
Abstract
The present study was designed to explore the role of sulfur dioxide (SO2) in the regulation of vasorelaxation in the spontaneously hypertensive rat (SHR). Twenty-two Wistar rats and 15 SHRs were divided randomly into the following groups: Wistar-Kyoto (WKY) control (n = 8), WKY+Na2SO3/NaHSO3 (n = 8), WKY+
Introduction
Hypertension is the most common cardiovascular disease, but our understanding of the cellular processes involved is limited. Elucidating the physiological mechanisms responsible for hypertension is one of the most important issues in the field of medical science today. Historically, great progress has been made in research on hypertension concerning, for example, neurohumoral factors, 1 the renin–angiotensin system, 2 vasoactive peptides 3 and central nervous system control, 4 but the mechanisms of hypertension remain unclear.
Sulfur dioxide (SO2) is considered a waste gas and air pollutant.
5–7
However, biochemical metabolic studies indicate that SO2 can be derived endogenously from sulfur-containing amino acids in vivo.
8
Nitric oxide (NO) is the first gasotransmitter to be discovered in the body, and plays an important role in the pathogenesis of hypertension. Previous studies have indicated that SO2 regulates smooth muscle tone, in synergy with NO, in the aorta of Wistar rats. 16 It has also been demonstrated that SO2 increases the activity of endothelial nitric oxide synthase (eNOS) and increases expression of the eNOS gene at the transcription and translation levels in Wistar rat aorta. 17 However, the interaction between SO2 and NO in hypertensive rats has not been defined. This study was designed to explore the possible role of SO2 in the regulation of vasorelaxation in SHRs and to elucidate the possible mechanisms.
Materials and methods
Preparation of the animal model
The study protocol was approved by the Animal Research Committee of Peking University (Beijing, China).
In the in vivo part of the study, 22 male four-week-old Wistar-Kyoto (WKY) rats and 15 SHRs were divided randomly into a WKY control group (n = 8), a WKY+Na2SO3/NaHSO3 group (n = 8), a WKY+
In the in vitro part of the study, isolated thoracic aortic rings from WKY rats weighing 220–250 g and male SHRs weighing 320–350 g, all aged 12 weeks, were used to analyse the interaction between SO2 and NO. N
G-nitro-
Chemicals and solution preparation
Sodium sulfite and sodium bisulfite (Na2SO3/NaHSO3, SO2 derivatives), HDX, noradrenaline (NE), acetylcholine (Ach), sodium nitroprusside (SNP) and monobromobimane (mBrB) were purchased from Sigma (St Louis, MO, USA). Other chemicals and reagents were of analytical grade. Levels of NO were measured using an NO detection kit (nitrate reductase method), which was purchased from NIBI (Nanjing, China). Krebs–Ringer bicarbonate (pH 7.4) incubation medium contained (mmol/L): 120 NaCl, 5.5 KCl, 2.5 CaCl2, 1.2 MgSO4, 1.2 NaH2PO4, 20 NaHCO3, 0.03 EDTA-Na2 and 10
Measurement of blood pressure
The blood pressure of rats in the WKY control group, the WKY+Na2SO3/NaHSO3 group, the WKY+HDX group, the SHR control group and the SHR+Na2SO3/NaHSO3 group was measured after eight weeks of treatment by tail plethysmography (Beijing, China) while the rats were awake.
Preparation of isolated aortic rings
In the in vitro part of the study, isolated thoracic aortic rings were prepared following the procedure described in previous publications. 18,19 Briefly, male WKY rats (n = 22) and SHRs (n = 15) were anesthetized, and their thoracic aorta removed immediately and stripped of fat and connective tissues. The aortas were then cut into rings of length of approximately 3 mm. The rings were placed in a bath of Krebs buffer at pH 7.4 and 37°C under a resting optimal tension of 1 g, while 95% O2 and 5% CO2 was bubbled through the buffer from the bottom of the tube. The tension was recorded with a JZ101 Force Transducer (Beijing, China), connected to a BL-420S Data Acquisition System (Chengdu, China) during the experiment. The aortic rings in the bath tubes were equilibrated for 60 min before beginning the experiment, and the Krebs buffer was changed every 15 min. The endothelial viability and integrity of the artery were tested by the dilatory response of the rings to Ach (1 μmol/L).
