., demethoxycurcumin or an appropriate synthetic analog) is added prior to extraction to ensure quantification accuracy. The quantification of curcumin and its derivatives is performed using LC–MS. 16
MTT assay
IMR-32 cells are seeded in 96-well plates at 5600 cells per 0.1 mL per well and incubated at 37°C in a humidified atmosphere containing 5% CO2 and 95% air. After overnight attachment, cells are treated with different concentrations of curcumin, curcumin monoglucoside, and curcumin diglucoside for 24 h. Following treatment, cell viability is assessed using the MTT assay according to the manufacturer's instructions. MTT reagent is added to each well, and the plate is incubated to allow formazan crystals to form. The crystals are then solubilized by adding DMSO, and absorbance is measured at 570 nm using a microplate photometer. All treatments are performed in duplicate. Dose-response curves are generated, and IC50 values are calculated from the growth inhibition data. The IC50 concentrations obtained from the analysis are used for all subsequent experiments.
Trypan blue exclusion assay
To determine total cell viability and concentration, IMR-32 cells were harvested by trypsinization with Trypsin-EDTA. The resulting cell suspension was centrifuged at 1500 rpm for 3 min, then resuspended in fresh DMEM supplemented with 10% FBS. An aliquot of the cell suspension was mixed with an equal volume of 0.4% (w/v) Trypan blue solution. After a 2-min incubation at room temperature, 10 µL of the mixture was loaded into a Neubauer hemocytometer. Viable (unstained) and non-viable (blue-stained) cells were counted under a Nikon Eclipse E200 microscope. Cell viability was calculated.
Cell cycle analysis
IMR-32 cells (1 × 105 cells/flask) are seeded in T25 flasks and allowed to adhere for 24 h. After incubation, the DMEM culture medium is replaced with fresh medium containing the IC50 concentrations of curcumin, curcumin monoglucoside, and curcumin diglucoside, and the cultures are incubated for 24 h. Cells are harvested by gentle trypsinization, collected into centrifuge tubes, and washed with PBS. The cell pellets are fixed by slowly adding chilled 70% ethanol while vortexing, followed by overnight storage at −20°C or at least 2 h of fixation. After fixation, the cells are washed to remove residual ethanol and stained with PI staining buffer (5 µg/mL PI) containing RNase A, prepared according to the manufacturer's guidelines. The samples are incubated at room temperature in the dark for 20–30 min to ensure complete RNA digestion and uniform DNA staining. The stained cells are analyzed on a BD FACS Aria II flow cytometer. The proportions of cells in the G0/G1, S, and G2/M phases are quantified using MoD Fit LT software.
Apoptosis analysis
IMR-32 cells are seeded into T25 flasks and allowed to adhere for 24 h. After incubation, the existing DMEM culture medium is removed and replaced with fresh medium containing an IC50 dose of the test compounds: curcumin, curcumin monoglucoside, or curcumin diglucoside. The cells are treated for an additional 24 h. After treatment, the cells are detached by trypsinization and prepared for analysis of apoptotic and necrotic cell death. The analysis involves a double-staining procedure using FITC-conjugated Annexin V and Propidium Iodide (PI). This procedure relies on Annexin V binding to phosphatidylserine (which is externalized on the cell surface during early apoptosis) and PI labelling of cells with compromised membranes, characteristic of late apoptotic/necrotic stages. The staining is performed according to the manufacturer's incubation protocol. Finally, the percentage of apoptotic cells is quantified by flow cytometry, and the acquired data are processed and analyzed using the FACS Diwa Software.
Nuclear changes through confocal changes
IMR-32 cells were seeded onto cover slips, grown overnight, and then treated with IC50 concentrations of curcumin and its derivatives, curcumin monoglucoside and curcumin diglucoside, for 24 h to observe morphological changes. Images were captured at 20× magnification using an inverted Leica confocal microscope. Outer morphology was observed in bright-field images, and the nucleus was stained with DAPI.
Effect on ROS, total antioxidant activity, and 8-Oh-G*
The IMR32 cells are treated with curcumin at its IC50, curcumin monoglucoside, and curcumin diglucoside. ROS levels are measured within the cells using Bioivt kits, and antioxidant activity is assessed using DCFH. 8-OH-G* is measured using Creative Diagnostic kits. Doxorubicin is used as a positive control.
