
Abstract
Helicobacter pylori remains a significant global health concern, with rising antibiotic resistance posing challenges for conventional treatments. Bacteriophages, viruses that specifically target and lyse bacterial cells, present a promising alternative therapeutic approach. This review explores the advancements in phage research related to H. pylori, including the isolation, genomic and proteomic characterization, and therapeutic potential of lytic and lysogenic phages. Novel isolation techniques have identified diverse phages from clinical and environmental sources, such as sewage and wastewater, revealing unique genetic and structural adaptations that enhance their effectiveness in targeting H. pylori. Genomic analysis has highlighted the role of prophages in H. pylori evolution, while proteomic studies have uncovered structural features that enable phages to survive the acidic gastric environment. High-throughput technologies, such as next-generation sequencing, clustered regularly interspaced short palindromic repeats and CRISPR-associated proteins (CRISPR-Cas) systems, and microfluidic platforms, have revolutionized phage discovery and characterization. Furthermore, the potential for phage-antibiotic synergy offers new avenues for combating antibiotic resistance. Despite these advancements, challenges such as H. pylori’s genetic diversity, its fastidious growth requirements, and the development of robust delivery mechanisms for gastric application persist. This review highlights the need for further research to optimize phage-based therapies as a viable alternative or adjunct to current treatments for H. pylori infections.
Overview of Helicobacter pylori
Helicobacter pylori is a slow-growing and fastidious bacterium that infects approximately 43% of the world’s population. 1 Infection with this bacterium has been associated with the development of gastroduodenal diseases that include peptic ulcers, gastritis, and gastric cancer. World Health Organization has declared H. pylori as Type I carcinogen that causes gastric cancer, and it is currently the only bacteria listed in that category. 2 Gastric cancer causes approximately 700 thousand deaths worldwide annually, and it is still one of the most commonly diagnosed cancers in the world, with approximately one million cases diagnosed every year. 3 Prognosis and diagnosis of gastric cancer are poor, with most cases diagnosed at late stage, particularly in developing countries. 4 Thus, eradication of H. pylori is a strategy to prevent development of gastric cancer. Eradication of H. pylori can involve several treatment regimens. In first-line therapy, administration of proton pump inhibitor (PPI) coupled with antibiotics, namely amoxicillin and clarithromycin, is prescribed to patients. However, if first-line therapy failed, clinicians can prescribe bismuth quadruple therapy that consists of bismuth, PPI, metronidazole, and tetracycline as second-line therapy or triple therapy that consists of levofloxacin, amoxicillin, and PPI as second-line therapy. 5 However, the emergence of H. pylori strains that are resistant to multiple antibiotics has complicated treatment strategy to eradicate this pathogen and prevent gastric cancer. 6
Helicobacter pylori harbors many virulence factors that are involved in pathogenesis of gastric diseases. These factors include cytotoxin-associated A (CagA) protein essential in disturbing normal cellular signaling, vacuolating cytotoxin A (VacA) important in formation of vacuole in cells, and urease pertinent in neutralization of stomach acid. 7 Interestingly, more than 80% of H. pylori strains isolated from Asian populations harbor CagA, while only 60% of H. pylori strains from Western populations harbor this protein, which may partially explain why the Asian population has a high prevalence of gastric cancer compared to the Western population.8,9 Apart from those factors, some strains also harbor a cluster of virulence genes located at the cag pathogenicity island that are important in secretion of inflammatory cytokines and formation of secretion system to transfer CagA oncogene. 10 Helicobacter pylori genetic is highly diverse and is strongly associated with its host. Global phylogenetic analysis of H. pylori isolated from different world population reveals that H. pylori can be divided into four primary clusters, including (1) Southwest Europe, (2) Central, East, and South Asia, (3) Southern and Western African cluster, and (4) Northern and Southern Europe, and Middle East cluster. 11 Highly diverse genetic of H. pylori shows that its interaction with host is variable and may be a key factor in its pathogenesis.
Bacteriophages in H. pylori
Bacteriophages, or phages, are viruses that specifically infect and kill bacteria. They are highly specific to their bacterial hosts and play a crucial role in controlling bacterial populations in various environments, from soil and water to the human microbiome. Phages operate by attaching to a bacterial cell, injecting their genetic material, and using the bacterial machinery to replicate. This eventually leads to the lysis of the bacterial cell, releasing new phage particles. 12 Due to their specificity and ability to kill bacteria, phages are considered a promising alternative to antibiotics in treating bacterial infections, especially those resistant to conventional treatments.
