
Abstract
Serum paraoxonase 1 (PON1) has been reported to be an important contributor to the antioxidant and anti-inflammatory activities of HDL, avoiding LDL oxidation. The activity of this enzyme is reduced in patients with renal insufficiency, caused by elevated oxidative stress and disturbances of apolipoprotein metabolism. Therapeutic utilization of antioxidants to control renal oxidative stress may be an effective therapy in renal protection. The aim was to investigate the protective effects of several antioxidant compounds against the oxidative stress associated to renal failure induced by ethylene glycol (EG), focusing on the possible role of serum PON1 activity. Fifty-four male Wistar rats were randomly assigned to six groups (n = 9): an untreated control (C) group, an EG-treated group, a catechin (CAT)-treated group, an epicatechin (EPI)-treated group, a quercetin (QUE)-treated group and a folk herbal extract (FHE)-treated group. After 16 d of treatment, calcium oxalate lithiasis was induced in the rats using EG. After eight days (treatment + EG), the animals were sacrificed. EG treatment impaired kidney composition, increased oxidative damage, and decreased serum paraoxonase and arylesterase activities. CAT, QUE and the FHE Fagolitos improved oxidative status by enhancing antioxidant defenses – superoxide dismutase and PON1 activities – and reducing oxidative damage, thus reinforcing the idea of a possible role of PON1 in the protective effects of QUE against the deleterious consequences of oxidative stress in kidney.
Introduction
Ethylene glycol (EG) administration to rats constitutes a useful model for the study of renal papillary stone development linked to oxidative cell damage due to hyperoxaluria. 1 The oxalic acid that results from the detoxification of EG precipitates as calcium oxalate monohydrate in numerous tissues, especially the kidney. 2 Oxalate exposure alters mitochondrial function, induces changes in gene expression, activates apoptosis and promotes free radical production and lipid peroxidation, which can result in renal failure. 2,3 The oxidative damage appears primarily due to hyperoxaluria and is augmented by crystal deposition in the renal tubules. 4
Chronic renal failure (CRF) is the end-stage of most chronic renal diseases and is associated with a tendency to atherosclerosis due to enhanced oxidative stress and insufficient antioxidant enzyme activities together with abnormalities in lipid parameters. 5 The incidence of cardiovascular complications in CRF is on average about 20 times higher than the normal population. 6
Serum paraoxonase 1 (PON1) is a calcium-dependent esterase synthesized in the liver and closely associated to HDL containing both apolipoprotein A1 and apolipoprotein J (apoA1 and apoJ). 7 PON1 has been reported to be an important contributor to the antioxidant and anti-inflammatory activities of HDL, avoiding LDL oxidation. Thus, PON1 protects the endothelium from the pro-oxidant effect of oxidized LDL and from atherosclerosis development. 8 In addition, PON1 hydrolyses homocysteine thiolactone and prevents homocysteinemia, which is also involved in atherogenesis. 9 The activity of this enzyme is reduced in patients with enhanced atherosclerosis, in disorders of lipoprotein metabolism, in renal insufficiency and after renal transplantation, caused by elevated oxidative stress and disturbances of apolipoprotein metabolism. 10 Given that increased oxidative stress has been observed in patients with cardiovascular diseases, the use of nutritional antioxidants for the attenuation of atherosclerosis development has been suggested. 11 In fact, supplementation with antioxidant agents is reported to rescue renal epithelial cells from oxalate-induced generation of reactive oxygen species (ROS), lipid peroxidation and other toxic effects. 12 Since ROS appears to be responsible for cellular injury, therapeutic utilization of antioxidants to control renal oxidative stress may prove an effective therapy in renal protection. 13
Plants remain the basis for a large proportion of the medications used today for the treatment of a variety of diseases, 14 due to their content in polyphenolic compounds, such as quercetin (QUE) and catechin (CAT), which have a high antioxidant activity. In fact, a traditional herbal preparation, very popular in Spain during the 1970s (known as Fagolitos), has been shown to have anti-lithiasic action probably due to its antioxidant properties. 1
This study was performed to investigate the potentially protective effects of the antioxidant flavonoids CAT, epicatechin (EPI) and QUE and of the traditional herbal preparation named Fagolitos against the oxidative stress associated with renal failure induced by EG. Since EG induces oxidative-stress-related renal failure and subsequent atherogenesis, we focused on the possible involvement of serum PON1 activity in the antioxidant response and we analyzed the main factors involved in its stability and function.
