
Abstract
The aim of this study was to elucidate which of the following two factors plays a more important role in the localization of atherogenesis: the barrier function of the arterial endothelium modulated by wall shear stress or flow-dependent low-density lipoprotein (LDL) concentration at the blood/wall interface. To determine this, the rabbit aorto-iliac bifurcation was perfused with 3,3′-dioctadecylindocarbocyanine (DiI)-LDL solution under three different flow conditions: (i) forward flow (perfused in the in vivo flow direction); (ii) backward flow (perfused in a reversed flow direction); and (iii) static group (no flow). The results showed that there was a peak in the curve of DiI-LDL uptake distribution along the lateral wall of the bifurcation for all three groups, which was located in the branching areas where the endothelial cells were round and polygonal with no preferred orientation. Nevertheless, the peak of the forward flow group was much sharper than those of the other two groups. The overall DiI-LDL uptake was the highest for the static group. The present experimental study supports the concept that both the barrier function of the endothelium modulated by wall shear stress and the mass transport phenomenon of LDL concentration polarization are involved in the infiltration/accumulation of atherogenic lipids within the arterial wall. Nevertheless, the latter might play a larger role in the localization of atherogenesis.
Keywords
Introduction
Atherosclerosis is a degenerative disease that affects large arteries by progressive thickening and eventual hardening of the vessel wall through the formation of atheromatous plaques rich in cholesterol content. It has been well documented that atherosclerotic lesions in the arterial wall develop at certain sites such as the inner wall of curved segments and the outer walls of arterial bifurcations, 1–3 where the vessel walls are subjected to slow blood velocity with low wall shear stress as a result of flow disturbances. 2–5
It has been widely recognized that flow-induced shear stress plays a very important role in modulating endothelial functions and is one of the most important hemodynamic factors in atherosclerosis. 6 Nevertheless, due to the fact that the early event leading to the genesis of atherosclerosis is the accumulation of cholesterol and other lipids within the arterial wall, in recent years researchers have been paying more and more attention to material transport in the circulation and the interactions of blood cells with the blood vessel walls. 7 From the viewpoint of mass transport, the concepts of ‘residence time’ for atherogenic agents 8 and deposition of atherogenic particles onto the blood vessel walls 9 were proposed to account for the localization of atherogenesis.
Because the endothelium of arteries displays low permeability to plasma proteins, Deng et al. 10 have theoretically predicted a mass transport phenomenon of concentration polarization of atherogenic low-density lipoproteins (LDLs), with the LDLs increasing in concentration from bulk value towards interface within the arterial system and verified this experimentally in vitro. 11 Apparently, the occurrence of concentration polarization of LDLs in the arterial system can affect the ‘residence time’ and the deposition of atherogenic particles. They therefore have suggested that flow-dependent LDL concentration at the blood/wall interface may play an important role in the localization of atherogenesis.
On the other hand, it has been documented that abnormal wall shear stress in disturbed blood flow regions can alter the normal barrier function of arterial endothelium, leading to higher lipid infiltration into the arterial wall in these flow regions. 12–15
Our question is which of the aforementioned two mass transport explanations accounting for the localization of atherogenesis plays a more important role. In order to clarify this, we designed the present study using the rabbit aorta–iliac bifurcation as our model and compared the profiles of LDL uptake distribution along the bifurcation for three different experimental groups. We correlated the distributions of LDL uptake with the flow patterns and the morphological patterns of endothelial cells in the bifurcation. In this way, the role of endothelial cell barrier function to LDLs can be differentiated from the role of LDL concentration polarization in LDL uptakes.
Materials and methods
Preparation of LDLs and 3,3′-dioctadecylindocarbocyanine-LDLs
LDLs were prepared and purified from fresh plasma obtained from Beijing Red Cross Blood Centre by gradient ultracentrifugation according to the method of Redgrabe et al. 16 Then, the LDLs were filter sterilized through a 0.2-μm filter and dialyzed extensively against 0.9% saline and 0.01% EDTA. The purity of the LDLs prepared was over 90% (LDL/total proteins) as determined by the method of Lowry et al. 17 The LDLs were stored in sterile polystyrene tubes for periods less than five days at 4°C until ready for use, so that the level of oxidation of the LDLs was very low. Prior to each perfusion experiment, the purified LDLs were labeled with 3,3′-dioctadecylindocarbocyanine (DiI) (Biomedical Technologies Inc, Stoughton, MA, USA) following the procedure described by Li et al. 18 The labeled LDLs were added to Dulbecco's modified Eagle's medium (DMEM) cell culture medium that was used as the experimental perfusion solution. Then, the concentration of DiI-LDLs in the prepared perfusion solution was determined by measuring the protein concentration of the DiI-LDLs using the Bradford Protein Assay Kit (Sigma, St Louis, MO, USA).
