
Abstract
Caspase-3 is a key molecule involved in the inflammation and apoptosis of ischemia reperfusion (IR) injury. Statins are known to inhibit IR injury, but the mechanism of action remains uncertain. In the present study, the effect and underlying mechanism of ischemia alone, and reperfusion with or without atorvastatin (AT) as a timed intervention were examined, since clinically the kidney is only exposed to drug delivery during reperfusion. Male Sprague‐Dawley rats were subjected to 45‐min clamping of the left renal hilus followed by four hours reperfusion with a right nephrectomy. AT 10 mg/kg was intravenously administered after clamping the renal hilus, but prior to kidney reperfusion. Ischemia alone did cause tubulointerstitial damage (TID), protein carbonylation and caspase-3 activation with an increase in 12 kDa subunit, while reperfusion further enhanced TID, monocyte (ED-1+ cell) infiltration, apoptosis and necrosis together with caspase-3 activity and 17 kDa subunit, but reversed protein carbonylation. AT significantly reduced TID (26%), ED-1+ cell infiltration (74%), tubular apoptosis (47%) and necrosis (73%), and interstitial apoptosis (64%), as well as caspase-3 activity (26%), but did not change serum creatinine and cholesterol. Importantly, without affecting either caspase-3 active protein cleavage or S-nitrosylation, AT directly inhibited caspase-3 active enzyme in a dose-dependent manner in vitro. In conclusions, IR and AT exerted opposing effects on caspase-3 activity by differing mechanisms, with IR stimulating caspase-3 proteolytic cleavage and AT inhibiting active caspase-3 enzyme. This new inhibitory mechanism of AT may improve reperfusion tolerance in ischemic kidneys and benefit transplant recipients.
Introduction
The benefits of statins have generally been attributed to the ability of lower serum cholesterol. As inhibitors of hydroxyl-3-methylglutaryl coenzyme A reductase, statins can also produce cholesterol-independent effects. 1,2 The experimental and clinical evidence demonstrates pleiotropic effects of statins in different types of renal disease including allograft dysfunction. 3–5 Statins preserve renal function following aortic cross-clamping–declamping in man, where ischemia reperfusion (IR) injury is common. 6 Rodent studies show pretreatment with statins to reduce renal IR injury. 7,8 Clinically however, any drug treatment of the transplant recipient only begins after kidney reperfusion. The beneficial effects of statins administered prior to ischemia in experiments may be therefore overestimated when employed in transplantation, as drug treatment of the donor would be required.
Stains reduce isoprenoids synthesis, interfere with its anchoring to intracellular signaling molecules such as Ras and Rho, and directly affect the prenylation of low molecular weight proteins in intracellular signaling pathways. 1,9,10 Intracellular signaling molecules facilitate communication between growth factor receptors and cellular cytoskeletons to influence transcription factor activation, and subsequently induce cell proliferation and cytokine/chemokine activation. Statins can also directly down-regulate inflammatory chemokines and alter the downstream effects of the GTPase Ras superfamily. 11–13 In addition, statins adjust the balance between proliferation and apoptosis in different renal cell types and protect kidneys against experimental nephritis and IgA nephropathy. 12,14 However, the precise mechanisms underlying beneficial actions are not fully understood. Caspase-3 (cysteine protease) is an important enzyme involved in inflammation and apoptosis, key features of renal IR injury. 15,16
The present study was undertaken to examine the effects and mechanisms of atorvastatin (AT) on the reperfusion phase of IR injury with focuses on tubulointerstitial damage (TID), inflammation, apoptosis, necrosis and caspase-3 in rats. Three potential mechanisms for the inhibitory effect of AT on caspase-3 activity were evaluated: firstly by inhibiting caspase-3 proteolytic activation; secondly by indirect inhibition through S-nitrosylation; and thirdly by the direct inhibition of active caspase-3 enzyme.
Materials and methods
IR model
Groups (n = 4–7) of male Sprague-Dawley rats (300–350 g) were anesthetized with halothane and underwent occlusion of the left renal vascular pedicle using microvascular clips for 45 min together with right nephrectomy and followed by four hours reperfusion (IR). Animals undergoing sham-operation including laparotomy, vascular pedicle dissection and unilateral nephrectomy (C) or 45‐min ischemia only (I) were used as controls. This approach has previously been found to be the longest period of warm ischemia compatible with appropriate degree of injury and survival. 17 AT (10 mg/kg) dissolved in dimethyl sulfoxide was administered intravenously after clamping the renal artery before reperfusion (IR + AT). The dosage and administration route of AT were chosen based on previous studies, clinical doses for humans and our preliminary data. 4,18–20 Rats were housed at 20°C and 45% humidity on a 12 h light/dark cycle, and fed ad libitum on standard laboratory rat chow with free water access. All the experiments were carried out according to the regulations laid down by the Home Office (Animal Scientific Procedure Act 1986, UK).