The first bacteriophage was discovered in 1915 by British bacteriologist Frederick W. Twort, who observed a “glassy transformation” in bacterial colonies that was caused by an unknown agent. 13 Two years later, in 1917, French-Canadian microbiologist Félix d’Hérelle independently discovered bacteriophages while studying patients with dysentery. 14 He named the agent “bacteriophage,” meaning “bacteria eater,” and proposed the concept of using bacteriophages therapeutically to treat bacterial infections. These discoveries marked the beginning of the scientific study of bacteriophages.
The first study investigating bacteriophages for H. pylori dates back to the early 1990s. In this study, researchers identified a lysogenic strain of H. pylori that spontaneously produced phage particles. 15 The study demonstrated the potential presence of bacteriophages that could infect and lyse H. pylori. This finding was significant as it suggested that phages might play a role in the genetic diversity and evolution of H. pylori and opened the possibility for phage therapy as a treatment option for H. pylori infections. These discoveries marked the beginning of the scientific study of bacteriophages. This review aims to observe the advancements and evolution of bacteriophages associated with H. pylori and explore recent developments in phage therapy. Although some studies have detected the presence of phage sequences in H. pylori genome using next-generation sequencing (NGS) platform, 16 only a few studies have successfully isolated lytic and temperate phages either from environmental sources (lytic) or the bacteria itself (lysogenic). The bodies of research that have successfully isolated lytic or lysogenic phages related to H. pylori are presented in Table 1. All studies have examined the isolated lytic and lysogenic phages of H. pylori in vitro, but not in vivo models.
Overview Progress of Researches on the Isolation of Phages Related to Helicobacter pylori

Isolation and Identification of Novel Phages Against H. Pylori
Discoveries of novel phages specifically targeting H. pylori have focused on identifying and characterizing phages that could be used for therapeutic purposes, especially considering the rising antibiotic resistance of H. pylori. Many studies analyzing the genomes of Helicobacter species and confirmed the presence of prophages within these bacterial genomes and examined their roles and functions.19,25,26
Studies have isolated new temperate and lytic phages that show potential in targeting H. pylori.23,24 These novel lytic phages have been identified that can infect multiple strains of H. pylori, and genome sequencing has revealed unique genetic features that may help overcome bacterial resistance mechanisms. They revealed that these phages have specific genes or genetic traits that are not commonly found in other known phages. These unique features could be linked to their ability to infect H. pylori or to evade bacterial defenses.19,21 Nevertheless, isolation of phage from H. pylori is challenging, considering that it is fastidious and slow-growing bacteria. Culture of H. pylori from stomach biopsies is difficult as it requires extensive training in the laboratory. Besides that, H. pylori usually requires 5–7 days incubation before colony growth can be observed. A previous study adopted enrichment procedure (coculture of bacteria with filtered environmental sample) using special media for H. pylori culture, including Columbia broth prior to spot test to increase the chance of phage isolation. In addition, all procedures involving enrichment and phage isolation must be conducted in microaerophilic condition. 23 Similar to lytic phage isolation, induction of lysogenic phage from H. pylori also has challenges that include the culture condition of H. pylori and diverse H. pylori genomes that differ according to the population from which it is isolated. A recent study reveals that only less than 30% of H. pylori genomes assessed harbor prophage, and the presence of prophage in H. pylori is associated with the population where H. pylori was isolated. 27
Bacteriophages can be isolated from various samples, including environmental sources such as sewage, wastewater, soil, and water bodies such as rivers and lakes, which are often rich in diverse microbial communities (Fig. 1). Additionally, clinical samples such as biopsies from patients and positive H. pylori antigen stool samples have also been utilized to isolate phages. Most studies have identified phages against H. pylori through genome sequence analysis, revealing the presence of prophages that can be induced to enter a lytic cycle and target the bacterial cells. 23 The strains containing these prophages were isolated from gastric biopsies of patients suffering from gastric diseases.19,21,22,24,26 The study by Lehours et al. 19 sequenced the genome of H. pylori strain B45 using the 454 Titanium platform, revealing a 24.6-kb prophage homologous to H. acinonychis prophage II. Prophage induction was tested using UV irradiation and mitomycin C, with transmission electron microscopy (TEM) confirming phage-like particles resembling Siphoviridae morphology, featuring heads (∼62.5 nm) and tails (∼92.4 nm). PCR screening for the integrase gene in 341 H. pylori strains showed a prevalence of 21.4%, with distinct phylogenetic clustering by geography. The prophage was integrated near restriction modification (R-M) system genes, suggesting its role in H. pylori genome evolution and adaptation. The bacteriophage HPy1R was isolated from H. pylori strain 11057A using UV radiation (24). TEM revealed its Podoviridae morphology, with a short, noncontractile tail and an icosahedral head. Genome sequencing identified a 31,162 bp genome with 37.1% GC content, encoding 36 genes related to DNA replication, morphogenesis, lysis, and packaging. Stability tests showed HPy1R remained viable at pH 3–11 and 37°C, with minor loss of phage concentration under simulated gastric digestion. It formed visible haloes in 78.9% of strains tested, formed plaques in 6.6%, and suppressed bacterial growth for 24 h. Proteomic analysis identified 17 structural proteins, highlighting their therapeutic potential.