Arteries preparation
Male New Zealand white rabbits weighing approximately 3.5 kg were obtained from the Laboratory Animal Center, Peking University (Beijing, China). The experiment followed a protocol approved by the institutional committee on animal use, and all animal care complied with the ‘Principles of Laboratory Animal Care’ and the ‘Guide for the Care and Use of Laboratory Animals’ (NIH Publication No. 86-23, revised 1985). Rabbits were anesthetized intravenously through the right marginal ear vein with a mixture of xylazine (2.2 mg/kg) and ketamine (22 mg/kg). The abdomen was opened with a midline incision and the aorta–iliac bifurcation was exposed and the surrounding tissue was gently removed. In order to minimize damage to the endothelium, we kept the vessel pressurized throughout the experiment by a reservoir placed 1.0 m above the artery and filled with Krebs solution (concentrations in mmol/L: NaCl, 118; KCl, 4.7; NaHCO3, 25; KH2PO4, 1.2; MgSO4, 1.2; CaCl2, 2.5; glucose, 11). The aorta was thoroughly examined for leakage in situ by pressurization of the vessel, and any points on the vessel that leaked were sealed by coagulation with an electrical fulgurator. The experimental artery section was flushed slowly to wash out any remaining blood. The cannulae were fixed rigidly onto a metal frame to hold the bifurcation in its in vivo shape and configuration. The cannulae were then advanced approximately 2 mm, into the vessel, enabling their tips to lay beyond any regions liable to have been damaged during the excision procedure. After a ligature was tied around each cannula as close to the tip as possible, the cannulated vessel was dissected from the body. During the entire process, Krebs solution was constantly applied to the outer surface of the vessel to prevent it from drying.
Perfusion solution
For each experiment, cell culture medium DMEM containing fetal calf serum (10%), penicillin (100 IU/mL) and streptomycin (100 μg/mL) (Sigma Chemical, St Louis, MO, USA) was freshly prepared as the experimental perfusion solution, in which DiI-LDL was added at a concentration of 10 μg/mL. The pH value of the perfusion solution was adjusted to 7.2.
Perfusion system
Figure 1 shows a schematic drawing of the experimental perfusion system, which consisted of a head tank, a downstream collecting reservoir, a peristaltic flow pump to circulate the perfusion fluid (DMEM, a cell culture medium) and a blender of air and CO2 with a constant temperature of 37 ± 1°C. All of the components of the perfusion system were connected using tygon tubing. The flow rate was controlled by adjusting the height of the overflow head tank and the resistance of the needle valve so that both the desired flow rate and perfusion pressure could be achieved simultaneously. A pressure transducer and a flow meter were used to monitor the perfusion pressure and the flow rate through the artery, respectively. The harvested bifurcation segment with the metal frame was immersed in Krebs solution and then connected horizontally to the overflow head tank perfusion system.
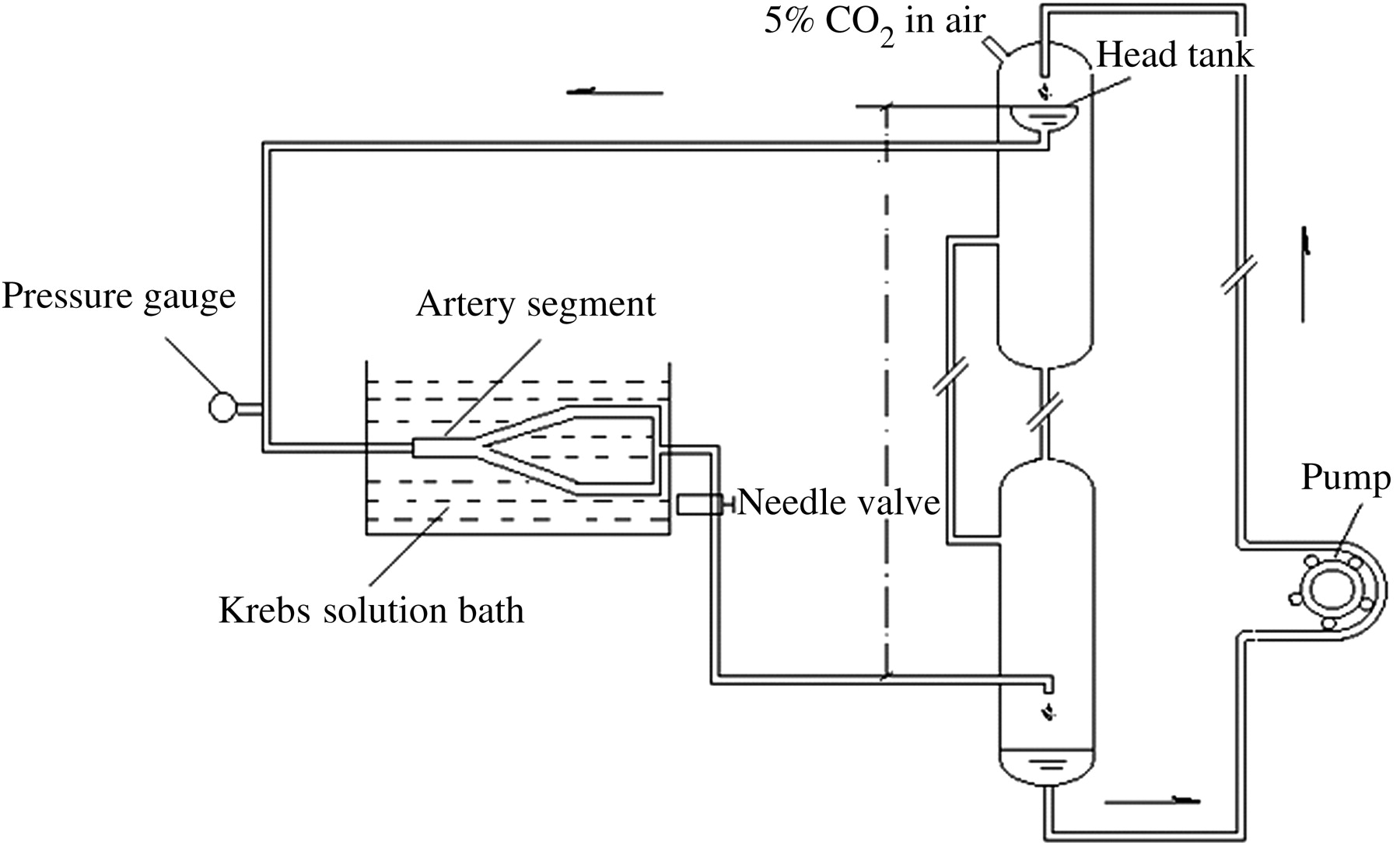
Schematic drawing of the experimental perfusion system. The overflow head tank provides a steady flow to the aorta–iliac bifurcation segment
Scanning electron microscope observation of endothelium
The arterial tissues were prepared for scanning electron microscope (SEM) observation following a modified procedure described by others. 19 Briefly, after being cannulated, the arteries were washed with 5% buffered glucose solution and stained with 0.2% silver nitrate solution. The vessels were then fixed in modified Karnovsky's fixative (2% paraformaldehyde, 1.5% glutaraldehyde, 2.5 mmol/L CaCl2, in 0.1 mmol/L cacodylate buffer, pH 7.2) at physiological pressure for 24 h. The tissue samples were incubated in 70%, 90% and 95% alcohol for five minutes each and in 100% alcohol for 10 min. The lumens of all tissue samples were opened under a binocular light microscope, coated with gold and analyzed by SEM (JSM 5800, Tokyo, Japan).
