
Abstract
Background:
The development of phage libraries is vital for phage-based applications and to further the understanding of phage–host interactions. However, phage isolation from environmental samples on a large scale is resource-intensive.
Methods:
To optimize the detection of phages from environmental water samples with low phage concentrations, we assessed and optimized a small-volume enrichment method using the important fish pathogen Flavobacterium psychrophilum as a target host.
Results:
The 96-well plate enrichment assay demonstrated phage activity in samples with initial phage concentrations of a single plaque-forming unit (PFU) per well, and the overall detection limit of the method was 0.011 PFU mL−1 in the water sample. For phage concentrations >10 PFU, the phage concentration in the sample could be predicted from optical density inhibition during incubation. When applied to 45 water samples collected from a recirculating aquaculture system, phages were detected in 36 of the samples.
Conclusion:
While this method provides resource-efficient means to isolate phages from aquatic environments, phage–host system-dependent variations are shown, and optimization for other phage–host systems is recommended.
Introduction
The impact of Flavobacterium psychrophilum on trout aquaculture
Flavobacterium psychrophilum is a Gram-negative, rod-shaped, 1 proteolytic bacterium 2 that has a global distribution in freshwater aquaculture. It is found in cold, fresh waters with an optimal growth temperature below 16°C.3,4 F. psychrophilum is a bacterial pathogen that causes rainbow trout fry syndrome (RTFS) and bacterial cold water disease in salmonids. 5 RTFS can result in high rates of juvenile mortality, leading to losses reaching up to 60% in severe cases. 4 Infected fry display symptoms such as inappetence, partial dark skin coloration, 6 exophthalmia, 7 anemia, ascites, and vertebral deformities. 1 In older fish, bacteria colonize the skin and gills, leading to the formation of necrotic lesions and, ultimately, death in ∼20% of cases. 8 Alternatively, it may lead to the development of spinal deformities and abnormal swimming behavior in surviving fish. 9 Historically, the first isolation of F. psychrophilum was done in the United States in 1960, 10 and isolation of F. psychrophilum as a disease-causing agent in fish farms has been reported since the 1980s in Asia, Australia, North America, South America, and several European countries. 11
Phage-based pathogen control challenges
Flavobacterium psychrophilum infections require high labor costs for treatment and expenditure on chemotherapy. The undeveloped immune system poses challenges to preventive immunization during the early farming stages, 12 and antibiotics to control the disease are becoming less effective due to the emergence of antibiotic-resistant bacterial strains. 11 In this context, phages have been suggested as a promising alternative to antibiotics for treating and preventing F. psychrophilum outbreaks in aquaculture.13,14
Phage libraries are vital for the development of efficient phage-based applications and to progress our understanding of phage–host interactions. Well-assorted collections of phenotypically and genomically characterized representatives of phages from natural communities can reveal patterns in the distribution of phage phenotypes and genomic features between and within taxonomic groups. 15 These collections can also suggest the molecular basis of receptor specificity in various phage groups, thereby expanding our understanding of bacteriophage ecology, evolution, and their interactions with bacterial immunity systems, while enhancing the selection process for therapeutic applications. 16 Phage libraries need to be continuously expanded and updated to keep up with the rapid phage–host coevolution in natural communities.17,18
Modern phage isolation: Scaled-down innovations of traditional techniques
The double agar overlay assay, 19 hereby referred to as direct plating, is the most common method for detecting infectious bacteriophages. This method facilitates the identification and isolation of individual phages from plaques formed in lawns on single host cultures on agar plates. This simple method can be implemented in any laboratory with minimal investment costs, making it applicable to any plaque-forming phage infecting a culturable, lawn-forming host. 20 This method is often used for phage enumeration, in experimental conditions, of phage lysates or high-phage titer environmental samples such as wastewater samples.21,22 However, the assay can be time-consuming and may exhibit high variability if not well standardized. 23 This method is also not suitable for the isolation of phages from environmental water samples with lower titers, without prior concentration of the phages, as the volume that can be screened on each replicate plate is quite limited. 24 As phages isolated for therapeutic purposes must be adaptable to culture-based methods, the double agar overlay remains essential for detecting infective phage units.
To overcome the challenge of low phage concentrations in natural samples, larger sample volumes can be enriched with the target host strain to increase the probability of phage infection and propagation prior to direct plating to detect the phages.20,24 Alternatively, the efficiency of phage isolation can be increased by up-concentration of phages in water samples using various approaches, such as ultrafiltration, ultracentrifugation, or phage precipitation,25–27 followed by direct plating. These concentration approaches require large sample volumes and the extensive use of materials, time, and space. Adaptations of the methods to reduce the need for extensive resources have emerged in recent years, mainly targeting human pathogens in wastewater. This includes a 96-well assay for isolating phages against Salmonella enterica, Escherichia coli, Enterococcus faecalis, or Pseudomonas aeruginosa in wastewater, 28 which was later modified successfully for isolating phages infecting Klebsiella pneumoniae and Enterobacter cloacae from wastewater. 29
While high-throughput phage isolation methods have been developed for isolating phages from high-density phage environments such as wastewater, there is a need for efficient methods aiming at phage isolation from environments with low phage concentrations. In 2019, Rajnovic et al. 30 proposed an optical density-based approach for the detection and quantification of bacteriophages. The method was developed by studying the absorbance over time of phage-infected bacterial cultures, as a function of both phage abundance and initial host cell concentration, utilizing Escherichia coli phage T4 and its host, Escherichia coli DSM 613 (DSMZ, Germany). This technique demonstrated the ability to detect as few as 10 initial phages per 220 µL by monitoring optical density kinetics. The sensitivity of this approach was further validated by Doss et al. in 2022. 31 In this study, we tested a series of approaches for optimizing the isolation efficiency of phages against F. psychrophilum in a fish farm environment, by quantification of phage recovery rate and detection limit using different concentration methods and enrichment assays. By combining phage enrichment of host bacterial cultures with optical density kinetics, we developed a sensitive method for isolating F. psychrophilum phages from environmental samples using a multiwell plate assay. The method’s high sensitivity and efficiency are demonstrated through the simultaneous screening of numerous water samples from a 12-month sampling effort from a rainbow trout (Oncorhynchus mykiss) recirculating aquaculture system (RAS), using single bacterial hosts.
Materials and Methods
Bacteriophages, bacterial isolates, and water samples
Phage FpV-4 13 was used for the assessment of phage recovery and detection limit using different isolation and concentration methods. The phage stock was stored at −80°C with SM buffer (8 mM MgSO4, 99 mM NaCl, 50 mM Tris-Cl (pH 7.5), 0.01% gelatin) and glycerol (15%). F. psychrophilum strain 950106-1/1 (Supplementary Table S1) 32 was used as phage FpV-4 proliferation and quantification host.