Screening of lytic bacteriophages against Helicobacter pylori that starts with sampling from potential sources of the bacteriophage. The samples collected are then tested against H. pylori using multiple tests that include enrichment assay, spot test, and plaque assay. Last, bacteriophage with lytic characteristic is isolated and stored for further experiments that include in vivo and toxicity test. Lysogenic phage can be induced from H. pylori strains with prophage sequences and tested for its anti-H. pylori activity.
The lytic bacteriophages for H. pylori have been successfully isolated from hospital’s sewage and wastewater.23,28 In the study by Khosravi et al. 23 phage isolation was performed using hospital sewage samples screened via spot tests to identify lytic phages against H. pylori. Characterization revealed phage-specific proteins using SDS-PAGE showed the presence of proteins with size of 58 kDa and 64 kDa. These proteins were absent in the bacterial host, and further test using RAPD PCR confirmed that those proteins were distinct from H. pylori. Electron microscopy further validated the phage’s morphology, confirming its successful isolation and unique structural properties. This highlights sewage as a valuable source for isolating lytic phages targeting antibiotic-resistant bacteria. Abdel-Haliem et al. 28 screened wastewater samples for H. pylori-specific phages using the double-layer method, leading to the isolation of two phages, ΦHPE1 and ΦHPE2. Phages were purified using liquid enrichment and polyethylene glycol-dextran sulfate separation. Electron microscopy identified ΦHPE1 (Podoviridae family) with a 62 nm head and short tail (∼12 × 6 nm) and ΦHPE2 (Siphoviridae family) with a 92.5 nm head and long tail (∼180 × 15 nm). Both phages were stable at pH 5–9 and temperatures up to 65°C. Host range tests revealed strains Zag1 and Zag3, which were clinical strains isolated from Egyptian patients, were susceptible to both phages, demonstrating strong lytic potential and suitability for therapeutic applications.
Genomic and Proteomic Characterization of H. pylori Phages
Studies on isolated lytic and lysogenic phages of H. pylori are scarce. Major factor that contributes to the lack of studies on H. pylori phage includes the nature of this bacterium itself that is fastidious and difficult to grow in laboratory. 29 Despite a lack of studies on H. pylori phage, lytic, and lysogenic phages (Fig. 2) of H. pylori have been successfully isolated in previous studies.23,28,30,31
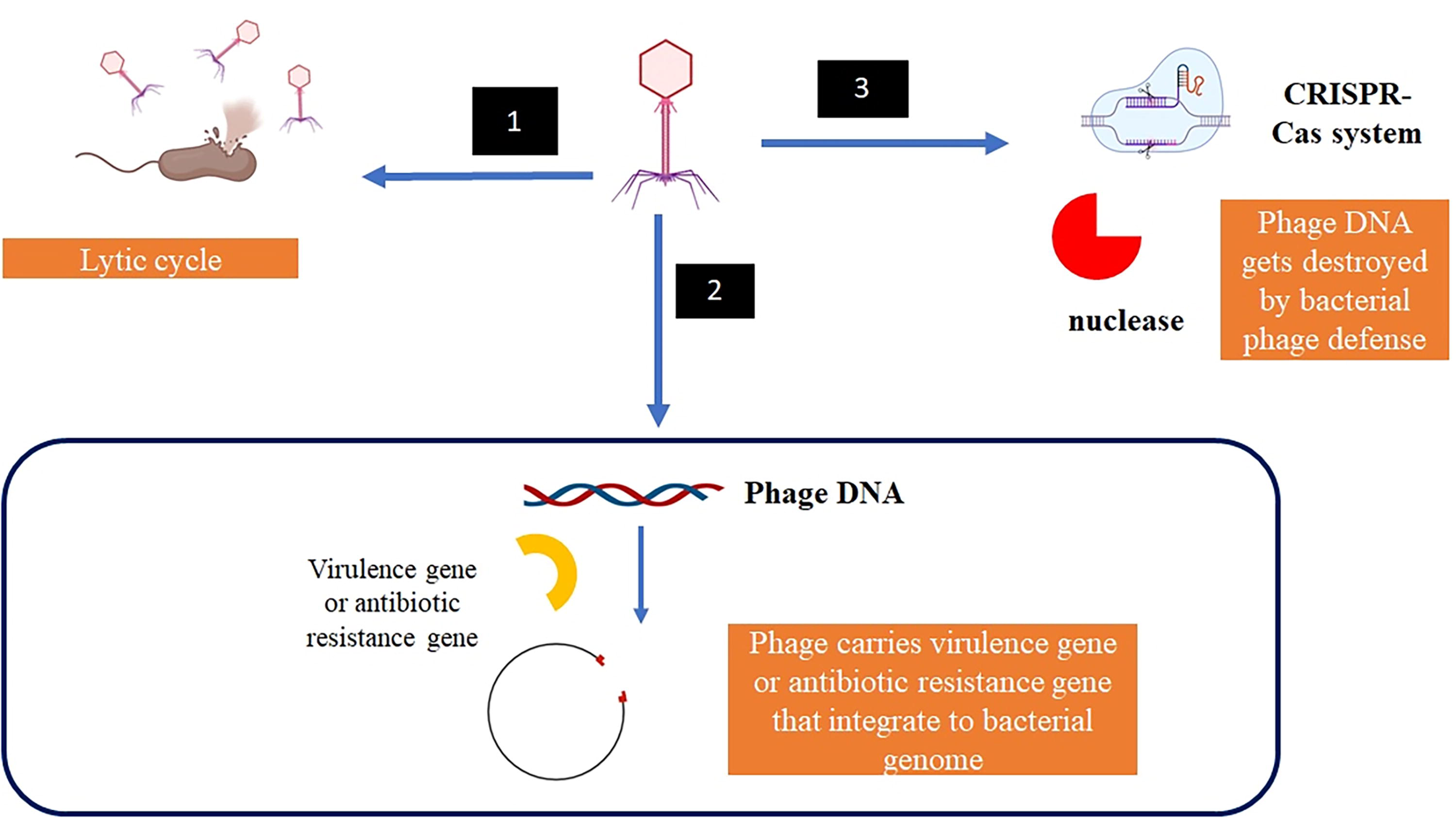
Life cycle of bacteriophage. (1) Bacteriophage infects bacteria and undergoes lytic cycle that causes bacterial cell to burst, (2) Bacteriophage infects bacteria and undergoes lysogenic cycle, in which phage genome is integrated into bacterial genome. This integration may cause bacteria to receive new gene that may cause harm to humans that include virulence factor and antibiotic resistance gene, and in (3) Phage defense system (i.e., restriction modification enzyme) in bacteria destroys phage genome before it can undergo lytic or lysogenic cycle.