Measurement of DiI-LDL uptake along the aorta–iliac bifurcation
A standard procedure was followed throughout the entire experiment. For all of the experiments, perfusion pressure and flow rate were kept at 90 mmHg and 40 mL/min, respectively, based on the physiological parameters in the rabbit aorta. 20 The concentration of DiI-LDLs in the perfusion solution was set at 10 μg/mL. 21
The outer diameters of the test vessels were measured with a caliper, and then the vessels were sectioned to measure the wall thickness, which was later used to determine the inner diameters of the vessels. 11 The test arteries were divided into three perfusion groups as follows: (i) forward flow group (perfused in the in vivo flow direction); (ii) backward flow group (perfused in a reversed flow direction); and (iii) static group (no flow, only filled with the perfusion solution). During the experiment, the flow rate and pressure in the aorta–iliac bifurcation were monitored constantly. The experiments were all carried out for three hours at an ambient temperature of 23 ± 1°C, with the perfusion solution head tank placed in a water bath maintained at 37°C.
At the completion of the experimental perfusion, the aorta–iliac bifurcation was flushed with phosphate-buffered saline (PBS) buffer to wash off the dissociative DiI-LDL. Then, the bifurcation was fixed with a modified Karnovsky's fixative at 90 mmHg for 24 h. The bifurcation was dissected from the cannulas and given a brief rinse in PBS. The tissues around the cannulas were discarded because of possible damage to the endothelium due to the ligatures. The arteries of the bifurcation were opened axially while 15 tissue sample segments were collected from the abdomen aorta (each of 1 cm in length) and iliac vessels (each of 2 cm in length) (Figure 2). Each segment was then sectioned into anterior and posterior parts to obtain a medial and a lateral wall specimen (Figure 2).
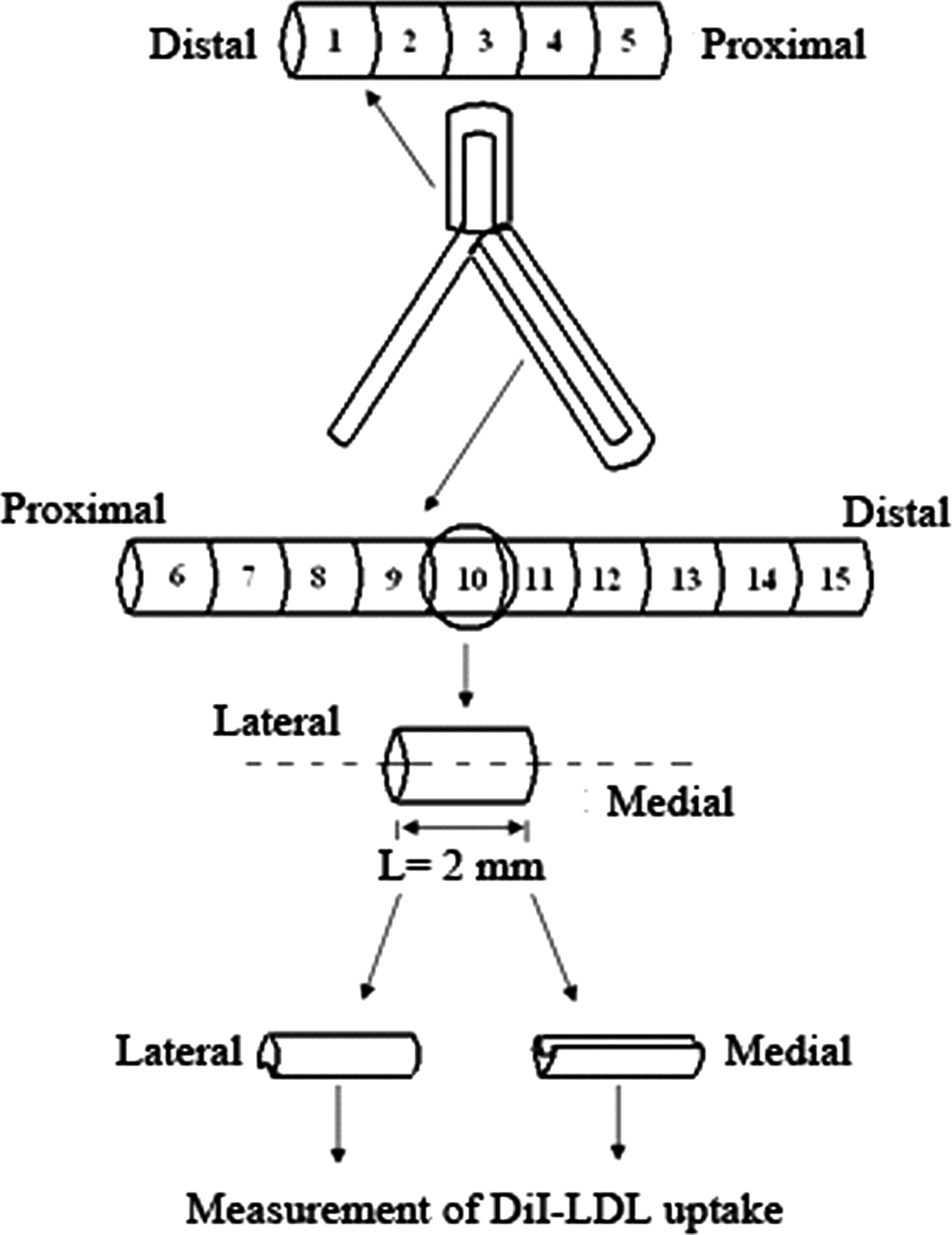
Schematic diagram of tissue sampling: 15 lateral wall and 10 medial wall specimens were obtained from each aorta–iliac bifurcation tested