Materials and methods
Materials
CAT, EPI, QUE and substrates for the measurement of PON1 activities (diethyl p-nitrophenyl phosphate and phenylacetate) were supplied by Sigma-Aldrich (St Louis, MO, USA). Folk herbal extract (FHE) was prepared and supplied by Salva Trobat Chemist's (Palma de Mallorca, Spain). An OxyBlot™ Protein Oxidation Detection kit was obtained from Invitrogen (Carlsbad, CA, USA). The kit for measurement of serum HDL-cholesterol (Ref. 1133505) was purchased from Linear Chemicals SL (Barcelona, Spain). Rabbit polyclonal antibody to human PON1 was kindly provided by MI Mackness and B Mackness (University of Manchester, Manchester, UK). 15 Rabbit polyclonal antibody to human apoA1 (Cat. No. 178422) was purchased from Calbiochem (Madrid, Spain) and goat polyclonal antibody to rat apoJ (Cat. No. sc-137147) was purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). The enhanced chemiluminescence Western blotting analysis reagents were supplied by Bio-Rad (Hercules, CA, USA). Routine chemicals were purchased from Sigma-Aldrich and Panreac (Barcelona, Spain).
Animals and treatments
Animal experiments were performed in accordance with general guidelines approved by our institutional ethics committee and EU regulations (86/609/CEE and 2003/65/CE). Fifty-four male Wistar rats of 15 weeks of age (Harlan Laboratories Models SL, Barcelona, Spain), each weighing approximately 260 g, were acclimated for seven days in cages prior to experimental treatment. Rats had ad libitum access to pelleted standard chow (A04 Panlab, Barcelona, Spain) and to tap water under a controlled 12-h light/dark cycle at 22 ± 2°C.
The animals were divided into six groups (nine animals per group): an untreated control (C) group, an EG-treated group, a CAT-treated group, an EPI-treated group, a QUE-treated group and a FHE-treated group. For 16 d, the control group and the EG group were supplied with drinking water without additives, the CAT, EPI and QUE groups were supplied with drinking water supplemented with 100 mg/L of CAT, EPI or QUE, and the FHE-treated group was supplied with drinking water supplemented with 7 mL/L of the FHE. The dose of FHE administered to the rats was equivalent to the human dose and was calculated considering that a dose of 30 mL/d of herbal preparation was recommended for humans. FHE contains fluid extracts of Arctostaphylos uva-ursi L. (2.16%), Zea mays L. (2.16%) and Ricinus zanzibariensis L. (46.48%), tincture of Sabal serrulata L. (21.5%), mother tincture of Agathosma betulina L. (17.5%), glycerin (10%) and anis essence (0.2%). After 16 d, the drinking water that contained the corresponding additives was also supplemented for a further eight days with 0.8% v/v EG plus 1% w/v NH4Cl. Antioxidant intake doses were 5.5 mg/kg body weight per day for CAT and QUE, 5.0 mg/kg body weight per day for EPI and 0.4 mL/kg body weight per day for the Fagolitos mixture.
Sample preparation
Rats were sacrificed by decapitation. Blood was collected and serum was obtained and stored at −80°C until analyzed. Kidneys were rapidly removed and weighed. Kidneys were minced and homogenized in an ice bath in 50 mmol/L potassium phosphate buffer (pH 7.4). The homogenate was divided into two aliquots. The first aliquot was centrifuged at 700
Serum PON1 activities and HDL-cholesterol levels
Serum PON1 activities were assayed in a microtiter plate spectrophotometer (Bio-Tek© Instruments, Winooski, VT, USA) by measuring the rate of hydrolysis of phenylacetate (arylesterase activity, EC3.1.8.1) and paraoxon (paraoxonase activity, EC3.1.1.2), as previously described. 16 HDL-cholesterol levels were measured using an enzymatic homogeneous assay kit.