Vasorelaxation experiments in vitro
Isolated thoracic aortic rings from rats of the WKY control group, the WKY+Na2SO3/NaHSO3 group, the WKY+HDX group, the SHR control group and the SHR+Na2SO3/NaHSO3 group were pre-treated with a submaximal dose of NE (1 μmol/L) to initiate contraction and then incubated with Ach (1 μmol/L) or SNP (1 μmol/L).
To explore the effect of endogenous SO2 on the vasorelaxation response to Ach, aortic rings from WKY rats were first incubated for 30 min with HDX (40, 100 or 160 μmol/L). Then, Ach (10−9−10−6 mol/L) was added cumulatively to the bath solution until the vasoconstriction induced by 10−6 mol/L NE reached a plateau at maximal tension.
Assay of NO production in rat arterial tissues
Aortic samples from rats of the WKY control group, the WKY+Na2SO3/NaHSO3 group, the WKY+HDX group, the SHR control group and the SHR+Na2SO3/NaHSO3 group were used to measure NO production. Aortic tissue NO levels were measured using an NO detection kit according to the manufacturer's protocol. The absorbance was determined at 550 nm with a UV 2100 spectrophotometer (Shimadzu, Kyoto, Japan).
Determination of SO2 content in rat arterial tissues
To measure the SO2 content of the aortic rings, the rings were first incubated with HDX (40, 100 or 160 μmol/L), and then their sulfite concentration, which indirectly represents SO2 content, was determined by high-performance liquid chromatography (HPLC) with fluorescence detection (Agilent series 1100; Agilent Technologies, Karlsruhe, Germany).
20
In brief, 100 μL of sample was mixed with 70 μL of 0.212 mol/L sodium borohydride in 0.05 mol/L Tris-HCl (pH 8.5) and incubated at room temperature for 30 min. The sample was then mixed with 10 μL of 70 mmol/L mBrB in acetonitrile, incubated for 10 min at 42°C and mixed with 40 μL of 1.5 mol/L perchloric acid. Protein precipitates in the mixture were removed by centrifugation at 12,400 ×
Statistical analysis
Results were expressed as mean ± SD. Statistical analysis was performed using SPSS 11.5 (SPSS, Chicago, IL, USA). Student's t-test for unpaired samples was used to compare results between two groups. For multiple group comparisons, analysis of variance followed by a post hoc analysis (Student–Newman–Keuls test) was used. Statistical significance was set at P < 0.05.
Results
The change of blood pressure in 12-week rats (mean ± SD)
WKY, Wistar-Kyoto; SHR, spontaneously hypertensive rat; HDX,
*Compared with WKY group, P < 0.05
**Compared with SHR group, P < 0.05
SO2 increased the vasorelaxant response to Ach in isolated aortic rings
The vasorelaxation to Ach and SNP (mean ± SD)
Ach, acetylcholine; SNP, sodium nitroprusside; WKY, Wistar-Kyoto; SHR, spontaneously hypertensive rat; HDX,
*Compared with WKY group, P < 0.05
**Compared with SHR group, P < 0.05
Next, we tested the vasorelaxant response to Ach after pre-incubation with HDX in vitro. Figure 1 shows the vasorelaxant response of aortic rings from WKY rats to Ach (10−9−10−4 mol/L) after pre-incubation with HDX (40, 100 or 160 μmol/L). The response decreased significantly following pre-incubation with 40 and 100 μmol/L HDX, with an EC50 of 0.42 ± 0.17 and 1.09 ± 0.45 μmol/L, respectively, compared with that of the control group (EC50 0.08 ± 0.01 μmol/L, P < 0.05). However, the EC50 showed no significant difference between the above two concentrations (P > 0.05). The vasorelaxant response to Ach following pre-incubation with 160 μmol/L HDX was significantly reduced, with an EC50 of 16.42 ± 12.45 μmol/L, compared with pre-incubation with 40 μmol/L or 100 μmol/l HDX (P < 0.05).