All the above data are statically analyzed for p value.
Results
Intracellular estimation of curcumin and its derivatives
Curcumin increases from ∼50 ng/mg at 0.5 h to a peak of ∼60 ng/mg at 1 h, then steadily declines to ∼12 ng/mg at 24 h. Curcumin monoglucoside rises from ∼80 ng/mg at 0.5 h to a maximum of ∼130 ng/mg at 2 h, then declines to ∼45 ng/mg at 24 h. Curcumin diglucoside shows the highest intracellular accumulation, rising from ∼120 ng/mg at 0.5 h to a peak of ∼210 ng/mg at 2 h, then decreasing to ∼85 ng/mg at 24 h. Overall, intracellular concentration follows the order: curcumin diglucoside > curcumin monoglucoside > curcumin at all time points. The glucoside derivatives exhibit greater cellular uptake and retention than native curcumin (Figure 2).

Bioavailability of curcumin and its derivatives in IMR32 cells.
MTT and trypan blue assays
The comparative analysis shows a clear, dose-dependent decrease in IMR-32 cell viability after curcumin treatment, as evidenced by both the MTT and Trypan blue exclusion assays. Across the tested range of log concentrations (−0.5 to 2.0 μM), viability declines significantly, dropping from approximately 95% at the lowest concentration to nearly 45% at the highest. Both methods show strong correlation, with the MTT assay consistently reporting slightly higher viability than the Trypan blue assay at every data point, suggesting that metabolic decline may lag slightly behind loss of membrane integrity. Overall, the overlapping trends from both assays confirm that curcumin effectively induces cell death in IMR-32 neuroblastoma cells in a concentration-dependent manner (Figure 3(a)). Doxorubicin: The overlaid comparative analysis reveals that Doxorubicin treatment induces a rapid, dose-dependent decline in IMR-32 cell viability, with nearly identical profiles between the MTT and Trypan blue assays. Starting from an initial viability of approximately 60% at the lowest log concentration (0.0 µM), both metrics show a steep reduction to 30% at log 0.5 µM, followed by a steady decrease toward a plateau. From log 1.0 µM onward, the two curves overlap perfectly, indicating that the loss of mitochondrial metabolic activity (MTT) occurs simultaneously with the loss of cell membrane integrity (Trypan blue) in this cell line. At the maximum log concentration of 2.7 µM, cell viability reaches its lowest point at approximately 10%, confirming the high potency of Doxorubicin across both functional and structural markers of cell health (Figure 3(b)). Curcumin monoglucoside: The comparative cytotoxicity analysis reveals a sharp, concentration-dependent decline in IMR-32 cell viability as treatment concentration increases. Both assays follow a nearly identical downward trend, indicating a strong correlation between metabolic impairment and loss of membrane integrity. Notably, the MTT assay consistently reports slightly lower viability percentages than the Trypan blue exclusion assay across all concentrations; for instance, at the maximum concentration of 160 µM, MTT viability drops to approximately 20%, while Trypan blue remains at 25%. This subtle divergence suggests that the treatment may affect mitochondrial dehydrogenase activity more severely or earlier than it compromises the physical stability of the cell membrane, though both metrics confirm the compound's potent cytotoxic effect on the neuroblastoma cell line (Figure 3(c)). Curcumin diglucoside: The comparative plot shows a steep, dose-dependent reduction in IMR-32 cell viability as the log treatment concentration increases from 0 to 2.6 µM. Both the MTT and Trypan blue assays show strong concordance, with 100% viability in the control and a significant drop at higher concentrations. Interestingly, a crossover effect is observed: at lower log concentrations (around 1.0 µM), the MTT assay indicates higher metabolic viability (80%) than the Trypan blue exclusion (75%), whereas at higher concentrations (above 1.8 µM), the MTT values decline more steeply than the Trypan blue counts, ending at approximately 5% and 8%, respectively. This suggests that while membrane integrity is initially more sensitive to the treatment, the eventual collapse of mitochondrial metabolic activity is more profound at higher doses, with both assays confirming potent cytotoxicity (Figure 3(d)).