Western blot analysis of serum PON1, apoA1 and apoJ protein levels
Thirty micrograms of serum proteins were fractionated on 15% (PON1) or 12.5% (apoA1 and apoJ) sodium dodecyl sulfate polyacrylamide gel electrophoresis gels and electrotransferred onto a nitrocellulose filter. Rabbit polyclonal antibodies against human apoA1 and rat PON1 and goat polyclonal antibody against apoJ were used as primary antibodies. Development of immunoblots was performed using an enhanced chemiluminescence kit. Bands were quantified by ChemiDoc XRS (Bio-Rad). Autoradiograms revealed an apparent molecular mass of 43, 50 and 28 kDa for PON1, apoJ and apoA1, respectively.
Kidney composition, protein carbonyl groups and enzyme activities
Protein and total DNA levels were determined in the homogenate supernatants by using the Bradford 17 and the diaminobenzoic acid 18 methods, respectively. Total lipid content was measured using the Folch method. 19 Protein carbonyl groups, as an index of protein oxidation, were measured in kidney homogenates by immunoblot detection using the OxyBlot™ Protein Oxidation Detection Kit according to the manufacturer's protocol with several modifications. 20 Citrate synthase (CS) 21 and SOD 22 activities were assayed by spectrophotometric methods.
Statistical analysis
All data are expressed as the mean values ± SEM of nine animals per group. Statistical differences between experimental groups were analyzed by one-way analysis of variance. Student's t-test, as post hoc comparison, was performed when a treatment effect was shown. A P value of less than 0.05 was considered statistically significant. All statistical analyses were performed using a statistical software package (SPSS 17.0 for Windows, SPSS Inc, Chicago, IL, USA).
Results
Kidney composition, protein oxidative damage and SOD and CS activities
In comparison to the C group, treatment with EG induced a decrease in kidney DNA content that did not reach statistical significance (P = 0.13 for treatment effect) (Table 1). EG treatment induced an increase in kidney lipid content that was prevented by all the antioxidant compounds assayed. Carbonyl groups increased with EG treatment, but this increase was prevented by all the antioxidants tested. SOD and CS activities were not modified by EG treatment but SOD activity was increased by CAT, and CS activity was increased by QUE, CAT and FHE. No treatment effect in protein levels was found.
Kidney composition, protein oxidative damage, and SOD and CS activities
C, control; EG, ethylene glycol; QUE, quercetin; CAT, catechin; EPI, epicatechin; FHE, folk herbal extract; SOD, Mn-superoxide dismutase; CS, citrate synthase
Values are means ± SE of nine animals per group. One-way ANOVA post hoc comparisons (P < 0.005): aindicates EG, QUE, CAT, EPI and FHE versus C, bindicates QUE, CAT, EPI and FHE versus EG, cindicates CAT, EPI and FHE versus QUE, dindicates EPI and FHE versus CAT and eindicates FHE versus EPI
Serum HDL-cholesterol levels and paraoxonase activities
Serum PON1 activity (Table 2) decreased with EG treatment and values were completely restored by QUE treatment and partially restored by FHE, resulting in statistically significant higher activity values in the QUE group compared with the rest of the antioxidants tested. EG treatment decreased serum AE activity. QUE and, to a higher degree, FHE increased this activity. In fact, FHE was the only compound that completely restored control AE activity values. HDL cholesterol levels did not show significant differences between groups.