Hematological and histological changes
At the end of reperfusion, the right kidney was removed immediately under anesthetic and blood was obtained to measure serum creatinine (SCr), triglyceride and cholesterol. Renal cortical tissues were fixed in 10% (w/v) neutral buffered formalin and paraffin-embedded for histological examination, or snap frozen and stored in liquid nitrogen for protein analysis.
Hematoxylin and eosin-stained sections were semi-quantitatively graded at ×200 magnification for TID (tubular dilation and interstitial expansion with edema, inflammatory infiltrate) based on a scale of 0–3: normal tubulointerstitium scored 0; mild TID affecting up to 25% field scored 1; moderate TID affecting 25–50% field scored 2; and severe TID exceeding 50% field scored 3. Examination was done blind by two examiners in 12 randomly selected consecutive fields and a mean value was given per kidney. 16
Evaluating inflammation by ED-1 immunostaining
Renal inflammation was evaluated by ED-1 (a specific rat monocyte/macrophage marker) immunostaining using a DAKO ChemMate EnVision™ Detection Kit (Dako, Glostrup, Denmark). Paraffin sections (4 μm) were pretreated with 0.125% (w/v) trypsin at 37°C for 10 min. An anti-ED-1 antibody (1:50 dilution, Serotec Ltd, Oxford, UK) was applied overnight at 4°C. Antibody binding was revealed using the peroxidase/diaminobenzidine. Negative control sections were incubated with normal IgG at the same protein concentration of primary antibody. Renal inflammation was evaluated by a semi-quantitative analysis of ED-1+ cells in 20 randomly selected consecutive fields at ×400 magnification. 21
Detection of apoptosis
In situ end-labeling (ISEL) fragmented DNAs was performed on 4‐μm paraffin sections using an ApopTag™ peroxidase kit (Appligene Oncor, Illkirch, France). 16 Sections were digested using 40 μg/mL of proteinase K for 15 min at 37°C, incubated with terminal deoxynucleotidyl transferase and digoxigenin-deoxyuridine triphosphate for 60 min and transferred to ‘stop’ buffer for 30 min at 37°C. After adding antidigoxigenin-peroxidase for 30 min, these sections were developed by 3′-amino-9-ethylcarbazole substrate. To substantiate the specificity of results, apoptosis was also confirmed by characteristic morphological features such as cell shrinkage and nuclear condensation. The number of apoptotic cells in tubular and interstitial areas was examined at ×400 magnification in up to 20 randomly selected consecutive cortical fields for each animal. Clusters of necrotic cells picked up by this labeling, without condensed nuclei and clear boundaries between cells, were also counted in a similar way.
Caspase-3 activity assays
Caspase-3 activity was detected by a modified Fluorometric CaspACE™ Assay System (Promega, Southampton, UK) using 100 μg renal supernatant protein. 21 This assay is based on the ability of caspase-3 to cleave the fluorogenic substrate of Ac-DEVD-amino-4-methyl coumarin (AMC). Its specificity was determined using caspase-3 inhibitor Ac-DEVD-CHO. The fluorescence of the reaction was monitored at 360 nm excitation and 460 nm emission wavelengths using an Mx 4000™ Multiplex Quantitative PCR System (Stratagene, Foster City, UK). The fluorescence intensity was calibrated with standard concentrations of AMC and caspase-3 activity was expressed as pmol AMC liberated per min per μg protein at 30°C.
Caspase-3 activity was also detected in IR kidney supernatant and human recombinant caspase-3 enzyme samples (containing 12 and 17 kDa subunits; Sigma, Dorset, UK) incubated with AT of different concentrations (5–8000 μmol/L) for 30 min at 30°C.
Measurement of caspase-3, S-nitrocysteine and β-actin protein
In all, 20–40 μg renal supernatant protein was separated on a 15% (w/v) polyacrylamide denaturing gel and electro-blotted onto Hybond-C membrane (Amersham Life Science, Buckinghamshire, UK). 21 The membrane was blocked and probed with anticaspase-3 (1:1000 dilution, Santa Cruz Biochemicals, Santa Cruz, CA, USA), anti-S-nitrosocysteine (1:500 dilution, A.G. Scientific Inc, San Diego, CA, USA) or anti-β-actin antibody (1:5000 dilution, Insight Biotechnology, Middlesex, UK). The antibody binding was revealed using a peroxidase conjugate (Dako) and ECL chemiluminescent detection system (Amersham Life Science). Developed films were semi-quantitatively analyzed using a Bio-Rad GS-800 densitometer (Bio-Rad Laboratories Ltd, Hertfordshire, UK). The optical value of volume density for caspase-3 or S-nitrosocysteine was corrected for protein loading using β-actin.