Following the method assessment study, the optimized high-throughput method was applied in a long-term phage isolation effort in a fish farm facility. Here, 59 F. psychrophilum isolates (Supplementary Table S2) were used as potential hosts for isolating F. psychrophilum phages from trout farm water samples. All the bacterial strains used in this study were preserved at −80°C in tryptone-yeast extract-salts broth (TYES-B) 1 with 20% glycerol. Before use in the phage isolation experiments, the bacterial hosts were taken from freezer stocks, spread on TYES-A (tryptone 0.4%, yeast extract 0.04%, CaCl2 × 2H2O 0.05%, and MgSO4 × 7H2O 0.05% with 1.1% agar), and incubated at 15°C for 3–5 days. 13 A single colony was inoculated into TYES broth, incubated in the dark at 15°C with 100 rpm shaking, and grown to an optical density (OD) of 0.2–0.9 for the double agar overlay assays, and to an OD of 0.01–0.03 for the 96-well plate enrichment assay.
Water samples were collected from two RAS facilities rearing rainbow trout in Denmark, both experiencing recurrent F. psychrophilum infections. The first RAS facility was sampled once during August 2017. Water from this facility was collected in 1 L brown high-density polyethylene (HDPE) bottles (Nalgene™, Thermo Fisher ScientificTM, Rochester, USA) and filtered (0.22 µm, polycarbonate, GE Water and Process Technologies, Pennsylvania, USA). The water was stored at 4°C in the dark before being used for phage detection and up-concentration methods described below (Assessing protocols for phage detection: Recovery efficiency and detection limits).
The second RAS facility was sampled over a 12-month period (July 2023–July 2024). A total of 45 water samples were collected at six time points from different locations within the facility, including the hatchery tanks, rearing tanks, outflow water, biofilter, and sludge collection. The water was collected in 1 L brown HDPE bottles and filtered using glass fiber filters (Whatman) to remove solid particles, followed by filtration through 0.22 µm polyethersulfone membrane pressure filters (Millipore, Germany) into sterile 1-L HDPE bottles. The filtered samples were stored at 4°C in the dark and used as described below (Application of high-throughput detection method on water samples). A subsample of the rearing tank water collected in July 2023 was autoclaved after filtration for use in optimization experiments (High throughput method assessment).
Fifty-nine F. psychrophilum isolates were isolated from fish from the second RAS facility and identified using Matrix-Assisted Laser Desorption Ionization Time-Of-Flight mass spectrometry. 33 These isolates were subsequently used for the single-host enrichments.
Assessing protocols for phage detection: Recovery efficiency and detection limits
To increase the chance of phage isolation from environmental water samples, the phage size fraction in water can be concentrated or exposed to potential host populations. We compared different phage concentration methods, such as FeCl3 precipitation, ultracentrifugation, Amicon filtration, and polyethylene glycol (PEG) precipitation, and single-strain enrichment approaches for the quantification of recovery efficiency and detection limit for F. psychrophilum phages in water samples.
To assess phage recovery and detection limits, the phage concentration methods were compared using sterile-filtered RAS water. Infective phages were not naturally present in this water, and the samples were spiked with phage FpV-4 to obtain five final concentrations: ≈ 100 PFU/mL, ≈ 10 PFU/mL, ≈ 1 PFU/mL, ≈ 0.1 PFU/mL, and ≈ 0.01 PFU/mL, with a total of 10 L of each dilution (Table 1). The exact concentrations of PFUs in the spiked water were measured using the TYES-soft agar (0.4% agar) overlay method (plaque assay 34 ) with some modifications. 800 µL of mid-log-phase bacterial culture (OD = 0.5) was mixed with 500 µL of FpV-4 spiked water and incubated for 30 min to allow phage adsorption to the hosts. Then, 8 mL of 42.5°C, 0.4% molten TYES soft agar was added, poured on square TYES-A plates, and incubated at 16°C for 5 days. The presence of infective phage was counted as plaques in the bacterial lawn, and the actual phage concentrations were calculated (Table 1). Since the amount of phage in some water treatments was very low (<1 PFU/mL), it was not possible to titer the actual phage concentration of the spiked solution, and these concentrations were estimated from the dilutions of the phage stock. However, to maintain clarity of expression, the theoretical phage concentrations will be used throughout the text, and the corresponding concentrations based on the PFU enumeration of each phage dilution will be reported in the results section. The 10 L phage-spiked water samples for each dilution were then processed using a selection of protocols to assess their efficiency in concentrating and detecting the phages.
Results from Phage Recovery Experiment with a Range of Initial Phage Concentrations
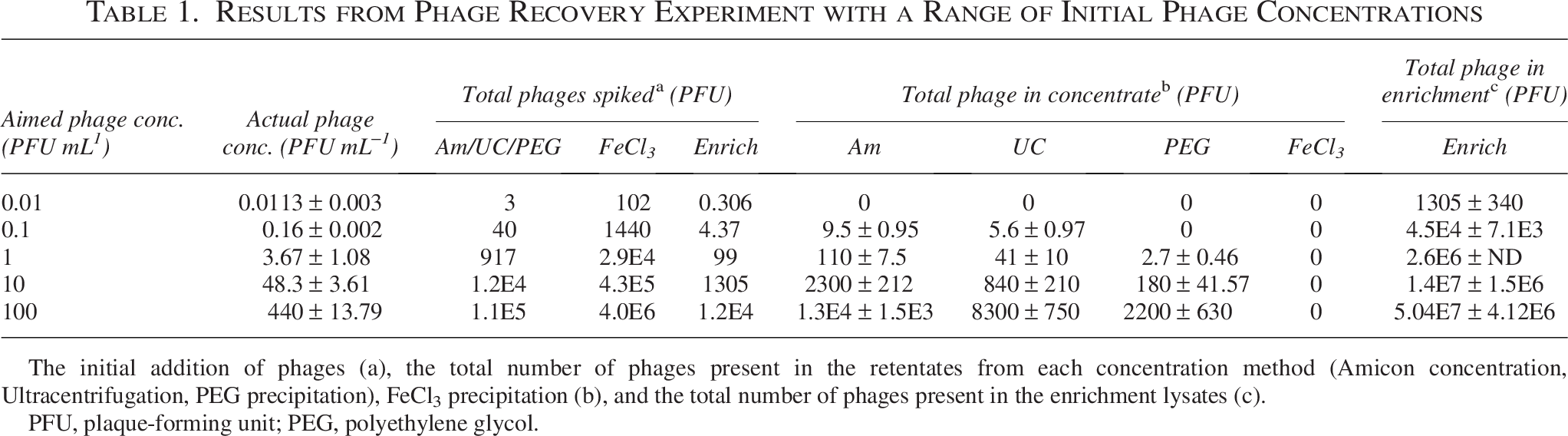
The initial addition of phages (a), the total number of phages present in the retentates from each concentration method (Amicon concentration, Ultracentrifugation, PEG precipitation), FeCl3 precipitation (b), and the total number of phages present in the enrichment lysates (c).