Estimation of IC50 concentrations of curcumin and its derivatives by MTT assay in IMR32 cell lines.
Cell cycle assay for curcumin and its derivatives
Cell cycle profiling was performed to determine how curcumin and its glucoside derivatives affect cell cycle progression compared with untreated cells and the positive control, doxorubicin. The results show that each compound induced a distinct pattern of phase arrest. Untreated cells displayed a normal distribution across phases, with many cells in the S phase (48.15%), followed by G0/G1 (39.68%) and a smaller proportion in G2/M (12.17%), indicating normal proliferative activity. Curcumin treatment led to a strong accumulation of cells in the S phase (76.62%), indicating that it predominantly induces S-phase arrest, which interferes with DNA synthesis and reduces cell division. The corresponding histograms show an increased S-phase peak compared with the control. Curcumin monoglucoside produced a significant increase in the G0/G1 population (66.44%) with a sharp reduction in S-phase cells (6.54%). This indicates that the monoglucoside leads to G0/G1-phase arrest, preventing cells from entering DNA synthesis. The histogram shows a dominant G1 peak, consistent with this arrest. Curcumin diglucoside markedly increased the G2/M population (39.54%), indicating a prominent G2/M arrest. This suggests that cells treated with the diglucoside derivative are unable to complete mitosis. The histogram supports this with an elevated G2/M peak. Doxorubicin, the positive control, exhibited a strong S-phase arrest, with 69.08% of cells in S phase, consistent with its known mechanism of disrupting DNA replication. The histogram clearly reflects this shift toward S-phase accumulation, as shown in Figure 4(a)-(e) and Table 1.

Effect of curcumin and its derivatives on cell cycle pathways like S, G0, G1, and G2 phases.
Effect of curcumin and its derivatives on cell cycle pathways.

Role of curcumin and its derivatives in inducing apoptosis and necrosis
Annexin V–FITC/PI staining was performed to evaluate the mode of cell death after treatment with curcumin and its glucoside derivatives. The distribution of cells across the four quadrants—live cells, early apoptosis, late apoptosis, and necrosis—was determined for each condition. The untreated sample showed 98.5% live cells, with only 0.6% early apoptosis and 0.2% late apoptosis, confirming normal viability and minimal natural cell death. This served as the baseline reference. Doxorubicin induced marked apoptosis, with 29.2% early apoptotic cells and 1.5% late apoptosis. The live cell population decreased significantly to 69%, indicating strong induction of apoptosis and high cytotoxicity. Curcumin-treated cells showed increased necrosis (8.4%), significantly higher than in any other treatment group. Only 1% total apoptosis (early + late) was observed, and 91.6% of cells remained viable. These data suggest that curcumin at the tested concentration induces more necrotic rather than apoptotic cell death. Curcumin monoglucoside demonstrated very low apoptotic activity, with only 2.6% early apoptosis and 0.1% late apoptosis. A high percentage of cells (95.9%) remained viable. This indicates that mono-glycosylation substantially reduces curcumin's ability to trigger programmed cell death. Curcumin diglucoside induced slightly higher apoptosis than the monoglucoside derivative, with 2.3% early apoptosis and 1.7% late apoptosis, totaling around 4% apoptosis. The live cell population remained high (94.8%), indicating that although diglucoside is more active than monoglucoside, its apoptotic potential is still relatively low, as shown in Figure 5(a)-(d).

Role of curcumin and its derivatives in causing apoptosis and necrosis changes in IMR32 cells.
Morphological changes in the nucleus after treatment with curcumin and its derivatives, curcumin monoglycosides and curcumin diglycosides in IMR-32 cells
The primary finding is that IMR-32 cells treated with curcumin, its derivatives (curcumin monoglycosides and curcumin diglycosides), and the standard drug doxorubicin shows morphological changes after 24 h of treatment (Figure 6). Bright-field images were obtained after treatment with curcumin, its mono- and di-curcumin glycoside derivatives, and doxorubicin. The nucleus was stained with DAPI. Doxorubicin induced disrupted and elongated morphological changes in the nucleus and outer membranes. Curcumin induced moderate changes, as indicated by earlier general morphological studies. The curcumin glycosides (mono- and di-) did not induce any significant changes in IMR-32 cells, as indicated by confocal imaging (Figure 7).