Measurement of protein carbonyls
Renal supernatants containing 35–80 μg/μL protein were used for protein carbonyl analysis by an enzyme-linked immunosorbent assay (ELISA) kit (Zenith Technology Corp Ltd, Dunedin, New Zealand). A volume of 5 μL of samples were derivatized with dinitrophenylhydrazine for 45 min at room temperature (RT) and then added into 1 mL of ELISA buffer. A volume of 200 μL of each mixture was applied to an ELISA plate in triplicate and incubated overnight at 4°C. The plate was blocked and incubated with antidinitrophenyl-biotin antibody for one hour at 37°C. After incubating with streptavidin-horseradish peroxidase for one hour at RT, the plate was developed with chromatin reagent for seven minutes and determined at 450 nm. The protein carbonyl content was calculated from a standard curve that was constructed by plotting different concentration of protein carbonyl standards (nmol/mg) against the absorbance. 22
Statistical analysis
The results are expressed as mean ± standard error of the mean. Normality tests were carried out and statistical differences were assessed by GraphPad InStat version 3 (GraphPad Software Inc, San Diego, CA, USA). Differences between two groups were compared by unpaired t-test or Mann-Whitney U test. P values <0.05 were considered to be statistically significant.
Results
Serum chemistry
Serum creatinine was significantly increased following IR compared with the sham-operated group (160 ± 18 versus 62 ± 4 μmol/L, P < 0.01), while this increase remained unaffected by AT treatment (163 ± 11 μmol/L). Neither serum triglyceride (0.50 ± 0.15 versus 0.39 ± 0.04 mmol/L) nor serum cholesterol (2.46 ± 0.14 versus 2.53 ± 0.14 mmol/L) was affected by IR + AT compared with the IR group.
Renal histology
Tubulointerstitial injury
Uninephrectomy kidneys showed a normal histology, while red blood cells accumulated in interstitial areas in ischemic kidneys (Figures 1a and b). Acute tubular injury was detected after IR with increased tubular dilation, tubular cell detachment, cells and cell debris in tubular lumens, early nuclear pyknosis and peritubular capillary dilation (Figure 1c), all of which were improved by AT treatment (Figure 1d). The score for TID was significantly increased by ischemia alone and furthered by reperfusion, but partially reduced by AT treatment (26%, Figure 1e).

Tubulointerstitial damage in hematoxylin and eosin-stained sections and ED-1+ cell infiltration in kidneys. The normal histology was shown in unineprecomy kidneys (a), while red blood cells accumulated in the interstitial area of ischemic kidneys (b). After reperfusion, the acute tubular injury was shown with increased tubular dilation, tubular cell detachment, cells and cell debris in tubular lumens, early nuclear pyknosis and peritubular capillary dilation (c), which was noticeably improved by AT treatment (d). ED-1+ cells were mainly located in tubulointerstitial areas (e, f). Histograms demonstrated the score of tubulointerstitial damage (g) and semi-quantitative data of ED-1+ cells (h) in IR kidneys with or without AT treatment. Additional AT significantly improved tubulointerstitial damage and decreased ED-1+ cells by 26% and 74%, respectively. Data represent mean ± SEM, n = 4–7. IR, ischemia reperfusion; AT, atorvastatin; SEM, standard error of the mean; C, uninephrectomy; I, 45‐min ischemia; IR, I followed by four hours reperfusion; IR + AT, IR with 10 mg/kg of AT. ^^ P < 0.01: I, IR or IR + AT compared with C; # P < 0.05, ## P < 0.01: IR or IR + AT compared with I; *P < 0.05: IR + AT compared with IR (A color version of this figure is available in the online journal)
ED-1+ cells
ED-1+ cells were located in tubular areas (Figure 1e) and interstitial areas (Figure 1f) and to a lesser extent in glomerular and vascular areas. The number of ED-1+ cells in the tubulointerstitium was significantly increased by ischemia (2-fold), markedly increased further (14-fold) following reperfusion, but significantly decreased by 74% with AT treatment (Figure 1b).
Apoptosis
Apoptotic cells were mainly located in tubulointerstitial areas (Figures 2a and b) with few in glomerular areas. Tubular apoptosis remained unaffected by ischemia alone, was greatly increased (28-fold) following reperfusion, but inhibited by AT approximately 47% (Figure 2e). Interstitial apoptosis was also significantly increased by ischemia alone (3-fold), furthered by six-fold following reperfusion, but significantly reduced by AT approximately 64% (Figure 2f).