PFU, plaque-forming unit; PEG, polyethylene glycol.
PEG precipitation
PEG is used to precipitate phages for concentration and purification purposes. The precipitation was conducted following the protocol described by Nissimov, 35 with some modifications. Briefly, 14.61 g NaCl was added to 250 mL phage dilutions (final concentration 1M), then the salt/sample solution was incubated at 4°C overnight and centrifuged at 5000 g for 1 h. The supernatant was transferred to sterile 50 mL conical centrifugation tubes, and PEG was added to a final concentration of 10% (w/vol). The solution was incubated overnight at 4°C and centrifuged at 10,000 g for 50 min. The supernatant was discarded, and the tubes were inverted for a few minutes to dry. The pellet was resuspended in ∼800 µL SM buffer. 100 µL of concentrated water sample was serially diluted, mixed with 400 µL (OD = 0.5) of host strain, incubated at 15°C for 30 min with agitation, mixed with 4 mL soft agar, and poured on TYES-A plates. Plaques were counted after 5 days of incubation at 15°C.
FeCl3 precipitation
About 0.9 mL Fe(III)Cl3 working stock solution (10 g L−1 Fe) was added to 9 L of each phage dilution in 10 L carboys, followed by 1 min vigorous shaking and overnight incubation at 4°C in the dark to allow precipitation. Using a peristaltic pump, with a 15 psi pressure gauge, the samples were then filtered through GE Water Systems Polycarbonate membranes (cat.no. K10CP14220; 142 mm, 1.0 µm) on top of Pall Supor-800 support filters (cat.no.60114; 142 mm, 0.8um) in a stainless-steel filtration system. The filters containing the FeCl3 precipitate were resuspended in 10 mL of freshly prepared 0.2M Ascorbate-0.1M Ethylenediaminetetraacetic acid (EDTA)-Mg buffer at pH 6 to 7. 36 For viable phage detection and quantification, 600 µL of each concentrated spiked water sample was used.
Ultracentrifugation
A 250 mL of 0.2 µm phage-spiked water samples were distributed in polyallomer tubes and centrifuged in a swinging bucket rotor at 100.000 g (Optima LE-80K Ultracentrifuge, Beckman, USA) for 1 h to pellet the phages. Ten rounds of centrifugation were required to centrifuge the entire sample volume. The pellets were resuspended in 500 µL SM buffer. The concentrated phage dilutions were serially 10-fold diluted in sterile SM buffer, and the subsequent plaque assay method was used to screen for phages.
Amicon concentration
250 mL of each phage in facility water dilution was divided into 15 mL 30 kD Amicon concentration devices (Millipore, Germany) and centrifuged at 3000 g for ca 20 min to concentrate the sample above the membrane to a volume of ≈ 500 µL. The retentates from each tube were combined and used in subsequent direct plating assays to screen for phages.
Single-strain enrichment
Enrichment cultures for phage proliferation were set up with each phage dilution. Three milliliter of 10× TYES-B was mixed with 27 mL of each dilution of the phage-spiked water sample inoculated with 1 mL (OD = 0.5) of host strain 950106-1/1. The cultures were incubated for 5 days in the dark at 15°C with agitation to allow for the propagation of phage FpV-4. After incubation, the enrichment cultures were centrifuged at 5000 × g for 10 min at 4°C and filtered through 0.2 µm polycarbonate syringe filters. Triplicate 10-fold serial dilutions of the phage solutions were prepared, and the infective phages were quantified using the soft agar overlay method.
High-throughput method assessment
Based on the comparison of different concentration methods, it was clear that the probability of detecting a single phage unit is higher when its concentration is increased through propagation in enrichment cultures than by using any of the other phage concentration protocols. The detection limit suggested that the enrichment method is sensitive enough to isolate a single phage unit in a 27 mL water sample. To optimize this approach and design it for more high-throughput screening, we scaled down the enrichment culture volumes from 30 mL to 230 µL each, in 96-well plates. The efficiency of this phage isolation method was tested for three different phage-host combinations (Supplementary Fig. S1). F. psychrophilum strains 950106-1/1, FPS-S6, and FPS-D27 were combined with phages FpV-4, FPSV-D22, and FPSV-D45 (Supplementary Table S3), respectively, to examine whether the detection limit and phage propagation efficiency varied among phage-host systems. Using a dilution series of known phage concentrations in enrichment cultures with specific hosts allowed the determination of phage detection limits for each phage–host system.
To test and optimize the method under conditions closely resembling the intended application of isolating from water samples, the 10-fold serial dilution of each phage stock was prepared using 0.2 µm filtered and autoclaved water from the rainbow trout RAS. 500 µL of each phage dilution was tested using the soft overlay method to estimate the actual phage concentration.
The bacterial host strains were grown from −80°C stocks in 5 mL of TYES-B for 12 h at 15°C with 100 rpm shaking. A negative control and three phage dilutions were tested in every experiment, preparing a minimum of 8 replicates for each host strain. For each well, 200 µL of known phage concentration was mixed with 20 µL of 10× TYES-B and 10 µL of OD: 0.01–0.03 host broth culture in transparent, U-bottom, 96-well plates (Greiner Bio-One, Austria). The initial host OD of 0.01–0.03 was based on preliminary tests, which showed the best phage inhibition at low initial host density. The plates were incubated at 15°C, with pulsed shaking between readings, in a Varioskan LUX Multimode Microplate Reader (Thermo Scientific, Massachusetts, USA). OD was recorded at 3-h intervals over a 4-day period. After the 4-day incubation, the final concentration of infective phages was quantified in all wells using the spot assay method on square plates. A tube for each host was filled with 10 mL melted TYES top agar (0.4%). After the top agar (TYES 0.4% agar) was cooled down in a water bath to 42°C, 1 mL of OD 0.2–0.6 of host strain liquid culture was added to the tube. The 11 mL of top agar and host strain mix were shaken and poured on previously solidified TYES-A on each 120 × 120 mm square plate. Using a multichannel pipette, 5 µL from each well was spotted on the square soft agar overlay plate with the embedded enrichment host to identify and measure the number of wells positive for phage propagation activity. After the first spotting, 100 µL from each replicate well of the same initial phage concentration was pooled, mixed, and serially diluted to estimate the average final phage concentration using a plaque assay. The plates were incubated in the dark at 15°C for 5 days. These estimates of final PFU were combined with the OD measurement to examine the relationship between bacterial host growth inhibition and phage propagation as a function of initial phage density.