The morphological changes in IMR32 cells upon treatment with Condition A: Untreated, Condition B: Curcumin, Condition C: Curcumin monoglucoside, Condition D: Curcumin diglucoside, and Condition E: Doxorubicin as positive control.

The role of curcumin and its derivatives in causing redox-active oxygen species in IMR32 cells with doxorubicin as a positive control. The curcumin mono- and diglucoside values are statistically significant at p > 0.001 compared to curcumin and doxorubicin.
ROS, antioxidant activity, and 8-OH-G* levels
There is a significant increase (p > 0.001) in ROS induced by curcumin and doxorubicin, whereas curcumin glucosides did not induce any significant change in ROS (Figure 7). The antioxidant activity levels are significantly elevated by Curcumin glucosides (mono- and di-), whereas Curcumin and doxorubicin show no significant changes (Figure 8). The 8-OH-G* levels are significantly elevated (p > 0.001) by curcumin and doxorubicin, whereas curcumin glucosides (mono- and di-) did not elevate the 8-OH-G* levels (Figure 9). These data indicate that curcumin and doxorubicin act as prooxidants, whereas curcumin glucosides act as antioxidants.

The effect of curcumin and its derivatives on antioxidant activity in IMR32 cells with doxorubicin as a positive control. The curcumin mono- and diglucoside values are statistically significant at p > 0.001 compared to curcumin and doxorubicin.

The role of curcumin and its derivatives in modulating 8-hydroxy-guanosine (8-OH-G*) in IMR32 cells with doxorubicin as a positive control. The curcumin mono- and diglucoside values are statistically significant at p > 0.001 compared to curcumin and doxorubicin.
Discussion
Our data indicate three key findings: (1) The intracellular concentration follows the order: curcumin diglucoside > curcumin monoglucoside > curcumin at all time points. The glucoside derivatives exhibit greater cellular uptake and retention than native curcumin; (2) curcumin and the positive control doxorubicin have lower IC50 values than curcumin monoglucoside and diglucoside; (3) the three compounds induce distinct cell-cycle arrests (curcumin: S-phase; curcumin monoglucoside: G0–G1; curcumin diglucoside: G2); and (4) the modes of death differ (curcumin → predominantly necrosis; mono- and diglucosides → early apoptosis that later progresses to necrosis; doxorubicin control → apoptosis only). Our main question is how glycosylation can change potency (IC50). Glycosylation alters physicochemical properties (water solubility, polarity), cellular uptake, subcellular distribution, and metabolic stability of small molecules. Limited studies on the synthesis of glucoside-based curcumin derivatives have reported that specific O-glucoside derivatives exhibit enhanced anticancer activity compared with curcumin, attributed to improved solubility and altered interactions with intracellular targets (e.g., 4-O-β-glucosides showed improved activity in selected cell lines). However, glycosylation effects are not monotonic: position and number of sugar units change membrane permeability, recognition by transporters (e.g., GLUTs, SGLTs), and susceptibility to intracellular glycosidases, producing a prodrug-like behavior. Thus, a diglucoside can be more active than a monoglucoside if (a) it is better taken up (via sugar transporters) or (b) it is more stable in medium and releases the active aglycone intracellularly at concentrations sufficient to inhibit targets. Conversely, the monoglucoside might show a higher IC50 than the diglucoside because the single sugar either reduces passive membrane permeability without providing an effective transporter route, or it is cleared/metabolized differently, so the intracellular free aglycone is lower. These mechanistic possibilities have precedent in curcumin glycosides. The lower IC50 of curcumin and the monoglucoside could reflect either higher intrinsic target potency or simply higher intracellular exposure; distinguishing these requires measuring intracellular compound levels and/or enzymatic diglucosylation rates.17–20
Cell-cycle arrest depends on which molecular node is predominantly perturbed (e.g., DNA replication machinery → S arrest; cyclin D/CDK4–6 axis → G0–G1; DNA damage checkpoint/chromosome segregation → G2/M). Curcumin has been widely reported to cause S-phase accumulation in many cancer cell types, consistent with interference with DNA replication, topoisomerase function, or replication-stress signaling (ATR/Chk1 pathways). Glycosylation can alter molecular interactions: a glycoside that cannot enter the nucleus or that preferentially engages membrane or cytosolic signaling targets may cause G0–G1 arrest (via p21/p27 upregulation, cyclin D downregulation), whereas another derivative that produces DNA damage or microtubule problems might provoke G2 arrest. Thus, curcumin (aglycone), which causes S-phase arrest, is consistent with published observations that curcumin impairs replication and activates S-phase checkpoints. Curcumin-monoglucoside causing G0–G1 arrest suggests this compound either (a) preferentially upregulates CDK inhibitors (p21, p27) or (b) activates upstream signals that prevent G1→S transition (e.g., modulation of growth-factor signaling). Curcumin diglucoside producing G2 arrest suggests induction of DNA damage or microtubule/mitotic spindle stress, or potent activation of the G2/M checkpoint (Chk1/Chk2, ATM/ATR).21–22