Apoptotic and necrotic cells in kidneys. Apoptotic cells were mainly located in tubulointerstitial areas (a, b). Some necrotic cells, picked up by ISEL staining, were seen in tubular areas and tubular lumens (c, d), but no clear boundaries between cells and no condensed nuclei. Histograms demonstrated the semi-quantitative data of tubular apoptosis and necrosis (e), and interstitial apoptosis (f) in IR kidneys with or without AT treatment. Additional AT significantly reduced the number of tubular apoptotic cells, tubular necrotic cells and interstitial apoptotic cells by 47%, 73% and 64%, respectively. Data represent mean ± SEM, n = 4–7. IR, ischemia reperfusion; AT, atorvastatin; SEM, standard error of the mean; ISEL, in situ end-labeling; SEM, standard error of the mean; C: uninephrectomy; I, 45‐min ischemia; IR, I followed by four hours reperfusion; IR + AT, IR with 10 mg/kg of AT; Tapo, tubular apoptosis; Tnecro, tubular necrosis. ^^ P < 0.01: I, IR or IR + AT compared with C; ## P < 0.01: IR or IR + AT compared with I; *P < 0.05, **P < 0.01: IR + AT compared with IR (A color version of this figure is available in the online journal)
Necrosis
Some necrotic cells, picked up by ISEL staining, were noticeable in tubular areas reaching the tubular lumen (Figures 2c and d). However, there were no clear boundaries between individual cells and no condensed nuclei. The number of necrotic tubular cells remained unaffected by ischemia alone, but was greatly increased following reperfusion. Treatment with AT markedly reduced the number of tubular necrotic cells by 73% (Figure 2e).
Caspase-3
Enzyme activity
Caspase-3 activity was significantly increased by ischemia alone (69%) and further raised by 22% following reperfusion (Figure 3a). AT treatment significantly reduced caspase-3 activity by 26%, completely abolishing the increase caused by the reperfusion.
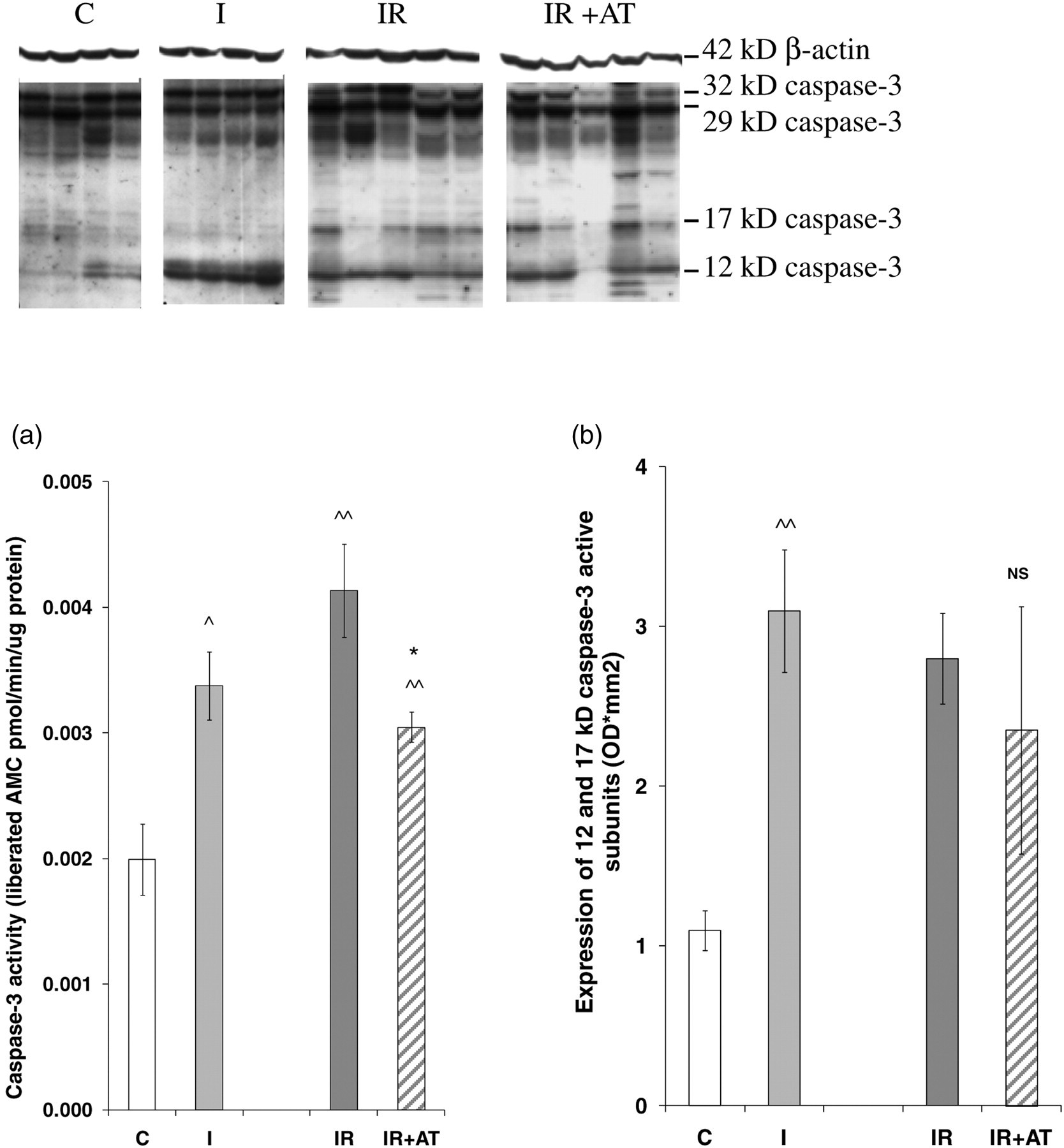