Besides lytic phage, lysogenic phage of H. pylori has also been identified, and some studies have isolated lysogenic phage of H. pylori.24,31 While lytic phage is more attractive as an alternative therapy against H. pylori than lysogenic phage, lysogenic phage also can be harnessed and studied for future therapy. Analysis of H. pylori genomes using a large sample size reveals that 29.5% of H. pylori genome harbor prophage, of which almost 33% have complete prophage genome. 19 Similar study also observed that the sizes of H. pylori genomes harboring prophage genes are also bigger than that of genomes without prophage genes. Interestingly, prophage genes in H. pylori genome depends on geographical location where the bacteria were isolated, in which most H. pylori strains isolated from Russian population had prophage gene in their genomes while no prophage gene was detected in H. pylori genomes isolated from Japanese population. 27 Although no prophage gene was detected in H. pylori strains isolated from Japanese population, two lysogenic phages (KHP30 and KHP40) have been successfully induced and isolated from Japanese patient, indicating that they present as episome in H. pylori.30,31 This result suggests that those phages exist at extrachromosomal regions of H. pylori. KHP30 and KHP40 possess dsDNA and have genome sizes of 26.2 kb and 26.4 kb, respectively. Both phages have GC content of 35.8% with open reading frames of 30 and 32, respectively. They also have a gene that encodes integrase, although they exist at extrachromosomal region of bacterial genome. 21 Similarly, lysogenic phage isolated from Taiwanese patient also has genome size ranging from 26 to 27 kb, suggesting that lysogenic phage of H. pylori shares similar genomic characteristic depending on geographical population from which it is isolated. 22 Another lysogenic phage, namely HyPy1, was isolated from Portuguese patient. The phage has genome size of 31.1 kb with GC content of 37.1%. 24 Overall, genomes of H. pylori phages are small compared to phages isolated from other bacteria that include Escherichia coli and Pseudomonas aeruginosa, with phage genome sizes that range from 40 to 90 kb. Furthermore, GC content of phages isolated from E. coli and P. aeruginosa is more than 40%.32,33 Notably, lysogenic phage isolated from both Japanese and Portuguese patients demonstrated stability at low pH 3,24,30 indicating that they harbor capsid proteins that can withstand human’s acidic stomach. Recent structural analysis of KHP30 H. pylori phage uncovers stable structural capsomeres of viral structure that consists of 12 pentons and 80 hexons at T = 9 symmetry. Not only stable capsomeres but the phage structure also possesses protruding loop from major capsid that holds the phage protein subunits together 31 that facilitates the phage to survive in low acidic environment such as human’s stomach.