Then, the tissue samples were digested in tissue solubilizer (Perkin-Elmer Life and Analytical Sciences, Boston, MA, USA) and complete lysis of the tissues was achieved by gentle pipetting of the lysate, followed by removal of cell debris by centrifugation. The fluorescence intensity of the tissue samples was measured with a spectrofluorometer (Perkin-Elmer LS-50B, Westport, CT, USA) at excitation and emission wavelengths of 514 and 550 nm, respectively. The fluorescence intensity was then converted to the concentration of lipoproteins with the calibration curve of DiI-LDLs. The amount of lipoproteins uptaken by the arterial wall was expressed as the weight of lipoproteins taken up per effective surface area per hour (ng/cm2 h).
Characterization of flow patterns
In order to correlate the distributions of LDL uptake along the aorta–iliac bifurcation with the flow patterns of different perfusion groups, the flow in the bifurcation was characterized numerically. Figure 3 shows the simplified model of the rabbit aorta–iliac bifurcation used in the numerical study. The geometry of the model was based on the measurement of the rabbit aorta–iliac bifurcation, which was symmetrical. The commercially available CFD code FLUENT 6.2 (ANSYS Inc., Canonsburg, PA, USA) was adopted for the flow study. In the present study, all numerical simulations were performed at a flow rate of 40 mL/min (Re = 220) based on the study by Becceli et al. 20 The simulations were carried out under steady flow conditions.
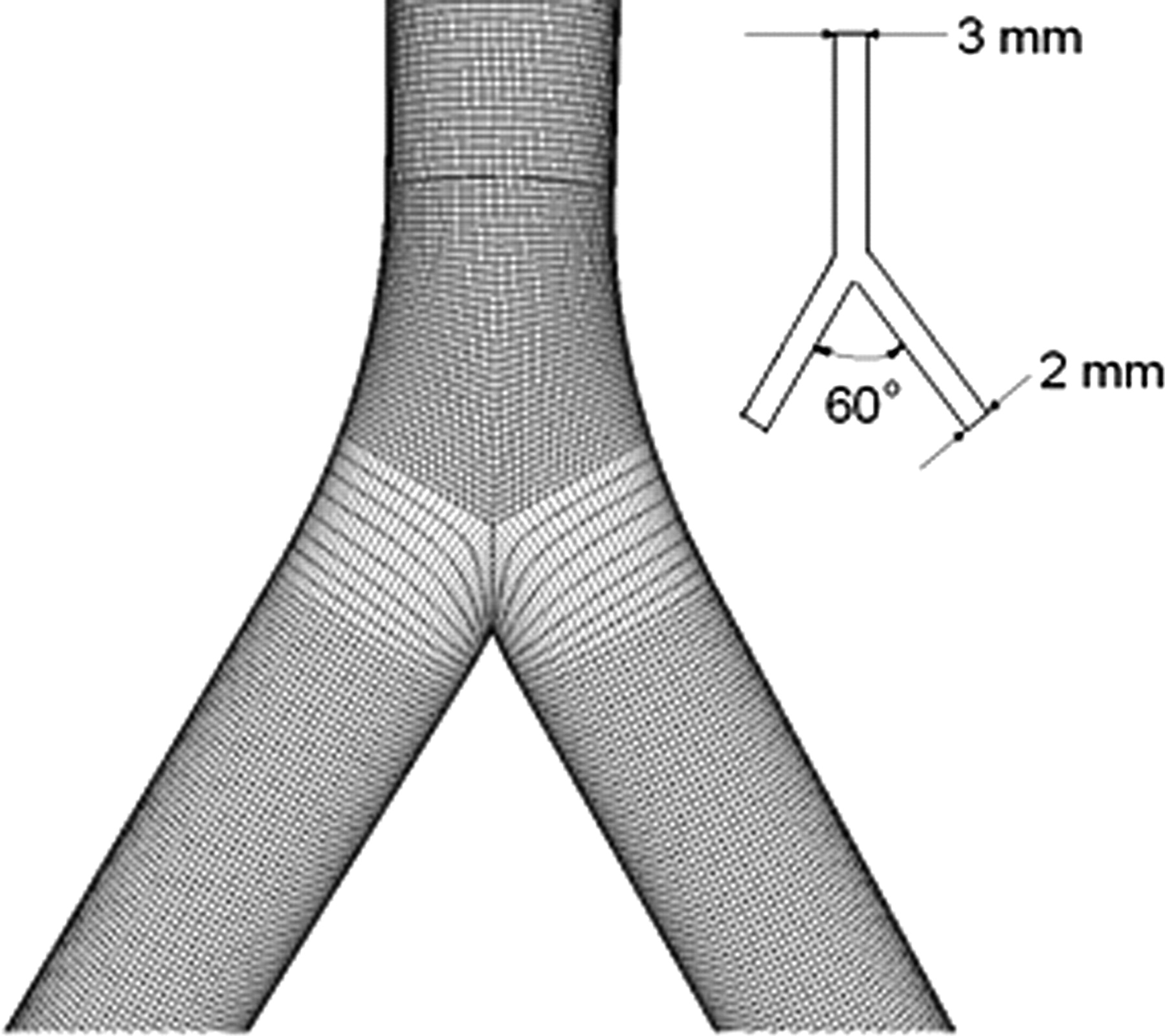
Model of the aorta–iliac bifurcation used in the computational study. The geometrical parameters of the model were based on the measurements of the rabbit aorta–iliac bifurcations
The governing equations for flow motion are given as
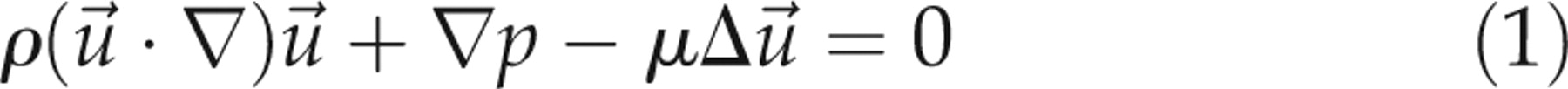


The boundary conditions for the numerical simulations are as follows:
For the forward flow:
Inlet: A uniform inflow velocity profile based on the flow rate of 40 mL/min for the axial velocity component and a zero transverse velocity component were assumed.
Outlet: At the outlet, pressure p was set at 90 mmHg. The artery wall was assumed to be rigid and non-slip.
Inlets: A uniform inflow velocity profile based on the flow rate of 20 mL/min for the axial velocity component and a zero transverse velocity component were assumed for both inlets.
Outlet: At the outlet, pressure p was set at 90 mmHg. The artery wall was assumed to be rigid and non-slip.
For the backward flow:
A finite volume method was used in the simulation. The created computational meshes of the models by the CAD software Gambit 2.2 (ANSYS Inc.) were all unstructured hexahedron grids. Discretization of the pressure and momentum at each control volume was in a second-order scheme. The iterative process of computation was terminated when the residual of continuity and velocity were all less than the convergence criterion 1.0 × 10−5.