Application of high-throughput detection method on water samples
Following the first sampling, in July 2023, 9 samples and 15 F. psychrophilum strains were used for a screening performance comparison with direct plating (plaque assay). We then employed the 96-well enrichment assay described above as a standardized approach for the isolation of F. psychrophilum phages from aquaculture samples during a temporal study.
For each assay, 200 µL of the filtered aquaculture water samples were mixed with 20 µL of 10× TYES-B and 10 µL of a 24-h host broth culture (OD 0.01–0.03) in transparent, flat-bottomed 96-well plates (Greiner Bio-One, Austria) (Fig. 1). For each plate, up to 10 different water samples, each distributed in eight replicate wells, were screened for phages against each of the F. psychrophilum pathogen strains isolated at the given sampling date. The total number of plates, therefore, varied depending on the number of pathogens isolated at each sampling date. For sampling timepoints where no bacterial strains were isolated (May 2024) or very little phage activity was detected using the corresponding host isolates (March 2024), a selection of other previously isolated F. psychrophilum strains from the same RAS facility was chosen. The plates were sealed with parafilm and incubated at 15°C in the dark with no shaking for 4–7 days.
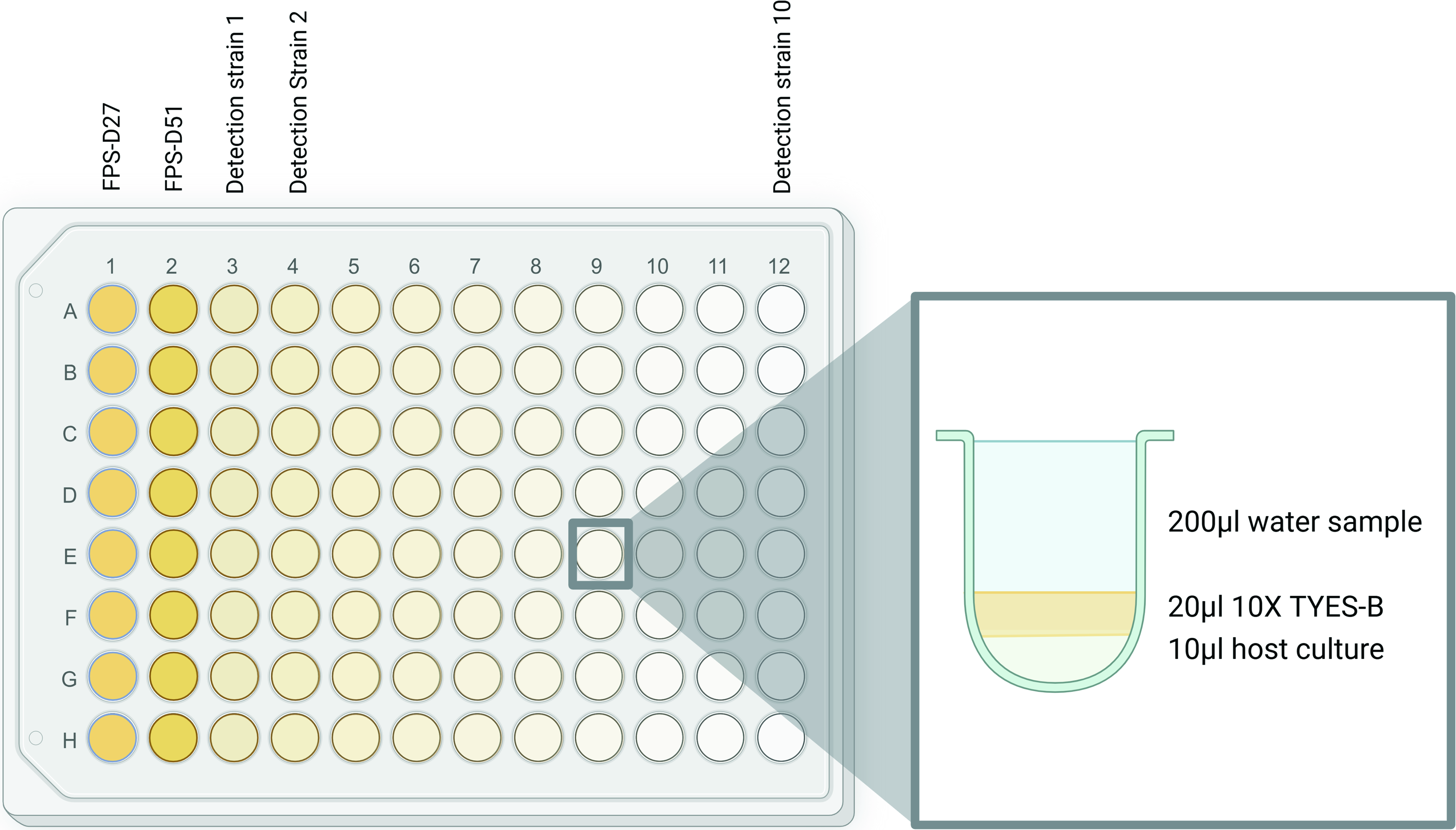
General 96-well plate enrichment assay sample distribution setup, showing the content of the individual wells.
Detection of phage activity in the water samples
After the enrichment, the presence of phage activity was determined for each well. As described above, soft overlay square plates were prepared to screen for the presence of phages in the wells. Using a multichannel pipette, 5 µL from each well was spotted on the soft agar overlay with the embedded enrichment host. The plates were incubated in the dark at 15°C for 2–5 days, until the bacterial lawn had formed. Following the incubation period, the plates were inspected for signs of phage activity (inhibition zones in the bacterial lawn). The presence of phages in inhibition zones was confirmed using plaque assays. In total, 45 collected water samples were combined with the 59 host strains, resulting in 538 water sample-single host strain screening combinations for the detection of phage activity.