Phage–Host Interaction Dynamics
Phage–host interaction depends on either phage infection to bacteria results in lytic or lysogenic cycle. Lytic cycle controls population of bacteria as infection with this type of phage leads to bacterial cell rupture and cell lysis, and finally bacterial death. Meanwhile, lysogenic cycle results in integration of phage genome into bacterial genome. 34 Once phage genome is integrated into bacterial genome, this process results in control of bacterial evolution through integration of new gene that may be virulent to humans via horizontal gene transfer. This gene can be virulent gene that evades human’s immune system or antibiotic resistance gene that confers resistance of bacteria to essential antibiotic. 35 Apart from that, integrated phage genome can also disrupt the function of bacterial gene. Assessment of phage genomes in pig reveals the presence of virulence gene essential in causing diarrhea that can be transferred to bacterial genome coexisting in pig’s gut. 36 Antibiotic resistance genes including genes that confer resistance to beta-lactam antibiotics (bla genes), methicillin (mecA), and colistin (mcr-1) have been found in phages isolated from hospital wastewater and water from treatment plants, suggesting phages as one of the main sources for horizontal gene transfer of highly important resistance genes to clinically important bacteria. 37
In order to defend itself against phage infection, bacteria have already evolved mechanisms that can detect the presence of foreign DNA and destroy it. These mechanisms include restriction modification systems, namely restriction DNA enzyme and the presence CRISPR-Cas system that cleaves foreign DNA. 38 H. pylori harbor various restriction modification systems that help its persistence against phage infection to ensure its survival in environment with high density of lytic and lysogenic phages, such as contaminated water and sewage system. 39 The presence of CRISP-Cas system in bacterial genome has been associated with antibiotic resistance in certain bacteria including Klebsiella pneumoniae and Acinetobacter baumannii.40,41 However, recent study conducted for H. pylori strains isolated from patients in Shanghai did not find association between the presence of CRISPR-Cas system with the occurrence of antibiotic resistance in H. pylori. 42 Bacteria also has mechanism, namely phase variation, to change its antigenic surface molecule to deter attachment of phage to its surface. 43 H. pylori harbors several cell surface molecules, namely outer inflammatory protein A, blood group antigen binding A, and sialic acid binding adherence A, that can be switched “on/off” during infection44,45 and how the mechanism of phase variation in these molecules contribute to bacterial immune response against phage infection is unknown.
Advancements in Phage Identification and Isolation
Advances in high-throughput screening techniques for phage identification have significantly enhanced the ability to discover and characterize bacteriophages more efficiently. These techniques employ automated and parallelized methods to rapidly screen large libraries of environmental samples, bacteria, or phages to identify specific bacteriophages that can effectively infect and lyse target bacterial strains.46,47 The following assays or techniques have been described for high-throughput screening for identification of bacteriophages.
High-throughput automated plaque assays
Traditional plaque assays have been adapted into automated high-throughput formats, where robotic systems can handle multiple plates simultaneously, reducing manual labor and increasing the speed of phage identification. Automated systems can detect and measure plaque formation (clear zones where phages have lysed bacterial cells) in real time, allowing rapid screening of large numbers of samples. Automated bacteriophage plaque assays have evolved to enhance the efficiency and precision of phage quantification and characterization. Notable advancements include plaque size tool (PST), micro-plaque assays, and high-throughput bacteriophage testing with automated pipetting. PST is developed to automatically detect and measure bacteriophage plaques. PST processes images of petri dishes to assess plaque morphology, including area and diameter. It supports various plaque types, such as clear, turbid, and bull’s-eye morphologies, and is compatible with multiple image formats. PST is available as an open-source tool on GitHub. 48 Micro-plaque assays are high-throughput method that utilizes a robotic pinning platform to miniaturize traditional plaque assays. By creating microcolonies from minimal inoculant volumes, it significantly increases throughput without compromising sensitivity, facilitating rapid isolation and characterization of bacteriophages. 46 High-throughput bacteriophage testing with automated pipetting employing a liquid-handling robot, this approach automates phage susceptibility testing by performing phage drop-off and potency determination. It enhances reproducibility and reduces variability compared to manual methods, streamlining the evaluation of phage efficacy against multiple bacterial strains. 49 Currently, there is no study published that employed the application of automated plaque assay for H. pylori phages. Challenge to adopt this method in discovery of lytic phage against H. pylori includes the nature of this pathogen that is slow growing. However, optimizing this method in H. pylori study is necessary to reduce workload associated with sample processing and plaque counting of many samples collected from environment that may be the source of lytic H. pylori phage.