Curcumin is reported to induce apoptosis, autophagy, and, at higher concentrations or under certain conditions, necrosis/necrotic-like death; the balance depends strongly on dose, exposure time, redox status, and cell type. Studies report that curcumin elicits early apoptotic signals but can rapidly progress to secondary necrosis (loss of membrane integrity), particularly when caspase activity is insufficient or when ROS are high; others report direct necrosis at high doses/short exposure. That curcumin treatment causes more necrosis could reflect either a higher effective intracellular dose or strong oxidative stress that overwhelms the apoptotic machinery. By contrast, the monoglucoside and diglucoside apparently initiate apoptosis (Annexin V+, caspase activation) and then later show low necrotic features—a classic pattern of secondary necrosis following apoptotic progression in vitro (especially where phagocytic clearance is absent). Doxorubicin is a canonical apoptosis inducer (topoisomerase II poisoning, ROS, DNA damage), and studies documented predominantly apoptotic death with Doxo, which fits our control.21–23 Further, curcumin glucosides both mono- and diglucosides do not alter any changes in nuclear membranes as evidenced by confocal images while curcumin moderately affected while positive control doxorubicin significantly altered the nuclear membrane architect.
A more comprehensive discussion of the pharmacokinetics of curcumin and its derivatives, specifically in relation to their structural features, would enhance the better understanding the pharmacokinetics of novel curcumin-derivatives. Native curcumin exhibits poor aqueous solubility, rapid metabolism, and low systemic bioavailability, primarily due to its β-diketone moiety and phenolic hydroxyl groups, which increase susceptibility to phase II conjugation. Following oral administration, curcumin undergoes extensive first-pass metabolism in the intestine and liver, resulting in glucuronide and sulfate conjugates, as well as reduced metabolites such as dihydrocurcumin and tetrahydrocurcumin. Structural modifications, such as methylation of hydroxyl groups, alteration of the diketone bridge, or incorporation into nanoparticles and liposomes, can improve pharmacokinetic properties by enhancing metabolic stability and membrane permeability. The relationships between chemical structure and ADME (absorption, distribution, metabolism, excretion) are well established and helps to understand the mechanistic rationale for the biological effects of derivatives. Existing literature on curcumin metabolite pharmacokinetics demonstrates how structural modifications influence biological activity. For example, tetrahydrocurcumin is more stable and, in some cases, exhibits greater antioxidant activity than the parent compound, even at lower plasma concentrations. Pharmacokinetic studies indicate that these metabolites can contribute substantially to in vivo efficacy; therefore, the therapeutic potential of curcumin derivatives should not be evaluated solely based on the parent compound. Conjugated forms, which are often regarded as inactive, may function as reservoirs that release free curcumin at target sites. Incorporating data on metabolite formation, bioavailability, half-life, tissue distribution, and clearance, in conjunction with structural considerations, would provide a more comprehensive and mechanistically informed perspective on curcumin derivatives.24–30
Our proposed mechanistic hypothesis for differential neurochemical profiles of curcumin and its glucoside-added derivatives
IMR-32 is a human neuroblastoma cell line widely used to study neuronal differentiation, neurotoxicity, and cancer-related signaling. Curcumin is a polyphenolic compound with reported antioxidant, anti-inflammatory, and anticancer properties.31–33 Glycosylation of curcumin (e.g., monoglucoside, diglucoside) increases water solubility and alters cellular uptake and intracellular distribution. Our data indicated progressively higher IC50 values for curcumin derivatives (curcumin < monoglucoside < diglucoside) than for doxorubicin, suggesting reduced cytotoxic potency but potentially distinct mechanisms of action.