Here, it should be mentioned that since it is impossible to keep the numerical analysis of the flow pattern in the rabbit aorta–iliac bifurcation completely identical to the experimental one, the results from the numerical flow simulation can only give qualitative indication of the flow in the experimental specimen.
Statistical analysis
Experiments were repeated at least three times under the same flow conditions. Statistical analyses were performed using the Origin 7.0 software. The results were presented as mean ± standard deviation.
Results
Flow patterns in the bifurcations
Figure 4a shows the contours of velocity on the middle plane of the aorta–iliac bifurcation for the forward flow and backward flow conditions. It can be seen from the figure that under the forward flow condition, the velocity is obviously lower along the outer walls of the bifurcation due to the vortices formed at the bifurcation (Figure 4b), while the flow was almost un-disturbed at the aorta–iliac bifurcation under the backward flow condition.

(a) Contours of velocity on the middle plane of the aorta–iliac bifurcation; (b) streamlines of flow in the bifurcation under forward flow condition, showing the formation of vortices along the outer walls of the bifurcation; and (c) wall shear stress distribution in the aorta–iliac bifurcation. Arrows indicate the flow directions. (A color version of this figure is available in the online journal)
Figure 4c shows the distribution of wall shear stress in the aorta–iliac bifurcation. As evident from the figure, wall shear stress distributes very differently for the two flow conditions. For the forward flow, wall shear stress drops suddenly from values ranging from 0.4 to 0.6 Pa in the non-disturbed areas to values less than 0.2 Pa along the outer walls of the bifurcation. At the flow divider, wall shear stress is relatively high (>1.6 Pa). The sudden drop in wall shear stress is apparently the result of flow disturbance with the formation of vortices at this location. The flow separation also leads to a directional change in wall shear stress in the zones of vortices. In contrast, for the backward flow, no low wall shear stress zones exist at the bifurcation. On the contrary, wall shear stress is enhanced just beyond the bifurcation when the two flows from the daughter branches join into the aorta.