Phage isolation
To confirm that pathogen inhibition was caused by phage activity and to isolate individual phages Supplementary matrix), selected inhibition zones were picked using a sterile pipette tip and eluted in 400 µL of SM buffer. The eluted spot was diluted 1:10–1:1000 in SM buffer. One part of host culture (48–72 h, OD 0.2–0.9) corresponding to the plates showing growth inhibition was added to one part of each dilution, and the mixture was incubated at 4°C for 30–60 min before being added (300 µL) to 4 mL of molten top agar (43°C). The top agar mixture was briefly vortexed and poured onto a room temperature underlay TYES agar plate. Once the top layer of agar surface had solidified, the plates were incubated at 15°C for 48–72 h. Following incubation, the presence of plaques was noted. Based on differences in plaque morphology, single plaques were selected to proceed with a minimum of three plaque purification and reinfection rounds. 37
Results
Assessment of concentration and enrichment protocols—recovery efficiency
Using known concentrations of a specific phage, the performance of four different methods for quantifying F. psychrophilum phage recovery and detection limit in natural samples was tested: ultracentrifugation, Amicon concentration, FeCl3 precipitation, and PEG precipitation.
Filtered RAS facility water was spiked with F. psychrophilum phage FpV-4 to prepare a concentration series ranging from ∼0.01 to ∼100 PFU/mL, corresponding to a total number of phages that ranged from ∼3 PFUs (highest dilution) to 1.1 × 105 PFUs (lowest dilution) in the 250 mL phage dilutions that were concentrated by ultracentrifugation, Amicon filtration, and PEG precipitation (Table 1). The enrichment cultures, which contained 27 mL of the phage dilutions, thus contained from 1.13 phage (highest dilution) to 4.4 × 104 phages (lowest dilution), whereas in the FeCl3 precipitation using 9 L of water, the total phage numbers ranged from 102 phages (highest dilution) to 4 × 106 phages (lowest dilution) (Table 1).
The quantification of the specific phage retentates revealed large differences in the ability of the methods to concentrate phages (Table 1). Amicon and ultracentrifugation achieved the highest phage concentrations in the retentates, while FeCl3 precipitation was unsuccessful in concentrating viable phages across the entire dilution series, despite using higher volumes. These numbers were converted into average concentration factors for each method, ranging from a maximum of 66 for the Amicon filtration to a minimum of 4 for the PEG precipitation (Table 2).
Average Concentration Factors and Phage Detection Limits for Each Method Tested in the Study
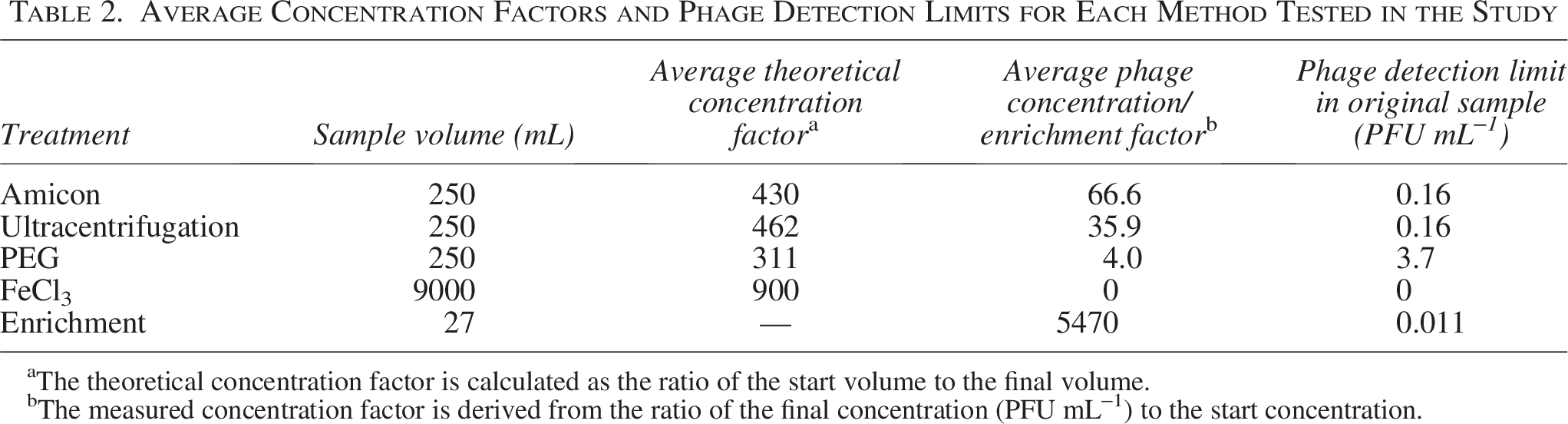
The theoretical concentration factor is calculated as the ratio of the start volume to the final volume.
The measured concentration factor is derived from the ratio of the final concentration (PFU mL−1) to the start concentration.
All the concentration methods, therefore, resulted in a relatively high loss of phages during the concentration process, with the maximum recovery efficiency of 24% (Fig. 2). Amicon filtration was the most efficient concentration method, with a recovery rate of 0.10–0.24, whereas phage recovery ratios from ultracentrifugation and PEG precipitations were only 0.04–0.14 and 0.003–0.02, respectively. It was not possible to recover phages from samples with an initial phage concentration of 0.01 PFU/mL with any of the concentration methods, and the minimum phage concentration that resulted in phage recovery by PEG precipitation was 1 PFU/mL (Fig. 2).