Our central hypothesis, as shown in Figure 10, is that glycosylation of curcumin shifts its action in IMR-32 cells from predominantly pro-oxidant/cytotoxic to redox-stabilizing and cell-cycle-modulatory activities. Specifically, at pharmacologically active concentrations, curcumin induces ROS-mediated mitochondrial apoptosis and G2/M arrest, whereas the monoglucoside and diglucoside act as milder modulators: they reduce baseline ROS, enhance antioxidant defenses (e.g., upregulate Nrf2 targets), and preferentially induce cytostatic effects (G1 arrest) without robust caspase-dependent apoptosis. This graded activity enables glycosylated derivatives to protect non-transformed neuronal cells while still limiting neuroblastoma proliferation. The balance between pro-oxidant and antioxidant outcomes is concentration- and context-dependent. Curcumin may be better suited as a direct cytotoxic agent at higher doses, whereas glycosylated derivatives could be used to reduce toxicity in healthy neurons or to serve as adjuvants that sensitize cancer cells to other chemotherapeutics by modulating redox homeostasis and cell-cycle checkpoints. Our data suggest that the distinct cell-death phenotypes elicited by curcumin (aglycone), curcumin monoglucoside, and curcumin diglucoside arise from fundamental differences in cellular uptake dynamics, intracellular processing, and the kinetics of oxidative stress induction. We hypothesize that curcumin (aglycone), at the concentrations used in our study, is rapidly internalized and partitions into lipid-rich microdomains of the plasma membrane. Owing to its highly conjugated structure and redox-active phenolic groups, aglycone curcumin may generate high levels of ROS within a short time frame, leading to lipid peroxidation of the plasma membrane. This rapid oxidative injury is expected to cause acute loss of membrane integrity, culminating in primary necrosis. The predominance of necrotic morphology in aglycone-treated cells supports a mechanism in which catastrophic membrane damage outpaces the activation of canonical apoptotic signaling pathways.
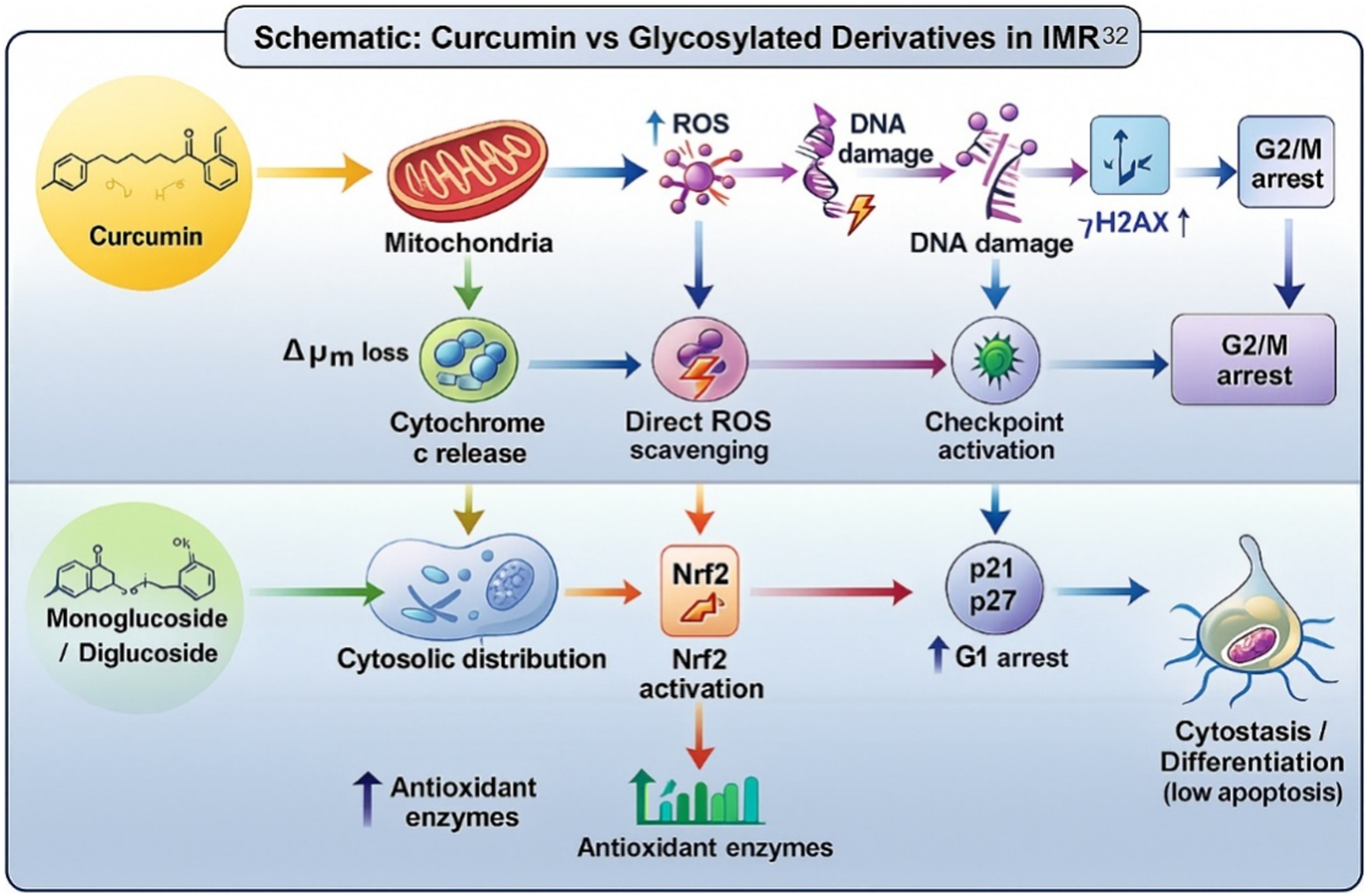
Schematic model comparing proposed intracellular fates and downstream effects of curcumin (left) versus glycosylated curcumin derivatives in IMR-32 cells. Curcumin is predicted to be more prooxidant and proapoptotic, and glycosylated derivatives are more antioxidant and cytostatic.