The vasorelaxation to Ach after incubation with HDX. The aortic rings were pre-incubated without HDX (black, -□- control) (n = 8), and with 40 μmol/L HDX (orange, -▪-) (n = 7), 100 μmol/L HDX (blue, -▴-) (n = 8), 160 μmol/L HDX (pink, -♦-) (n = 7) for 30 min and then in buffer containing 1 μmol/L NE to induce its constriction. Ach (10−6∼10−9 mol/L) was cumulatively added to the bath solution when the vasodilator curve of the ring reached a plateau phase. aCompared with control, P < 0.05; bcompared with HDX 40 μmol/L group, P < 0.05; ccompared with HDX 100 μmol/L group, P < 0.05. Ach, acetylcholine; HDX, 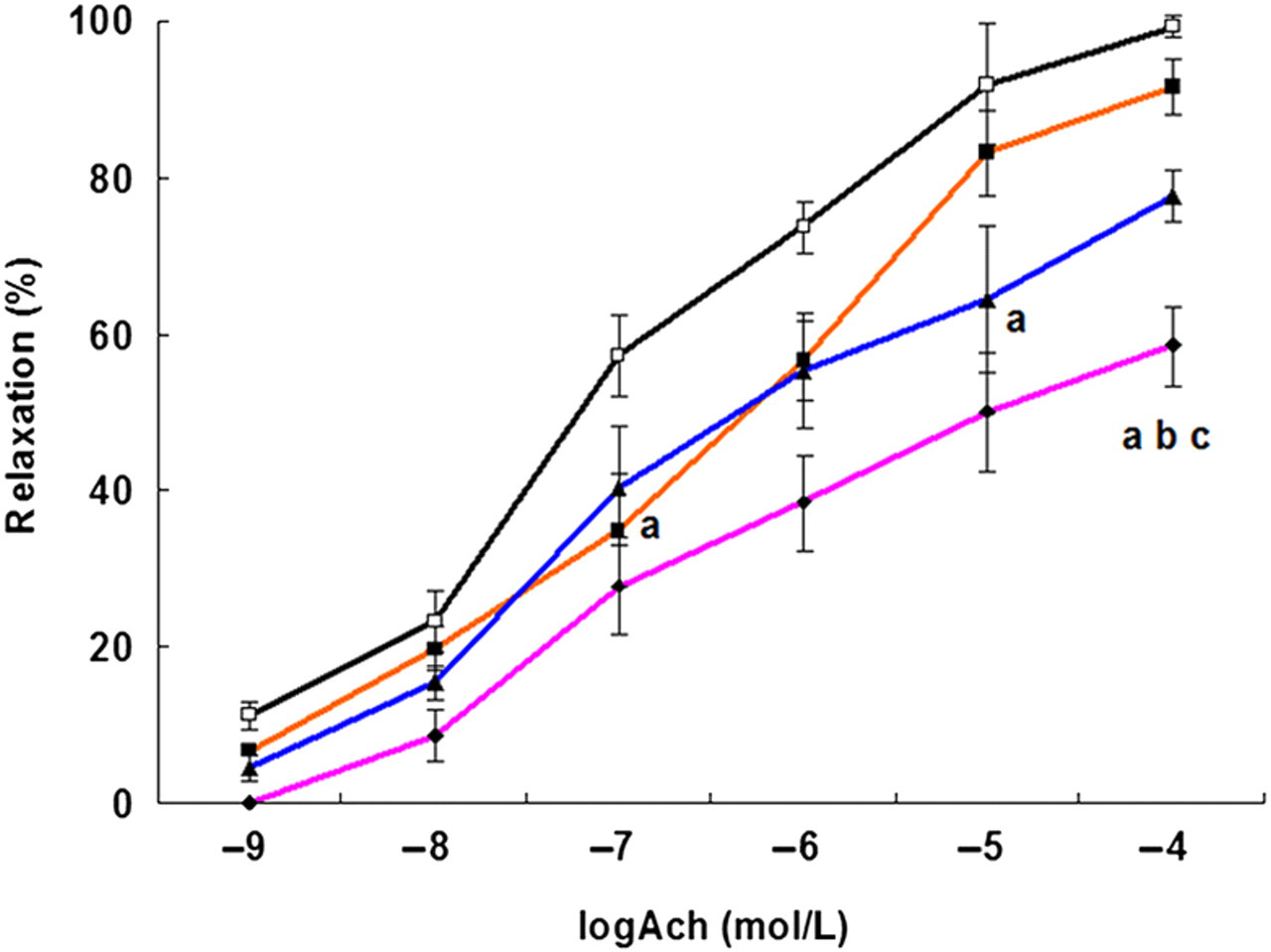
Figure 2 shows SO2 levels in rat aortas after pre-incubation with 40, 100 or 160 μmol/L HDX. At all three concentrations of HDX, SO2 levels were significantly reduced compared with those of the control group (P < 0.05), but there was no significant difference in SO2 levels between 40 and 100 μmol/L HDX (P > 0.05). SO2 levels in arteries preincubated with 160 μmol/L HDX were decreased significantly compared with those in arteries pre-incubated with 40 or 100 μmol/L HDX (P < 0.05).