Caspase-3 activity and protein expression in kidney tissues detected by enzyme cleavage assay and Western blotting. Caspase-3 activity was significantly increased by ischemia alone (69%), furthered by 22% following reperfusion and reduced about 26% by AT treatment (a). Immunoblots showed 12, 17, 29 (intermediate) and 32 kDa (precursor) caspase-3 proteins and 42 kDa β-actin. Active subunits including 12 and 17 kDa bands were significantly increased by ischemia alone, but were not further affected by IR or IR + AT, with no significant difference between IR and IR + AT. Data represent mean ± SEM. n = 4–7. AMC, Ac-DEVD-amino-4-methyl coumarin; IR, ischemia reperfusion; AT, atorvastatin; SEM, standard error of the mean; C, uninephrectomy; I, 45-min ischemia; IR, I followed by four hours reperfusion; IR + AT, IR with 10 mg/kg of AT. ^ P < 0.05, ^^ P < 0.01: I compared with C; *P < 0.05: IR + AT compared with IR; NS: P > 0.05: IR + AT compared with IR
Protein expression
Four caspase-3 bands, 12 and 17 kDa active subunits, a 29 kDa intermediate and a 32 kDa precursor, were demonstrated by Western blotting (Figure 3). Only the 12 kDa subunit was increased by ischemia alone (3.9-fold), but reduced following reperfusion (33%). Bands of 17 and 29 kDa were increased following reperfusion (4.7- and 1.5-fold). Active subunits, 12 and 17 kDa bands merged together, were increased by ischemia (2.8-fold), but were not significantly affected by reperfusion (Figure 3b). Although mean values for all four protein bands were slightly lower with AT treatment, no any individual band showed a significant decrease compared with IR.
Protein S-nitrosylation
Protein S-nitrosylation, including a 12 kDa band, was also detected by Western blotting in kidney homogenates from all groups (Figure 4a). The expression of S-nitrocysteine was not affected by ischemia, but increased by 50% following reperfusion, and remained unchanged by AT treatment (Figure 4b).
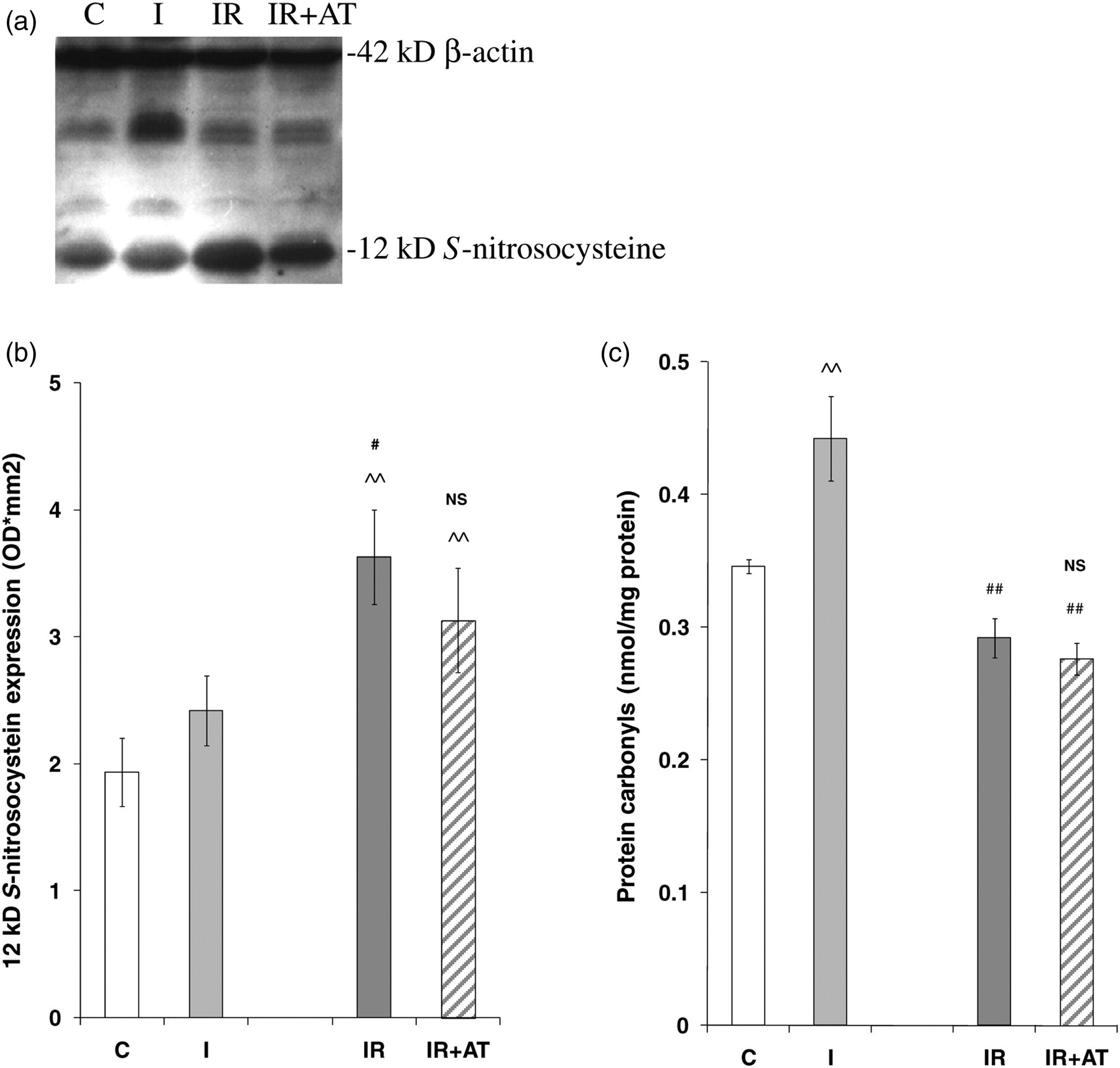