In contrast, glycosylated curcumin derivatives appear to follow a markedly different trajectory. Their hydrophilic sugar moieties increase aqueous solubility and reduce membrane partitioning, likely slowing cellular uptake and limiting the immediate generation of membrane-associated ROS. These conjugates may require intracellular glycosidase-mediated cleavage to release the active aglycone. We propose that this gradual intracellular liberation results in a slower rise in oxidative stress, allowing caspase-dependent apoptotic cascades to engage before extensive membrane rupture occurs. This model is consistent with the observation that curcumin monoglucoside predominantly arrests cells at G0–G1 and induces features of early apoptosis, whereas curcumin diglucoside arrests cells at G2–M and produces a mixed apoptotic profile. Another possibility is that the glycosides act as prodrugs, with delayed aglycone formation enabling an initial apoptotic phase that later transitions to secondary necrosis once mitochondrial integrity deteriorates or ROS accumulate beyond apoptotic thresholds. Such a biphasic model would reconcile the presence of apoptotic markers early in treatment with the appearance of necrotic morphology at later time points.
Together, these observations support a unified mechanistic framework in which aglycone curcumin induces rapid necrosis through acute oxidative membrane injury, whereas glycosylated derivatives promote controlled apoptotic death through slower cellular processing and delayed aglycone release. Further studies integrating time-resolved ROS measurements, lipid peroxidation assays, glycosidase activity profiling, and mitochondrial function analyses will be essential for validating this model and clarifying the precise biochemical sequence leading to these divergent cell death outcomes.
Footnotes
Acknowledgements
The authors thank the heads of ICMR-NIN, Hyderabad, India, and KLEF, Vijayawada, India, for their facilities and support.
Author contribution(s)
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: LSE is thankful to KLEF for its financial support through a doctoral fellowship. Lakshmi Sowmya Emani, (grant number KLEF-02 ).
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data availability statement
All data generated or analyzed during this study are included in this published article and available from the corresponding author's laboratory.