Morphology of the endothelial cells
Figure 5 shows the morphological features of the endothelial cells in different areas of the aorta–iliac bifurcation. Scanning electron microscopy revealed that, in the upstream and downstream of the branch where blood flow was unidirectional and not disturbed with higher wall shear stress, the endothelial cells elongated with their longitudinal axis aligning to the direction of blood flow. On the other hand, at branched areas where blood flow was disturbed with low wall shear stress, the morphology of endothelial cells was round and polygonal with no preferred orientation. Careful observation revealed that the endothelial cells showed a well-defined ‘border’ between the un-branched regions and the branched regions, where the wall shear stress is relatively low. From the viewpoint of atherogenic lipid transport, at branched areas, due to the low wall shear stress, it can increase the ‘residence time’ for atherogenic agents and deposition of atherogenic particles onto the blood vessel walls, which were proposed to account for the localization of atherogenesis.

Scanning electron microscopy pictures of endothelial cells in different areas of the aorta–iliac bifurcation (original magnification ×300). Arrow indicates the direction of blood flow in vivo. (A color version of this figure is available in the online journal)
Distribution of DiI-LDL uptake along the aorta–iliac bifurcation
Figure 6 illustrates the distribution of DiI-LDL uptake along the aorta–iliac bifurcation for the three perfusion groups. As shown in Figure 6a, the distribution of DiI-LDL uptake along the aorta–iliac bifurcation in the forward flow group was obviously different from those of the other two groups.
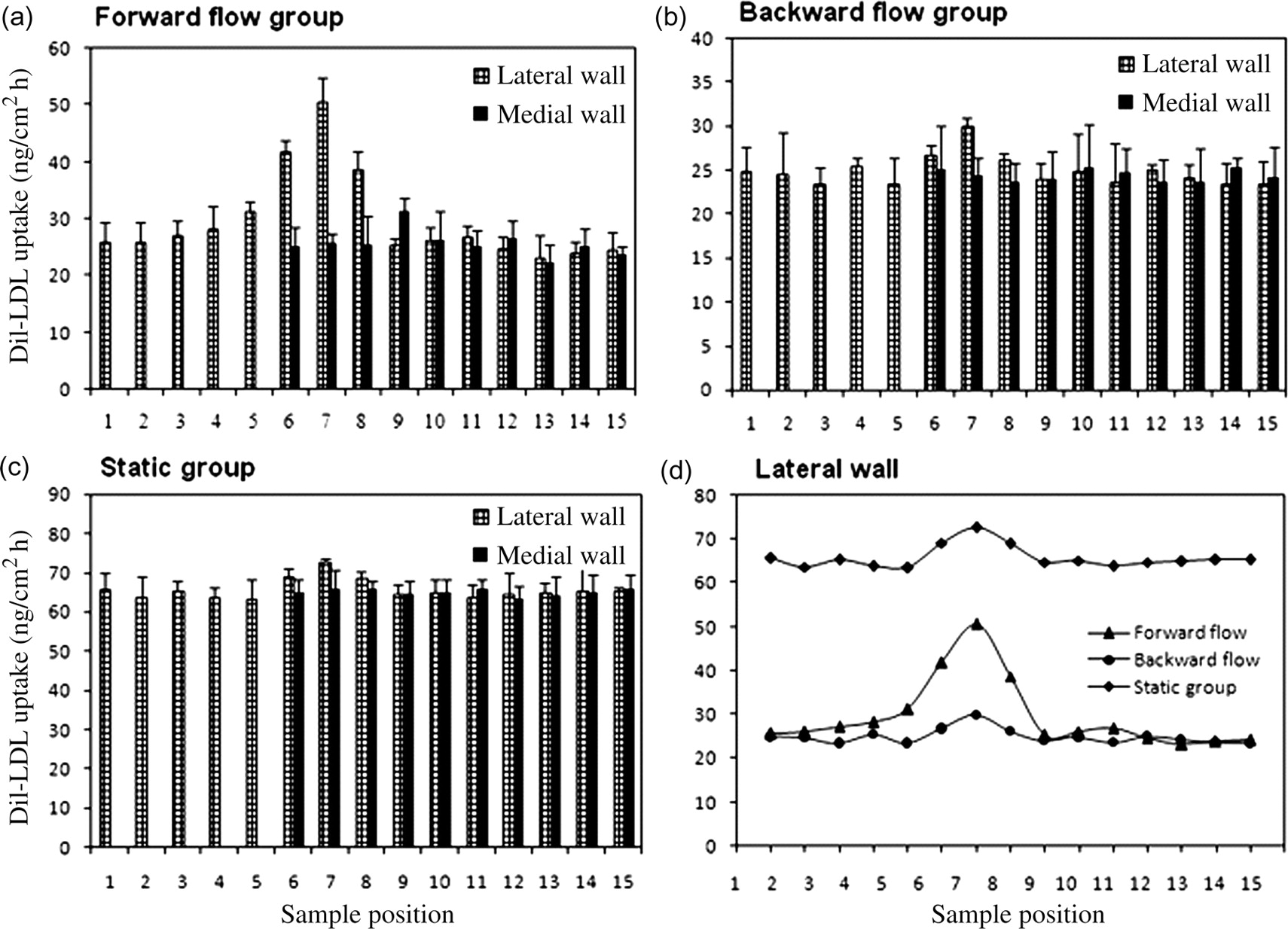
Distributions of 3,3′-dioctadecylindocarbocyanine-LDL uptake along the lateral and medial walls of the aorta–iliac bifurcation under three flow conditions. Sample position 1 corresponds to the most distal 2.0 mm specimen upstream of the bifurcation, while sample position 15 corresponds to the most distal specimen downstream of the bifurcation. The results are presented as mean ± standard deviation (n = 5)
For the forward flow group (Figure 6a), along the lateral wall of the bifurcation, DiI-LDL uptake remained almost unchanged until sample position 4, then increased drastically when the sample position approached the branching site, reaching a peak value at sample position 7. It dropped suddenly beyond sample position 7, and then gradually recovered to a similar value to that in the upstream of the branching. The trend of DiI-LDL uptake along the medial wall is very different from the lateral wall of the bifurcation. Statistically, DiI-LDL wall uptake was uniformly distributed along the medial wall.