SO2 level in rat artery after pre-incubating with 40 μmol/L HDX (n = 6), 100 μmol/L HDX (n = 6), 160 μmol/L HDX (n = 6) and in the artery of control group (n = 8). aCompared with control, P < 0.05; bcompared with HDX 40 μmol/L group, P < 0.05; ccompared with HDX 100 μmol/L group, P < 0.05. SO2, sulphur dioxide; HDX, 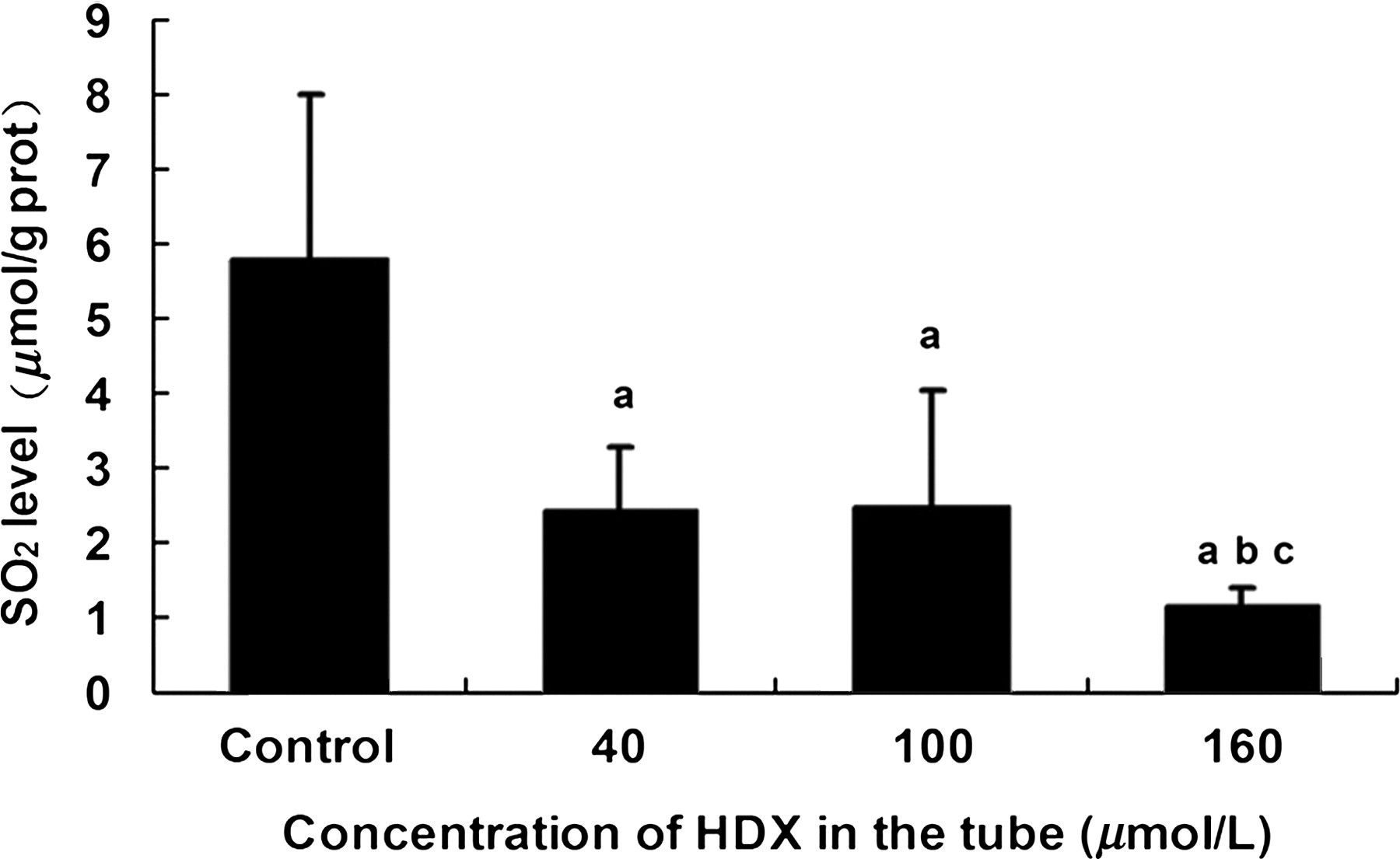
SO2 increased vasorelaxant response to SNP in isolated aortic rings from rats of different groups
To understand how SO2 increased the vasorelaxant response in isolated aortic rings, we first tested whether SO2 increased the vasorelaxant response to SNP. The results showed that the vasorelaxant response to SNP was lower in the WKY + HDX group than in the WKY control group (P < 0.05) and was markedly decreased in the SHR group compared with the WKY control group (P < 0.05). However, the vasorelaxant response to SNP was notably enhanced in the SHR+Na2SO3/NaHSO3 group compared with the SHR control group (P < 0.05) (Table 2).