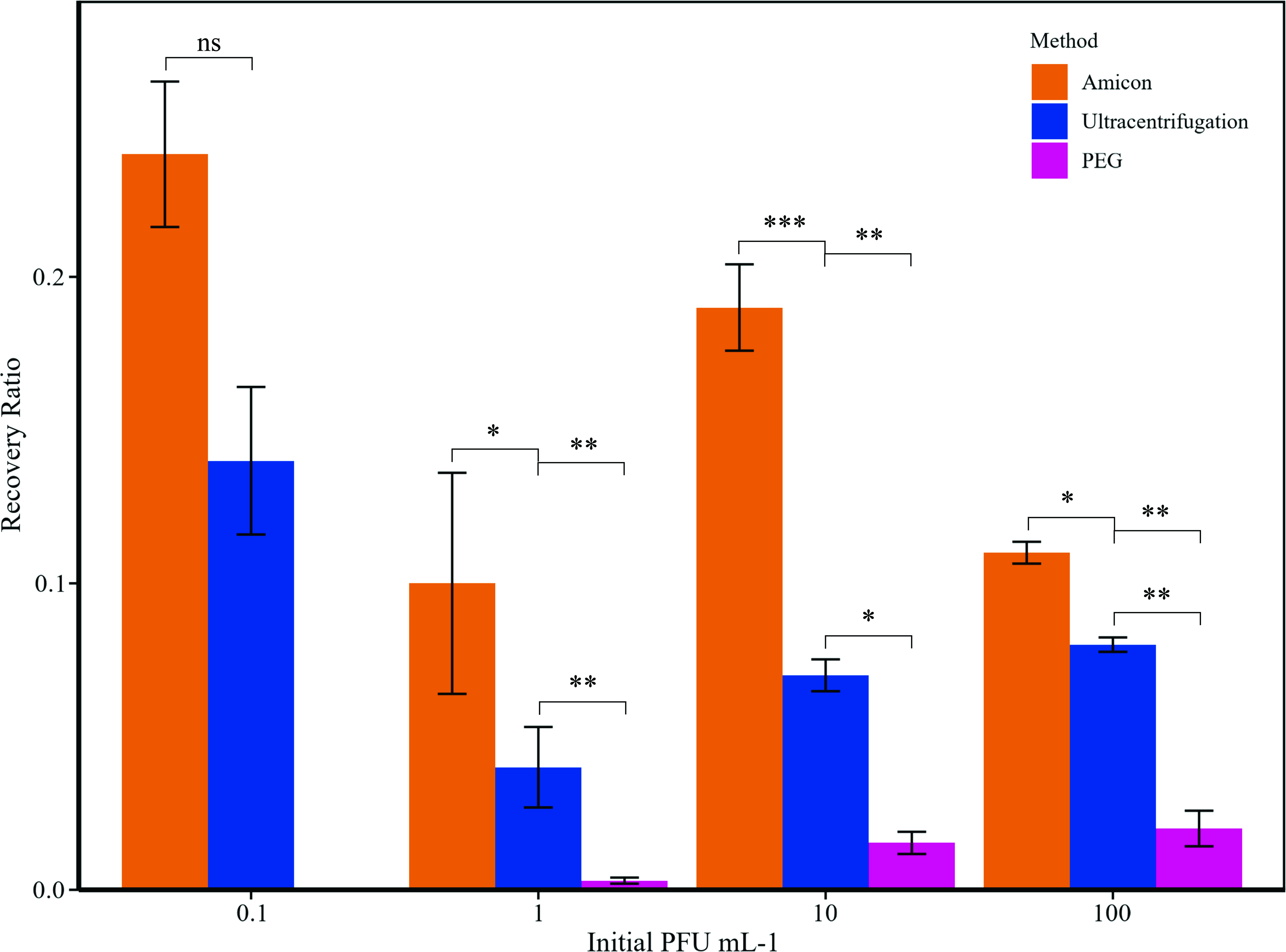
Recovery ratio in relation to the initial theoretical FpV-4 concentrations, for Amicon concentration, ultracentrifugation, and PEG precipitation. FeCl3 precipitation is not represented as the recovery rate resulted in zero for all the concentrations tested. PEG, polyethylene glycol.
The initial phage concentration in the water did not significantly affect the recovery rate, except in PEG precipitation, where increasing the initial phage concentration enhanced the recovery efficiency (Fig. 2). Enrichment cultures, where phage dilutions were incubated with a susceptible host strain, proved to be more efficient than any of the concentration methods in increasing phage abundance for subsequent isolation and resulted in significant phage propagation even at the highest phage dilution (0.01 PFU/mL) (Table 1, c).
Based on the highest phage dilution with detectable PFUs in the concentrate and the concentration factor, we could estimate the detection limit of each method (Table 2). For the ultracentrifugation and Amicon filtration approaches, the detection limit of the methods was 1.6 × 10−1 PFU/mL, whereas the lower concentration factor for the PEG precipitation resulted in a higher detection limit of 3.7 PFU/mL. Enrichment culture, with no previous concentration of the phage dilution, was by far the most sensitive method for phage isolation with a phage detection limit of 1.13 × 10−2 PFU/mL, corresponding to the detection and propagation of 1 phage in a ∼90 mL water sample.
Optimization of enrichment culture—determination of the detection limit
Based on the method assessment results, the enrichment method was then further optimized and adapted into a high-throughput approach. Using 200 µL enrichment cultures in 96-well plates, we tested different phage concentrations and phage–host systems to investigate the sensitivity of this modified isolation approach. The different concentrations tested of phages FpV-4, FPSV-D22, and FPSV-D45 ranged from 1 to 700 PFU mL−1 but varied between the parallel systems due to the difficulty of replicating the exact same titers starting from different phage stocks.
To quantify the effect of initial phage concentration on host cell density and phage propagation, in the 4-day incubations, the ratio between OD in phage samples (ODs) and control cultures without phages (ODc) was calculated at the end of the experiment, across a range of initial phage concentrations, in different phage–host systems.
A higher initial phage concentration was associated with stronger bacterial growth inhibition for all the different hosts, resulting in a lower sample-OD (containg phages) to control-OD ratio (without phages (ODs/ODc) (Fig. 3). However, at the lowest initial phage concentrations (<10 PFU well−1), the reduction in final OD was not significantly different from the controls, despite a phage propagation of up to 106 PFU mL−1 and 109 for FpV-4 and FPSV-D22, respectively, over the 4 days. The minimum initial phage concentration that showed a detectable OD inhibition varied between the phage-host systems. Phage FpV-4 on F. psychrophilum strain 950106-1/1 and phage FPSV-D22 on strain FPS-S6 showed consistent host growth inhibition starting from 10 PFUs per well, whereas no significant changes in optical density measurements were observed for phage FPSV-D45 using host strain FPS-D27 for initial phage units < 250 PFU/mL.