Phage display libraries
Phage display libraries are powerful and versatile tools in molecular biology and biotechnology, enabling the display of diverse peptides or proteins on the surface of bacteriophages. First introduced by George P. Smith in 1985, this technology establishes a direct connection between a protein’s phenotype and its encoding genotype, allowing for efficient screening and selection of molecules with desirable properties. By engineering phage genomes to include DNA sequences encoding specific peptides or protein variants, these libraries can display billions of variants, making them invaluable for applications such as antibody engineering, vaccine development, and studying protein–protein interactions. A landmark achievement of this technology is the development of therapeutic antibodies, such as adalimumab, the first fully human antibody approved for clinical use.50,51
Recent advances in phage display technology have further enhanced its applications, particularly in therapeutic antibody discovery and targeted drug delivery. The incorporation of noncanonical amino acids into phage-displayed peptide libraries has increased molecular diversity, enabling the selection of peptides with enhanced binding properties. 52 Furthermore, phage display has played a key role in the development of monoclonal antibodies for cancer treatment, with several phage-derived antibodies entering clinical trials and achieving regulatory approval. 53 These advancements highlight the critical role of phage display libraries in revolutionizing drug discovery, therapeutic development, and molecular biology research.
Phage display library have been used in research and discovery of new therapeutic agents against H. pylori. Previous studies adopted this method to target specific protein on H. pylori surface. Ortiz et al. 54 used phage display to screen antimicrobial peptides that can target VacA protein while Xiong et al. 55 targeted acid-sensing domain that facilitates H. pylori in acidic environment. Interestingly, this study found that antimicrobial peptide from phage display library that antagonize H. pylori acid-sensing domain inhibited the growth of H. pylori in vitro, suggesting the potential use of this peptide against H. pylori. Apart from peptide, chicken antibody against HpaA protein of H. pylori has been designed and screened using phage display library. 56
Next-generation sequencing
NGS techniques provide a comprehensive approach for analyzing the genomic composition of phage populations from environmental and clinical samples. By sequencing entire metagenomes, NGS enables the detection of bacteriophage DNA, identification of phage species, and assessment of their activity against specific bacterial hosts. This method is particularly advantageous for uncovering novel phages that are not detectable through traditional culture-based approaches. NGS has significantly advanced phage research by offering rapid and precise genomic sequencing, supporting the discovery of previously unknown phages and the investigation of their genetic diversity.
NGS has been used to sequence Bacillus cereus phage isolates, providing valuable insights into their genomic structures and evolutionary patterns. Moreover, NGS has facilitated the prediction of genome terminus sequences in Bacillus cereus-group bacteriophages, deepening our understanding of their genome organization and replication strategies. 57 Furthermore, NGS has been applied in phage display research to characterize the composition, quality, and diversity of unselected phage display peptide libraries. A study comparing two NGS platforms with different sequencing depths demonstrated the pivotal role of sequencing depth in accurately assessing phage display libraries. 58
In the context of H. pylori, a bacterium associated with gastrointestinal diseases, NGS has advanced the study of bacteriophages by enabling the identification and characterization of phages that infect this pathogen. This includes sequencing the genome of novel H. pylori bacteriophages, revealing their structural and functional genes, which can guide the development of phage-based therapies against antibiotic-resistant strains. 24 Furthermore, genomes of H. pylori have also been screened for the presence of prophage sequences that can then be induced for isolation of lysogenic phage that can be studied for potential therapy against H. pylori. Using shotgun metagenomic application, environmental sources can be screened for the presence of lytic phage against H. pylori that can then be isolated using standard protocol. Additionally, NGS has been employed to detect mutations in H. pylori strains associated with antibiotic resistance, supporting the design of effective phage therapy strategies. 59 Phage genome can also be screened for the presence of antibiotic resistance and virulence genes to mitigate the management of antibiotic resistance in bacteria. Together, these advancements highlight the pivotal role of NGS in revolutionizing phage research, ranging from elucidating phage–host interactions to driving the development of novel therapeutic applications.