Expression of S-nitrosocysteine detected by Western blotting and level of protein carbonyles in kidney tissues. Protein S-nitrosylation was revealed in tissue homogenates from C, I, IR and IR + AT kidneys, not affected by ischemia only, but increased by 50% following reperfusion, which remained unchanged with AT treatment. The protein carbonyl was significantly increased after ischemia alone, but completely abolished by reperfusion with or without AT. Data represent mean ± SEM, n = 4–7. IR, ischemia reperfusion; AT, atorvastatin; SEM, standard error of the mean; C, uninephrectomy; I, 45-min ischemia; IR, I followed by four hours reperfusion; IR + AT, IR with 10 mg/kg of AT. ^^ P < 0.01: I, IR or IR + AT compared with C; # P < 0.05: IR or IR + AT compared with I; NS: P > 0.05: IR + AT compared with IR
Protein carbonylation
The kidney content of protein carbonyls, a marker of protein oxidation, was significantly increased after ischemia, completely abolished after reperfusion and not affected by AT treatment (Figure 4c).
In vitro effects of AT on caspase-3 activity
Renal homogenates
Caspase-3 activity was significantly inhibited in IR kidney supernatants incubated with AT starting at 1000 μmol/L (Figure 5a). Protein S-nitrosylation represented by S-nitrosocysteine expression remained unchanged by AT or a known caspase-3 inhibitor (Figure 5b), which was detected by Western blotting in the same samples of activity assay.
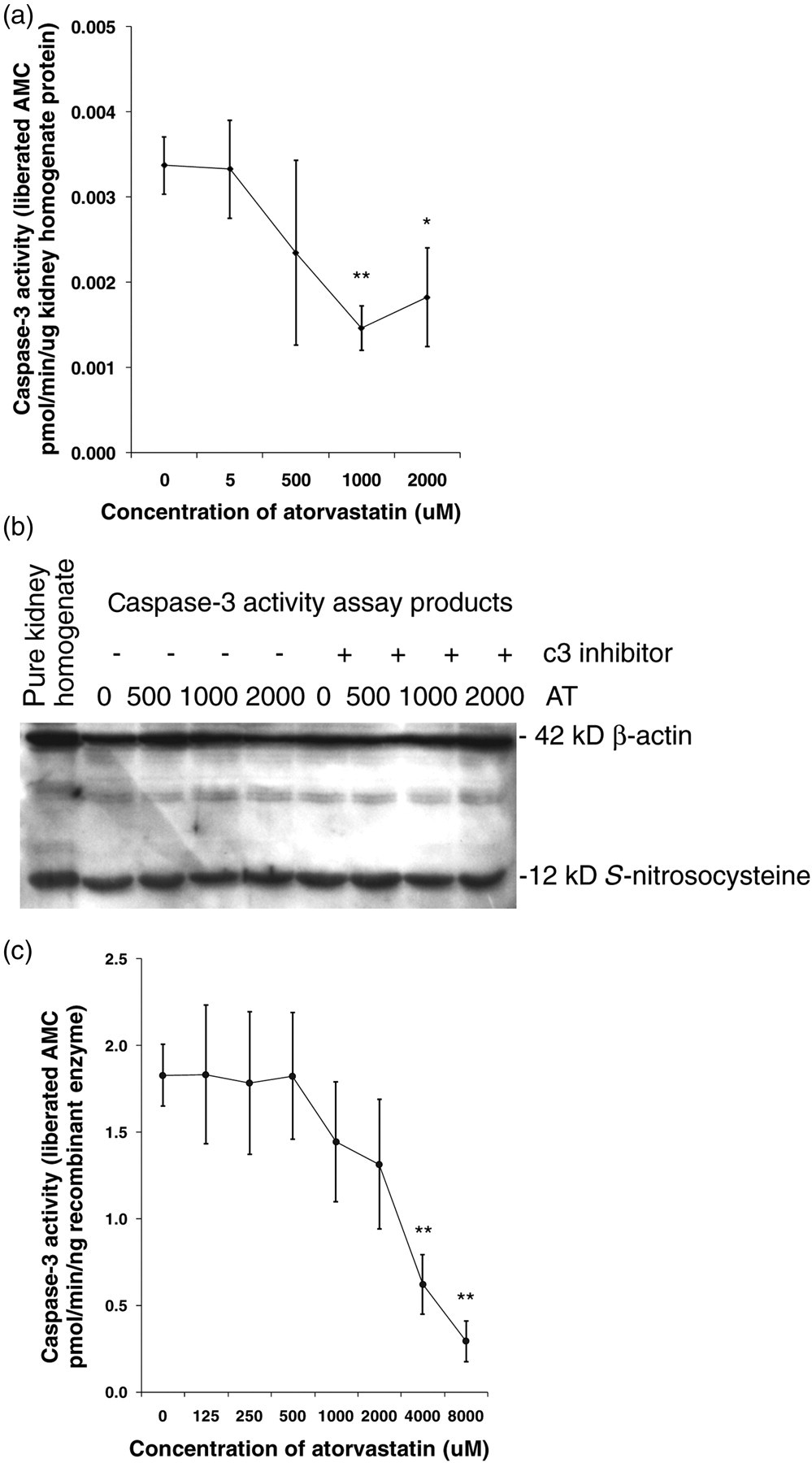
Caspase-3 activity in the IR kidney homogenate and recombinant caspase-3 enzyme incubated with AT of different concentrations. AT significantly inhibited caspase-3 activity in IR kidney homogenates after 1000 μmol/L (a). Immunoblots showed that the expression of S-nitrosocysteine was not affected by AT (0, 500, 1000 and 2000 μmol/L) in vitro or caspase-3 inhibitor (b). AT also dose-dependently inhibits caspase-3 activity in 5 ng recombinant human caspase-3, reaching a significant difference at dosages above 4000 µmol/L (c). The measurement was repeated at least three times and data represent mean ± SEM. IR, ischemia reperfusion; AT, atorvastatin; AMC, Ac‐DEVD‐amino‐4‐methyl coumarin *P < 0.05, **P < 0.01