Serum HDL-cholesterol levels and paraoxonase activities
C, control; EG, ethylene glycol; QUE, quercetin; CAT, catechin; EPI, epicatechin; FHE, folk herbal extract; HDLc, high density lipoprotein-cholesterol; PON1, paraoxonase 1; AE, arylesterase
Values are means ± SE of nine animals per group. One-way ANOVA post hoc comparisons (P < 0.005): aindicates EG, QUE, CAT, EPI and FHE versus C, bindicates QUE, CAT, EPI and FHE versus EG, cindicates CAT, EPI and FHE versus QUE, dindicates EPI and FHE versus CAT and eindicates FHE versus EPI
Serum PON1, ApoA1 and ApoJ protein levels
Although EG administration did not modify PON1 protein levels, they were significantly increased by FHE treatment (Table 3). In contrast, FHE treatment decreased ApoJ protein levels although differences did not reach statistical significance. As a whole, no significant treatment effect in ApoA1 or ApoJ protein levels was found.
Serum PON1, ApoA1 and ApoJ protein levels
C, control; EG, ethylene glycol; QUE, quercetin; CAT, catechin; EPI, epicatechin; FHE, folk herbal extract; PON1, paraoxonase 1; ApoA1, apolipoprotein A1; ApoJ, apolipoprotein J
Values are means ± SE of nine animals per group. One-way ANOVA post hoc comparisons (P < 0.005): aindicates EG, QUE, CAT, EPI and FHE versus C, bindicates QUE, CAT, EPI and FHE versus EG, cindicates CAT, EPI and FHE versus QUE, dindicates EPI and FHE versus CAT and eindicates FHE versus EPI
PON1 activity to ApoA1 and ApoJ protein level ratios
EG treatment induced a decrease of PON1 to ApoA1 ratio that was completely prevented by QUE treatment and partially by FHE (Figure 1a). The same profile was observed for PON1 to ApoJ ratio (Figure 1b).