CRISPR-Cas systems
High-throughput CRISPR-based techniques have been developed to identify and isolate specific bacteriophages within mixed populations by targeting their genomes. These methods utilize the precision of CRISPR-Cas systems to selectively cleave phage DNA, facilitating the enrichment of phages of interest. This approach streamlines the identification of new candidates for phage therapy, particularly against antibiotic-resistant bacteria. For instance, Locus Biosciences has engineered bacteriophages with CRISPR-Cas3 systems to target and degrade bacterial genomes, enhancing the specificity and efficacy of phage therapy. Their CRISPR-enhanced bacteriophage cocktail, LBP-EC01, has shown promise in treating urinary tract infections caused by multidrug-resistant Escherichia coli. In a Phase II clinical trial, this therapy demonstrated a significant reduction in bacterial load, highlighting the potential of CRISPR-based phage engineering in developing targeted antibacterial treatments. 60 Additionally, high-throughput platforms integrating CRISPR technologies have been employed to rapidly identify phage–host interactions, facilitating the development of personalized phage therapies. Such platforms enable the screening of vast phage libraries against specific bacterial pathogens, accelerating the discovery of effective phage candidates for therapeutic applications. 61
These advancements underscore the potential of CRISPR-based methodologies in revolutionizing phage therapy by enabling the precise targeting and isolation of bacteriophages, thereby enhancing the development of novel treatments for bacterial infections. Although this technology has been tested in other bacteria such as E. coli, currently, there is no published study on the application of this method for H. pylori. Future study to explore this technology in discovery of new therapy against H. pylori is necessary.
Microfluidic platforms
Microfluidic technologies have been adapted for bacteriophage identification by enabling the manipulation and screening of minute sample volumes with high precision and speed. These platforms create miniaturized environments where individual bacteria and phages can interact, facilitating rapid and efficient screening of phages with desired characteristics. A flexible and efficient microfluidics platform has been developed to study bacteria–phage interactions. This setup allows for high-throughput characterization of bacteriophage activity without the need for external pumps or tubing, enabling direct investigation of bacterial behavior in response to phage exposure. 62 Additionally, droplet-based microfluidic systems have been employed to investigate bacterial lysis by phages. These devices generate anchored droplets containing bacteria and phages, allowing precise observation of interactions and lysis events. Such systems provide a novel approach to quantify phage efficacy at the single-cell level. 63 Furthermore, microfluidic platforms have been utilized for the uniform amplification of phage display libraries. By encapsulating individual phage clones in microdroplets, these systems eliminate competition between clones with different growth rates, ensuring unbiased amplification and selection of phages displaying peptides or proteins of interest. 47
These advancements in microfluidic technology enhance the efficiency and accuracy of phage identification and characterization, contributing significantly to the development of phage-based therapies and diagnostics.
Mass spectrometry
Advancements in mass spectrometry, particularly matrix-assisted laser desorption/ionization time-of-flight (MALDI-TOF) and liquid chromatography–tandem mass spectrometry (LC-MS/MS), have significantly enhanced the rapid identification and characterization of bacteriophage proteins. These techniques facilitate comprehensive proteomic profiling, enabling researchers to elucidate phage protein compositions, posttranslational modifications, and functional roles, thereby advancing the development of phage-based therapeutics.
MALDI-TOF mass spectrometry is widely utilized in proteomics for the structural identification and characterization of proteins and peptides, including their various proteoforms resulting from posttranslational modifications or protein–protein interactions. This technique allows for the rapid analysis of complex protein mixtures, making it invaluable in understanding phage proteomes. 64 LC-MS/MS serves as a powerful analytical tool for separating, sequencing, and quantifying complex protein mixtures. In the context of phage research, LC-MS/MS enables detailed analysis of phage proteins, providing insights into their structural and functional properties. 65 These mass spectrometry-based proteomic analyses are instrumental in identifying unique phage proteins that can be targeted or harnessed for therapeutic purposes, thereby contributing to the development of novel phage-based treatments.
Advances in high-throughput screening techniques, including automated plaque assays, phage display libraries, NGS, CRISPR-Cas systems, microfluidic platforms, and mass spectrometry, have revolutionized the identification, characterization, and therapeutic application of bacteriophages. These technologies allow for rapid and precise screening, enrichment, and analysis of phage populations, overcoming the limitations of traditional methods. By enabling the discovery of novel phages, understanding of phage–host interactions, and identification of unique phage proteins, these advancements provide powerful tools for developing targeted and efficient phage therapies. Collectively, they have paved the way for innovative solutions to combat antibiotic-resistant bacteria, streamline drug discovery, and enhance our understanding of bacteriophage biology, making phage-based applications increasingly viable in modern medicine and biotechnology.