For the backward flow group (Figure 6b), DiI-LDL uptake distributions along both the lateral and the medial walls were flat with the exception of sample positions 6, 7 and 8 of the lateral wall, which had somewhat higher uptakes than other sample positions. However, the difference was not considered statistically significant.
For the static group (Figure 6c), DiI-LDL uptake distributions along both the lateral and medial walls were more flat than that for the backward flow group. Nevertheless, its overall DiI-LDL uptakes were higher than those for the forward and backward flow groups.
Most interestingly, the experimental results showed that there was a peak in all curves of DiI-LDL uptake distribution along the lateral wall for the three perfusion groups (Figure 6d). The location of the peak corresponds to the branching areas where the morphology of endothelial cells was round and polygonal with no preferred orientation. However, the peak values of DiI-LDL uptake were very different for the three groups. It was the lowest for the backward flow group and the highest for the static group. However, the peak of the DiI-LDL uptake curve was the sharpest for the forward flow group and the flattest for the backward flow group.
Discussion
In the present study, we designed an experimental study in which the rabbit aorta–iliac bifurcation as our model was perfused with DiI-LDL solution under three different flow conditions and the patterns of DiI-LDL uptake distribution along the bifurcation were compared.
The endothelial cells of the aorta–iliac bifurcations used in the three perfusion groups had been exposed to the same in vivo flow environment with a similar wall shear stress distribution for a long time before the perfusion study. Hence, the cells had been preconditioned by the wall shear stress. Therefore, the physiological features of the endothelial cells must have a similar pattern for the three perfusion groups. If the physiological features of the endothelial cells play more important roles in the uptake of LDLs by the experimental aorta–iliac bifurcations, then the distribution of LDL uptake will not be affected by the perfusion flow direction and should follow a similar pattern for the three perfusion groups. On the other hand, according to Deng et al., 11,12 concentration polarization of LDLs in an artery is flow-dependent, i.e. the distribution of LDL luminal surface concentration depends on the flow pattern in the artery. Therefore, if the mass transport phenomenon of LDL concentration polarization plays a more important role in the LDL uptake by the experimental aorta–iliac bifurcation, then the distribution of LDL uptake will be affected by the perfusion flow direction and should have different patterns for the three perfusion groups. In this way, from the obtained experimental results, the role of endothelial cell barrier function in LDL uptake can be differentiated from the role of LDL concentration polarization.
The experimental model has two special characteristics. On the one hand, as described by Berceli et al. 20 there exist different types of blood flow in the aorto-iliac bifurcation in vivo. In the upstream section of the bifurcation (from sample positions 1–5; Figure 2), blood flow is unidirectional where shear stress is normal and distributes symmetrically. In the branching area (from sample positions 6–8), blood flow is disturbed with the formation of flow separation along the lateral wall where wall shear stress is low and the skewing of velocity profiles toward the medial wall where wall shear stress is relatively high (Figure 4). From sample positions 9–11, blood flow is recovering from its disturbed state back to normal with a gradually reducing degree of flow reversal, and wall shear stress there begins to approach its fully developed value. In the distal zone of the bifurcation downstream of the branch (from sample positions 12–15), blood flow is again characterized by somewhat fully developed, unidirectional flow with symmetric shear stress along the medial and lateral walls. Due to the in vivo blood flow characteristics of the bifurcation, the endothelial linings of the bifurcation are preconditioned by the local blood flow and therefore should possess different physiological behaviors (particularly the barrier function to macromolecules such as LDLs) according to their locations. On the other hand, when the bifurcation is perfused in vitro from different directions (i.e. forward flow or backward flow), the flow patterns in the bifurcation for the two cases are completely different. Therefore, the luminal surface concentration of LDLs must have distributed differently along the bifurcation for the two cases. For the no-flow case, according to Deng et al. 10,11 the luminal surface concentration of LDLs must be constant in the whole bifurcation. Due to these two characteristics, the experimental model is very suitable for the present study to distinguish the role of endothelial cell barrier function modulated by flow shear stress from the role of flow-dependent LDL concentration polarization in LDL uptake by the arterial wall.
Experimental results by Langile and Adamson 22 and Davies et al. 23 suggest that alterations in the morphology of endothelial cells play a fundamental role in the arterial wall permeability to LDLs. Gerrity et al. 24 examined the morphology of endothelial cells in regions exhibiting spontaneously differing permeability, and found that in areas of enhanced permeability, the endothelial cells generally had little orientation with flow. Using high cholesterol diet-fed rats as a model, Kao et al. 25 found that more LDL accumulated in the branched regions than in the un-branched regions of the artery.