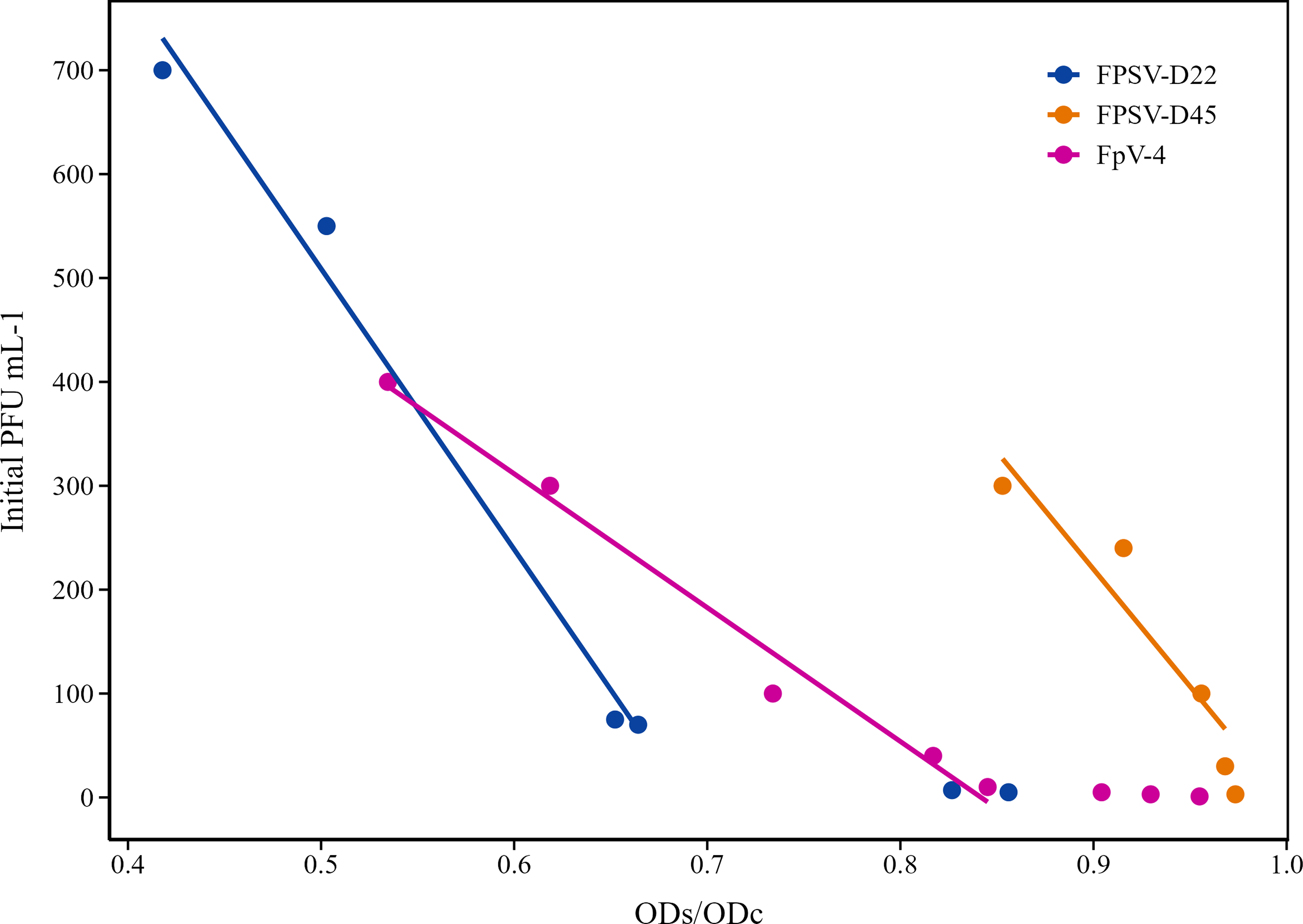
Detectable OD inhibition for each of the different phage–host systems tested, represented as sample OD to control OD ratio (ODs/ODc) in relation to various initial PFU mL−1 and the relative final PFU mL−1 after 93 h of small-volume enrichment. All the initial PFU mL−1 concentrations tested for each system are represented in the figure. The linear regression is based only on initial PFU/mL−1 concentrations ≥10. The R2 values for the systems FPSV-D22, FPSV-D45, and FpV-4 are, respectively, 0.99, 0.98, and 0.89. PFU, plaque-forming unit
In low-phage-titer treatments where the optical density failed to show a difference between phage treatment and control, spot assays were used to identify phage propagation in the wells at all initial phage concentrations. The sensitivity of the small-volume enrichment approach was determined at a single PFU per well.
The significant linear correlation between initial phage concentration and the OD inhibition ratio suggests that PFU of a specific phage in a water sample can be predicted from OD curves, even at very low concentrations, allowing for faster phage quantification.
Application of high-throughput detection method on water samples from RAS facility
The year-long sampling effort at the Danish RAS facility yielded a total of 45 water samples from different farm locations (13 biofilter samples, 11 tank water samples, 11 outflow samples, 6 sludge samples, 4 hatchery samples) tested against 59 F. psychrophilum strains using the high-throughput, low-volume isolation approach. For the water samples, growth inhibition on the bacterial lawn after enrichment was observed in 36 out of the 45 samples and on 35 of the 59 F. psychrophilum strains tested (Supplementary Table S2). The samples that showed the highest phage detectability were tank and outflow water, which showed phage activity in over 90% of the screened samples. Phage activity was also often present in the biofilter samples, with 85% showing host inhibition. The sludge and hatchery samples exhibited the least phage activity overall, with no inhibition observed in any of the tested hosts in 50% of the samples.
The performance comparison between direct plating and the 96-well enrichment assay, using RAS facility water from the first sampling (July 2023), resulted in 37 different host-sample combinations confirmed to be positive for phage activity. Of these, 21 were only detected using the 96-well enrichment assay (Supplementary Table S4).
The growth inhibition of the water samples was confirmed to be phage activity on 40 inhibition zones across all six sampling timepoints. In only one case, the growth inhibition observed in the outflow water sample W5-7 on the F. psychrophilum strain FPS-D65 could not be confirmed to be phage activity due to the lack of plaque formation. The plaques were used to isolate and purify phages for further characterization (Landor et al., unpublished).
Discussion
Assessment of phage isolation methods
Efficient phage isolation from samples with low PFU concentrations is crucial for expanding phage collections. Concentrating phages in water samples is a standard way to increase phage densities, thereby increasing DNA yield prior to extraction and improving phage isolation efficiency.36,38 However, in the present study, it was shown that a fraction of the phages was lost or inactivated in the up-concentration process. By comparing phage recovery from volumes ranging from 250 mL to 9 L using five phage concentration approaches, we found that even in the best-performing concentration approach (Amicon), the majority of the infective phages (>75%) were lost in the process (Table 1). Ultracentrifugation and PEG precipitation of the phages showed even lower recovery, and no viable phages were obtained from FeCl3 precipitation. The poor recovery of infective phages suggests that the physical and chemical treatment of the phages during the concentration process strongly affected their infectivity and/or resulted in the loss of phage particles.
Previous work on the concentration of viruses from seawater using filtration approaches 26 has found that water samples with a high presence of organic matter face a higher risk of clogging the filter membrane, resulting in the loss of viral particles during the filtration process.