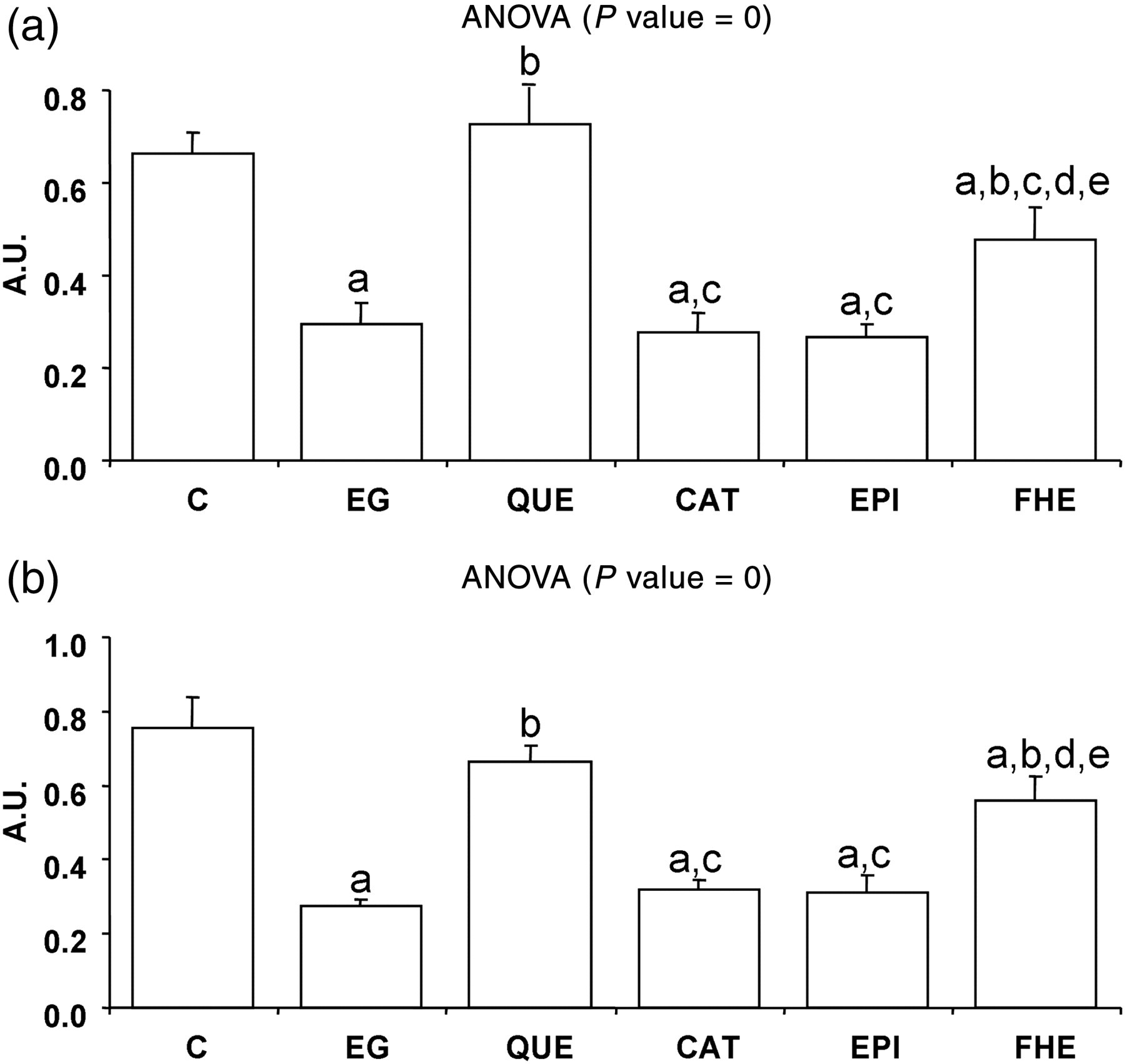
PON1 activity to ApoA1 ratio (a) and PON1 activity to ApoJ ratio (b). C, control; EG, ethylene glycol; QUE, quercetin; CAT, catechin; EPI, epicatechin; FHE, folk herbal extract; PON1, paraoxonase 1; ApoA1, apolipoprotein A1; ApoJ, apolipoprotein J. Values are means ± SE of nine animals per group. One-way post hoc comparisons (P < 0.005): aindicates EG, QUE, CAT, EPI and FHE versus C; bindicates QUE, CAT, EPI and FHE versus EG; cindicates CAT, EPI and FHE versus QUE; dindicates EPI and FHE versus CAT and eindicates FHE versus EPI
Discussion
Oral EG administration induces chronic hyperoxaluria and generates calcium oxalate deposition in kidneys. 4 Exposure to a high concentration of oxalate results in the production of free radicals, inducing oxidative stress and subsequent renal damage. 23 The results obtained in this study point to important changes in tissue composition induced by EG treatment. The kidney lipid content is increased but the effect of EG is counteracted by all the antioxidants tested, especially by QUE and FHE. The lipid accumulation in kidney associated with EG could be due to the mitochondrial dysfunction associated with hyperoxaluria, 24 which impairs fatty acid oxidation and leads to fat accumulation in cells. In this sense, the effect of the antioxidants would occur through the improvement of mitochondrial function. Accordingly, an enhancement of lipid catabolism by CAT in exercising muscle of high-fat diet-fed mice has been previously described, 25,26 and it is in accordance with the increased levels of CS activity observed in the CAT group.