Role of NO in SO2-induced vasorelaxation in rat arterial tissues
We next investigated whether SO2 affects the generation of NO in arterial tissues. Figure 3 shows that the NO level in aortic tissues from the WKY+HDX group was decreased significantly compared with that in the WKY control group (P < 0.05). However, the NO level in the aortic tissues of the SHR+Na2SO3/NaHSO3 group was increased significantly compared with that of the SHR control group (P < 0.05). We also measured the SO2-induced vasorelaxant response after incubation with NO levels in artery from rats of WKY control group (n = 8), WKY+Na2SO3/NaHSO3 group (n = 8), WKY+HDX group (n = 6), SHR control group (n = 8) and SHR+Na2SO3/NaHSO3 group (n = 7). aCompared with WKY group, P < 0.05; bcompared with SHR group, P < 0.05. NO, nitric oxide; WKY, Wistar-Kyoto; SHR, spontaneously hypertensive rat; HDX, The vasorelaxation to SO2 derivative on aortic rings after incubation with 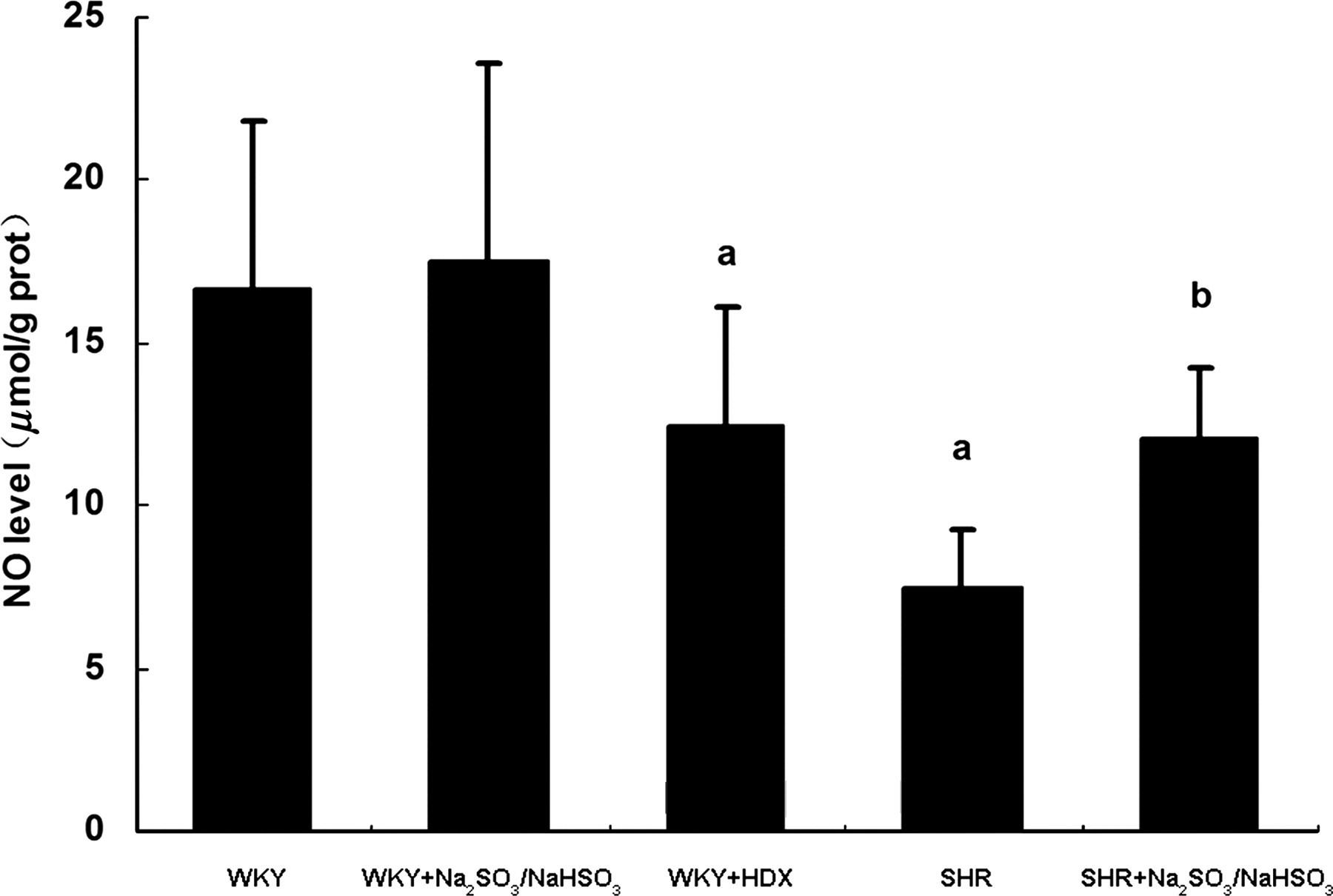
Discussion
NO, carbon monoxide, hydrogen sulfide and SO2 have been identified as toxic environmental pollutants. However, they can be synthesized in the body from arginine, glycine and cysteine, respectively, and serve as important signaling molecules in animals and humans. 16 The reference range for total serum sulfite (SO2 derivative) is 0–10 μmol/L in healthy individuals. 21 It was reported that SO2 derivatives (sodium sulfite and sodium bisulfite, about 3 and 1 mol/L, respectively, pH 7.0) administered via intraperitoneal injection decreased the blood pressure of male Wistar rats. 22 There have been studies concerning the vasodilatory effect of SO2 derivatives on isolated aortic rings from WKY rats in vitro. 23,24 Our previous study demonstrated that vascular endogenous SO2 was reduced in rats with hypertension. 14 In this study, we found that the blood pressure of SHRs was significantly decreased when SO2 derivatives were given by intraperitoneal injection for eight weeks, which further suggests that SO2 plays a role in the development of hypertension.
Abnormal vasorelaxation and vascular structural remodeling are the principal components of the pathogenesis of hypertension. 25,26 Imai et al. reported the reaction of dimerized NO with SO2 in a restricted slit-shaped micropore, 27 which suggested a possible interaction between NO and SO2 in the regulation of vasorelaxation in SHRs.