Colombet et al. 25 found that PEG concentration resulted in more than a 2-fold increase in viral particle recovery efficiency compared with ultracentrifugation. The reduced efficiency observed with ultracentrifugation was likely attributable to the high centrifugal forces, which may have caused structural disruption of viral particles. However, it has also been shown that PEG precipitation is not efficient for phage concentrations <105 PFU/mL, 39 suggesting that the recovery of phages at low concentrations (<10 PFU/mL) found in this study was surprisingly high.
Besides the iron chloride precipitation approach, all the phage concentration methods resulted in a higher phage concentration in the concentrate, ranging from 4 to 67-fold increases in phage titer, emphasizing their potential for increasing the chance of phage isolation in subsequent isolation procedures. However, the >5400-fold increase in phage concentration during incubation in enrichment cultures demonstrated the efficiency of increasing phage titers against known hosts.
The translation of this result into a high-throughput method therefore led to a setup that enables the simultaneous testing of multiple combinations of water samples and potential host strains in relatively low volumes. Before spotting the single-well enrichments, neither filtration nor chloroform treatment is necessary, making the method more cost-effective and efficient. Additionally, this method’s liquid-based cultivation allows for the use of more fastidious strains that do not perform well as hosts in the soft agar overlay method. The enrichment increases the phage concentration, thereby boosting the chances of observing an inhibition zone following spot assay when the plaques are hardly visible due to poor bacterial lawn quality.
The method was tested using known concentrations of infective F. psychrophilum phages to determine its ability to detect phages at low concentrations and to predict phage concentrations in the water samples based on the reduction in OD during incubation. Distinct phage–host combinations exhibited significant, unique relationships between OD reduction and phage concentration. Depending on the specific phage–host system, propagation starting from low concentrations (<10 PFU/mL−1) can occur without OD reduction. However, when starting from a concentration of 10 PFU/mL, regression analysis revealed a linear correlation for each phage–host system, where the OD-reduction could explain between 89% and 98% of the variability in initial phage concentration. The specificity observed for each system emphasizes that the model cannot be directly applied to quantify the concentration of F. psychrophilum phages in environmental samples due to phage diversity and the varying dynamics of unknown phage-host interactions. Nonetheless, combining the 96-well plate enrichment assay with optical density kinetics offers a promising alternative for estimating phage concentration during propagation processes, requiring less material and waiting time than traditional plaque counting based on double agar overlays.
High-throughput detection method: Application in temporal sampling in aquaculture facility
Following the performance assessment, the method was applied during an annual sampling effort in a fish farm facility with samples from various locations within the farm, including the hatchery, rearing tanks, treatment tanks, tank outflow water, biofilter tanks, and sludge tanks. The low phage isolation (<50%) in the sludge samples was likely caused by contained high levels of suspended material, which clogged the glass fiber filters, potentially retaining the phages on the particles on the filters. The low phage recovery from the hatchery water suggests that the pathogen host only reaches high densities in the facility when the fish enter the fry stage, in accordance with its infection pattern.40,41
The method was superior in isolating phages (10-fold more phages isolated) than using direct plating, emphasizing the low detection limit demonstrated in the method assessment.
The high-throughput detection method, proven successful in isolating phages from water samples with concentrations as low as 0.01 PFU mL−1, does not guarantee enrichment from a single phage particle in each well. Therefore, repeated rounds of single plaque selection are required to ensure phage isolation. Hoshino et al. 42 developed a method based on the isolation of single E. coli phage particles, followed by upscaled repropagation with simultaneous enumeration. The characteristic compartmentalization and many replicates for each sample of the current method eliminates the need to conduct numerous rounds of plaque assays to fully isolate phage particles. If adapted to Flavobacterium, this method could shorten the phage isolation process.
With the setup used during this study, the optical density approach was generally not sensitive enough to detect low initial phage concentrations (<10 PFU well−1), as the reduction in OD from phage propagation during the incubation of such low initial phage concentrations was insignificant relative to a control without phage. However, for concentrations of >10 PFU well−1 typically used for propagation, the OD inhibition proved to be consistent (Fig. 3) and a predictor of initial phage concentration as well as phage propagation (Supplementary Fig. S2) for specific phage–host systems. Therefore, by recording phage–host interactions at different phage concentrations, it may be possible to implement the method to estimate the abundance of phage in a sample at a specific enrichment time point. This could be a valuable tool for characterizing and monitoring host strain-phage growth profiles during phage production, as suggested by Rajnovic et al. 30
PFU detection for phage therapy depends on culture-based methods, as the isolated phages must be active against the pathogenic strains of interest and adaptable to industrial-scale propagation and preservation. Culture-based methods are constrained by the adaptability of host strains and phages to the conditions set by the protocol in use, as well as the host’s sensitivity to the phage. The enrichment approach has been shown to be sensitive enough to detect a single PFU in a well. However, in a mixed phage community, phages with strong propagation characteristics are probably more likely to be detected using this method. Another limitation of this method is that phages can only be propagated and detected in combination with a sensitive host, making the selection of the bacterial strains crucial for the method’s success. To obtain a better understanding of the method’s ability to cover the overall diversity of F. psychrophilum phages in environmental water samples, it would be relevant to compare the isolation results with metagenomic analyses of the total genomic diversity of phage. Meanwhile, the method is routinely used for phage isolation against F. psychrophilum in aquaculture environments.
Conclusions
High-throughput screening methods are needed to boost phage isolation and identification. Tools that are efficient enough to keep up with the speed of phage–host coevolution are essential when the goal is to use natural phages to prevent and treat fish bacterial infections. To the best of our knowledge, this is the first study to propose a 96-well single F. psychrophilum phage particle isolation method. The method has now become an essential tool in our phage isolation workflow.
Footnotes
Acknowledgment
The authors thank the personelle at the DanAqua fish farm for help with sampling and Lone Madsen, Valentina Donati and Chiara Cialini from the Danish Technical University for providing F. psychrophilum strains used in the phage isolation effort.
Authors’ Contributions
Conceptualization: V.R., J.J., and M.M. Data curation: V.R. and L.L. Formal analysis: V.R., J.J., L.L., and M.M. Funding acquisition: M.M. Investigation: V.R., J.J., and L.L. Methodology: V.R., M.M., J.J., and L.L. Project administration: M.M. Resources: M.M. Supervision: M.M. Verification: V.R., and M.M. Visualization: V.R., and M.M. Writing–original draft: V.R. Writing–review and editing: V.R., M.M., L.L., and J.J.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by grants from European Union under the
Supplemental Material
Supplemental Material
Supplemental Material
Supplemental Material
Supplemental Material
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.