Recombinant protein
When added as a recombinant protein, caspase-3 activity was inhibited in a dose-dependent manner by AT (1000–8000 μmol/L) reaching a significantly statistical difference at concentrations above 4000 μmol/L (Figure 5c).
Discussion
AT inhibited the increase in ED-1+ cell infiltration > tubular necrosis > interstitial apoptosis > tubular apoptosis > TID in the rat kidney following IR. Caspase-3 activity, enhanced following IR, was reduced by AT without affecting the cleavage of caspase-3 active subunits. Ischemia alone induced TID, protein carbonylation and up-regulated caspase-3 activity and its 12 kDa subunit, while the reperfusion further increased TID together with ED-1+ cell infiltration, apoptosis and necrosis, as well as caspase-3 activity and 17 kDa subunit, but reversed protein carbonylation. AT and IR exerted opposing effects on caspase-3 activity by differing mechanisms, with IR stimulating caspase-3 proteolytic cleavage and AT direct inhibiting active enzyme.
An important feature of the experimental protocol of this study was the time of AT administration, i.e. after the application of the ischemic clamp but before reperfusion. The pattern of AT responses was consistent with the effects primarily caused by reperfusion. This approach suggests that perfusing the donor kidney or pretreating the recipient with statins could provide a potentially important therapeutic window to limit IR injury following cadaveric donation. Since a live donor is increasingly important in kidney transplantation, further treatment of the donor with statins as well as the recipient to reduce IR injury may also be worthy of consideration. However, AT has no effect on serum creatinine, which might be due to a very brief reperfusion of four hours. Although it was believed that AT might have an effect on renal function after longer reperfusion, the evidence will be needed from a new set of long-term experiments.