Integration of Phage Therapy with Existing Treatments
Combining bacteriophages with antibiotics has demonstrated synergistic effects, enhancing the efficacy of treatments against multidrug-resistant bacterial infections. This synergy, known as phage-antibiotic synergy (PAS), occurs when sublethal concentrations of certain antibiotics enhance phage production, leading to more effective bacterial eradication. 66
Studies have shown that combining phages with antibiotics can effectively disrupt biofilms, which are often resistant to conventional treatments. For example, a study on Acinetobacter baumannii biofilms revealed that a combination of phage cocktails and antibiotics significantly improved biofilm eradication compared to either treatment alone. 67 A case study reported that adding phage therapy to systemic antibiotics for treating recurrent Enterococcus faecium infections, which were unresponsive to antibiotics alone, resulted in fewer hospitalizations and enhanced quality of life. 68 Additionally, research on carbapenem-resistant Klebsiella pneumoniae (CRKp) has shown that phage-antibiotic combinations can optimize therapeutic efficiency. By analyzing the phenotypic and genotypic characteristics of CRKp-specific phages, researchers developed a systematic model for phage cocktail combinations that enhance antibacterial effects when used alongside antibiotics. 69
Combining phage therapy with antibiotics offers a promising strategy to combat H. pylori infections. Combining phages with antibiotics may enhance treatment efficacy through synergistic effects. This approach could potentially reduce the required antibiotic dosage, thereby minimizing side effects and slowing the development of resistance. However, specific studies on PAS against H. pylori are currently lacking, necessitating further research. The unique environment of the human stomach, along with the bacterium’s genetic diversity and the scarcity of identified lytic H. pylori phages, poses significant challenges to this therapeutic approach. While the concept of combining phage therapy with antibiotics for H. pylori infection is promising, more extensive clinical research is necessary to establish its viability and effectiveness as a treatment modality. Future research should focus on identifying effective phage strains, optimizing treatment protocols, and assessing safety and efficacy through clinical trials.
Current Challenges and Future Directions in Phage Development
One of the main challenges in research and discovery of phage as an alternative treatment against H. pylori is the nature of this bacterium itself that is slow growing and fastidious to grow in the laboratory. 70 Furthermore, isolation of H. pylori requires collaborative effort among gastroenterologist, pathologist, and microbiologist during sample collection of gastric biopsies for bacteria culture. This barrier presents a challenge to low-resource countries with limited access to laboratory equipped with equipment for successful culture of H. pylori. Another barrier that needs to overcome is identification of source for isolation of lytic phage of H. pylori. Theoretically, lytic phages can be isolated from where the bacteria are usually found. For H. pylori, it is found in stomach and wastewater. While procedure for isolation of lytic phage from wastewater or environment sources has been established, the presence of limited H. pylori populations in these environments has hampered discovery of lytic phage. Collection of stomach biopsies for phage isolation needs to have optimized procedure and should be studied further. In addition, the discovery of lytic phage of H. pylori can also be extended to other natural sources that include diverse marine sources. H. pylori genetic is also diverse and lytic phage discovered against one strain of H. pylori may not work against another strain of H. pylori. 11 Thus, concerted effort should be made to characterize genome and receptor for phage binding on bacterial cell surface to tailor the design of phage cocktail that can be used for all strains of H. pylori. Phage discovered as new alternative therapy against H. pylori must also possess capsomeres that can protect them against a harsh acidic environment. Another way to overcome delivery of phage to acidic stomach is the delivery of phage using hydrogel that can protect phage against degradation. 71
Conclusion
Phage has huge potential to be harnessed as a new therapy to eradicate antibiotic-resistant H. pylori and to prevent development of gastric cancer. While lytic phage is an attractive choice for new therapy, lysogenic phage can also be explored for new therapy as lysogenic phage may be induced to become lytic using antibiotic. Nevertheless, isolation of lytic phage against H. pylori is challenging because of the nature of this bacterium that is slow growing and fastidious. Furthermore, delivery of phage to stomach can be difficult because of acidic environment in that organ that can destroy phage without proper drug delivery technique. New technology for research and discovery in therapeutic phage can be explored and adopted to mediate these challenges.
Authors’ Contributions
Conceptualization: A.H., A.S., and B.S.L. Funding acquisition: A.H., A.S., H.N., and B.S.L. Resources: M.A.A., H.Y., and S.M.P. Investigation: M.A.A., H.Y., S.M.P., and H.N. Writing (original draft): A.H. and A.S. Writing (review and editing): A.H., A.S., H.M., and B.S.L.
Footnotes
Acknowledgments
The authors would like to thank Universiti Kebangsaan Malaysia for supporting the student’s fee through the Research Fellowship Scheme (Skim Zamalah Penyelidikan) for M.A.A.
Author Disclosure Statement
All authors declare that there is no conflict of interest.
Funding Information
This work was supported by Universiti Kebangsaan Malaysia through the research university grant scheme (grant no. GUP-2023-070).