The present study revealed a similar pattern of DiI-LDL uptake distribution in the rabbit aorta–iliac bifurcation model to that found by Kao et al. 25 in the branched regions. However, significant differences in the distribution of DiI-LDL uptake along the bifurcation were observed among the three flow groups. Our results showed that when the bifurcation was perfused in the in vivo forward direction, DiI-LDL uptake by the lateral wall at the branched region had a peak value that was much higher than that of the backward flow group (51 versus 30 ng/cm2 h). Furthermore, for both the backward flow group and the static group, although the peak still existed in their DiI-LDL uptake curves, the peaks of these two groups were not as sharp as that of the forward flow group. More interestingly, the overall DiI-LDL uptake of the static group was obviously higher than those of other two groups. Considering that the mass transport phenomenon of LDL concentration polarization proposed by Deng et al. 11 is flow-dependent, the results obtained in the present study therefore indicate that the permeability of the arterial endothelial linings to LDLs was not the only factor involved in the uptakes of DiI-LDLs by the bifurcation. The luminal surface concentration of DiI-LDLs determined by the flow pattern in the bifurcation also played a role, most likely a more important role, in the uptakes of the DiI-LDLs. Our reasoning for this conclusion is this: according to Deng et al. 10,11 the luminal surface concentration of LDLs is adversely correlated with wall shear stress (or wall shear rate), i.e. it increases with decreasing wall shear rate. In the case of the static group, wall shear rate in the whole bifurcation was zero, which led to the highest luminal surface concentration of DiI-LDLs in the bifurcation among the three perfusion groups. This might account for why the overall DiI-LDL uptake of the static group was the highest (Figure 6). For the forward flow group, in the disturbed flow zone at the branched area, due to the low wall shear stress there, the luminal surface concentration of DiI-LDLs was the highest when compared with other regions in the bifurcation. Therefore, the peak value of DiI-LDL uptake there was very sharp (Figure 6). We believe that the sharp peak in the curve was the combined results of higher permeability of the endothelial linings to DiI-LDLs and higher luminal surface concentration of DiI-LDLs there, and that the latter most likely contributed more in the uptake.
Nevertheless, according to Deng et al. 10,11 the luminal surface concentration of DiI-LDLs should distribute uniformly in the bifurcation for the static group and somewhat evenly in the bifurcation for the backward flow group; the peaks in the uptake curves for these two groups indicate that the endothelial linings in the aorta–iliac bifurcation might indeed possess different permeability to DiI-LDLs along the lateral wall in the branched area. This suggests that the endothelial cells in these regions had probably been modulated in vivo by the low wall shear stress with high oscillation and had loose junctions with higher permeability to DiI-LDLs.
The experiment showed that the mass transport phenomenon of LDL concentration polarization plays a more important role in atherogenesis; this result has two significances. First of all, it can explain why Leriche syndrome, also referred to as aorto-iliac occlusive disease, is prone to develop at the abdominal aorta just above the site of its bifurcation because the infiltration of LDLs there is higher according to the present experimental results. Second, the result implies that swirling flow can reduce LDL concentration polarization at the aorto-iliac bifurcation, and therefore may suppress Leriche syndrome and atherogenesis. 5
There are different opinions on the role of LDL concentration polarization in the localization of atherogenesis. Based on the findings by several researchers 26,27 that increases in shear stress lead to an increase in the permeability of the endothelium to macromolecules, which seems to contradict the theory of LDL concentration polarization, Tarbell 28 refuted the relevance of concentration polarization to LDL uptake. Nevertheless, the mass transport phenomenon of LDL concentration polarization has been verified by more and more studies. 29–32
The development of intimal hyperplasia at the distal anastomosis is responsible for a significant proportion of late failure of small diameter artery bypass surgeries. 33 It has been suggested that the accumulation of LDLs in subendothelial spaces is an important step in the development of intimal hyperplasia and atherosclerosis. 34 Therefore, optimization of bypass surgery hemodynamics to minimize concentration polarization of LDLs in bypassed arteries would be an attractive strategy to suppress intimal hyperplasia and atherogenesis. Morbiducci et al. 35 studied the blood dynamics of aortocoronary bypass with different geometries and found a linear inverse relationship between the oscillating shear index and the spiral or helical flow index, indicating a positive effect played by spiral flow in preventing the bypassed arteries from developing intimal hyperplasia. Fan et al. 36 found that S-type bypass surgery could create a swirling flow in the distal part of the bypassed artery, which apparently reduced intimal hyperplasia along the host artery floor. We guess that this reduced intimal hyperplasia might be attributed to the improved hemodynamics of the swirling flow created that could reduce concentration polarization of LDLs in the host artery.
Conclusion
The present experimental study supports the concept that both the barrier function of the arterial endothelial linings and the mass transport phenomenon of LDL concentration polarization are involved in the infiltration/accumulation of atherogenic lipids within the arterial wall. Nevertheless, our data presented evidence that the latter might play a more important role in the localization of atherogenesis.
Footnotes
ACKNOWLEDGEMENTS
We thank Qinghua Hu and Meili Liu for care of the rabbits and excellent technical assistance. This work is supported by Grants-in-Aid from the National Natural Science Research Foundation of China (Nos. 30670517, 10632010) and the Innovation Foundation of BUAA for PhD Graduates.