Statins have previously been suggested to suppress inflammatory responses through inhibiting specific monocyte surface receptors, adhesion molecules and inhibiting integrin-dependent leukocyte adhesion or cytokines/chemokines. 23,24 In this study, AT dramatically reduced ED+ cell infiltration after reperfusion. Its mechanisms may at least include direct inhibiting caspase-3 activity to reduce the maturation of proinflammatory mediators. 15,25,26
It has been demonstrated that AT induced apoptosis in vascular smooth muscle cells. 27 However, in marked contrast, AT significantly reduced renal tubular apoptosis by decreasing transforming growth factor-β in the obstructed kidney. 28 In this study, tubular apoptosis was augmented only by the reperfusion, whereas interstitial apoptosis was increased by both ischemia and reperfusion. AT effectively reduced the number of apoptotic cells in tubular and interstitial areas. Both 12 and 17 kDa caspase-3 subunits may play coordinating roles in the execution of apoptosis following IR, as the increase of 12 kDa subunit during ischemia primed renal cells for apoptosis after reperfusion when the 17 kDa subunit was also elevated.
AT significantly reduced the increase in not only apoptosis, but also necrosis in IR kidneys, and consequently improved TID. In a previous study of rat IR injury, greater benefit was demonstrated following inhibition of caspase-1 than caspase-3, mediated primarily via necrosis in contrast to apoptosis. 29 Down-regulation of caspase-3 activity may therefore be just one possible mechanism for AT to enhance reperfusion tolerance. AT given to the isolated mouse heart at reperfusion attenuates lethal reperfusion-induced injury by inducing the phosphorylation of multiple prosurvival pathways involving PI3K/AKT, p44/42 MAPK, and p38 MAPK and HSP27. 2 AT‐improved TID following renal IR injury may also occur through similar pathways.
The caspase-3 precursor requires two-step proteolytic processing for activation. One is between 12 and 17 kDa proteins mediated by initiator caspases, which is followed by a caspase-3-dependent prodomain cleavage. 30,31 The 12 kDa caspase-3 was increased by ischemia alone, whereas the 17 kDa subunit was increased after reperfusion together with 29 kDa intermediate. Theoretically, both 12 and 17 kDa caspase-3 subunits contribute to enzyme activity. Here, the increase of caspase-3 activity at the ischemic stage might be attributed to the 12 kDa subunit, while the increase following reperfusion was due to both 12 and 17 kDa subunits. Interestingly, AT abolished the increase in caspase-3 activity following reperfusion to below ischemic levels, but without preventing the cleavage of its active subunits.
To explore the mechanism of AT action, protein S-nitrosylation was investigated. Pretreatment with AT enhances nitric oxide (NO) availability in aging rats and improves renal dynamics and histology following IR. 32 NO can cause S-nitrosylation of active-site cysteine residues of cysteine-containing enzymes such as caspase-3, which subsequently inhibits enzyme activity. 33,34 In this study, a 12 kDa S-nitrosocysteine was increased following reperfusion, which is total protein S-nitrosylation, but may include S-nitrosylation of 12 kDa caspase-3. However, the increase in S-nitrosocysteine was associated with an increase not a decrease in caspase-3 activity. In addition, S-nitrosocysteine was not changed by AT in vivo and in vitro, although caspase-3 activity was reduced. Therefore, protein S-nitrosylation is not associated with caspase-3 activity or AT treatment. A further study was then undertaken using recombinant caspase-3 enzyme incubated with AT, which showed good inhibition of activity albeit with an IC50 of approximately 2500 μmol/L. Therefore, AT can directly inhibit active caspase-3 enzyme, although its concentration is relatively high and non-specific effects cannot be absolutely excluded. In contrast, the caspase-3 inhibitor, DEVD-FMK, allows the first cleavage of caspase-3, but blocks the active site preventing the autocatalytic processing of its prodomain. 30
Finally, oxidized proteins also contribute to renal injury. Oxidation of extracellular matrix proteins may enhance mesangial cell apoptosis and lead to hypocellularity in glomerulosclerosis, with enhanced caspase activation. 35 Chronic accumulation of advanced oxidation proteins may promote inflammation in diabetic kidneys with macrophage infiltration and lead to deteriorated renal structure and function. 36 The increase of protein carbonyls was showed in rat kidneys with 30‐min ischemia and two hours reperfusion accompanied by aggravated tubular necrosis. 37 In this model, protein carbonyls were increased after 45‐min ischemia, but reversed following four hours reperfusion, which appeared to be irrelevant to AT treatment.
In conclusion, both ischemia and reperfusion contribute to an increase in caspase-3 activity that attributes to 12 and 17 kDa active subunits coordinately. Ischemia alone induces protein carbonylation, but few cellular infiltration, apoptosis and necrosis, while reperfusion has reverse effects on these parameters, together with further TID. AT, administered after ischemia but before reperfusion, improves reperfusion tolerance with reduced TID through a direct inhibition of the active component of caspase-3 enzyme, independent of cholesterol. The timing of AT intervention and a new mechanism of AT inhibitory effect on IR injury provide further evidence for its clinical application in kidney transplantation.
Footnotes
ACKNOWLEDGEMENTS
This study was supported by a research grant from The Pfizer and The Research & Development Directorate of the University Hospitals of Leicester NHS Trust. An abstract from part of this manuscript has been published in Transplant Int 2009; 22 s2: 43.