The increased oxidative damage shown by the kidney of EG-treated animals could be the result of both the increased ROS production associated with oxalate accumulation 23 and the lack of antioxidant response, as suggested by the maintenance of SOD activity. In fact, variations in SOD activity corresponding to the EG dose and the duration of the treatment have been previously reported. 27,28 Protein oxidative damage was prevented by all the antioxidants assayed, especially CAT, the effects of which were also extended to an increase in SOD activity. In fact, the inhibitory effect of green tea (which contains approximately 13% CAT) on calcium oxalate urolithiasis is most likely due to its antioxidant effects, which include increased activity of SOD. 29
Paraoxonase activity has been suggested to play a role as an antioxidant defense. 30 In our study, the decreased serum PON1 activity induced by EG treatment could be related to the oxidative stress generated by oxalate accumulation. 2 Thus, PON1 protection against oxidation has been reported to result in a partial, time-dependent inactivation of the enzyme. 31 It is well known that PON1 loses its activity in an oxidative environment. Therefore, any factors that affect the status of oxidative stress will also affect PON1 activity. 32 In fact, in a pro-oxidant state associated with body weight gain induced by a high-fat diet, we have found a decrease in PON1 activity. 30 The EG-associated decrease in PON1 activity could be mainly attributed to alterations in the PON1 structure affecting its function, since no changes in the levels of PON1 protein or in the enzyme stability have been observed. ApoA1, the main protein component of HDL, plays a key role in the stability and function of PON1, 33 being considered a PON1 stability marker, and their levels were unchanged in all the experimental groups. Since a decreased activity of PON1 has been found in hemodialysis patients, 34 a role for PON1 in the defence against the oxidative stress associated with renal failure cannot be ruled out. In this sense, a relationship between decreased PON1 activity and the higher incidence of cardiovascular disease in CRF patients has been suggested. 35
All the antioxidants tested in this study induced an increase in serum PON1 activity, probably reflecting the prevention of the possible inactivation of PON1 induced by EG treatment. This protective action is more marked when rats are treated with QUE, a ubiquitous flavonoid present in fruits and vegetables. QUE was the only compound among those analyzed in this study to be able to revert the PON1 levels in the EG-treated rats to control values. Our results are in accordance with the previously reported capacity of QUE to increase serum paraoxonase activity, compatible with an antiatherogenic role for this dietary antioxidant. 32 Although the effects of the compounds tested on serum PON1 activity could be a consequence of their antioxidant properties, a direct effect on PON1 gene expression cannot be ruled out, as has been reported for QUE. 32 In fact, in the FHE-supplemented rats, the increase in serum PON1 activity is accompanied by an increase in serum PON1 levels suggesting the increased expression of PON1 in this group. It is worth noting that at least two of the plant extracts included in the FHE mixture contain QUE, to which part of the observed effects of this herbal preparation could be attributed. Moreover, the higher PON1 to ApoJ ratio shown by QUE- and FHE-supplemented rats, compared with EG rats, suggest that this HDL subpopulation to which PON1 associates 30 is more enriched in the enzyme and further reinforces a role for QUE enhancing PON1 antioxidant capacity.
In summary, EG treatment impairs kidney composition by increasing lipid accumulation, which could be attributed to an alteration in mitochondrial oxidative capacity. CAT, QUE and the FHE Fagolitos are the antioxidants with a more marked protective effect on EG-induced renal damage. The oxidative damage protection exerted by these antioxidants involves an induction of SOD activity, in the case of CAT, and of PON1 activity, in the case of QUE, which is also present in Fagolitos mixture. Thus, PON1 activity emerges as a contributor of the protective effects of QUE against the deleterious consequences of oxidative stress in kidney.
Footnotes
ACKNOWLEDGEMENTS
We thank J Sastre-Serra and C Bergas Niell for helpful animal sacrifice and technical assistance, respectively, and Dr MI Mackness and Dr B Mackness from the University of Manchester (Department of Medicine) for kindly providing the rabbit anti-human PON1 antibody. This work was supported by the Ministerio de Ciencia y Tecnología (CTQ2006-05640) and the Fondo de Investigaciones Sanitarias (PI060293) of the Spanish Government, and by the Conselleria d'Innovació i Energia of the Comunitat Autónoma de les Illes Balears (PROGECIB-1C and PCTIB-2005GC4-06). EA-C, AN-C and YG-P were funded by grants from the Comunitat Autònoma de les Illes Balears. We also express our gratitude to Salva Trobat Chemist's (Palma de Mallorca, Spain) for preparing and supplying the herbal extract.