Ach could activate M carinic receptors on endothelial cells, resulting in NO release and a consequent decrease in the tension of vascular smooth muscle cells. HDX, as an inhibitor of SO2-producing enzymes, inhibits the generation of endogenous SO2. When investigating the possible mechanisms by which SO2 regulates vasorelaxation, we found that the vasorelaxant response to Ach in the WKY+HDX and SHR groups was significantly decreased compared with that in the WKY control group. However, the vasorelaxant response to Ach in the SHR+Na2SO3/NaHSO3 group was significantly increased compared with the SHR control group. We also used isolated aortic rings from WKY rats to investigate the vasorelaxant response to Ach after pre-incubation with HDX; this was markedly decreased, indicating that HDX significantly reduced SO2 production in the aortic rings.
SNP is a NO donor that directly relaxes blood vessels. We found that the vasorelaxant response to SNP in the WKY+HDX group was significantly decreased compared with both the WKY control group and the SHR group. However, the vasorelaxant response to SNP in the SHR+Na2SO3/NaHSO3 group was significantly increased compared with the SHR control group. These results suggested that SO2 possibly improved NO-induced vasorelaxation in SHRs.
In previous studies by Li et al., 17 it was found that SO2 could increase NO production in Wistar rats. The vascular endothelium can produce a wide array of vasoactive substances, including gaseous NO, that modulate the tone of vascular smooth muscle cells. 28,29 We therefore investigated whether SO2 can promote the generation of NO in the blood vessels of SHRs and whether SO2 can relax those vessels by increasing NO production. Our results showed that the NO level in the aorta decreased notably in the WKY+HDX and SHR groups compared with the WKY controls. However, the NO level was reversed in aortas from the SHR+Na2SO3/NaHSO3 group. These results suggested that SO2 increased NO production, which in turn facilitated vasorelaxation in the rats in the SHR+Na2SO3/NaHSO3 group.
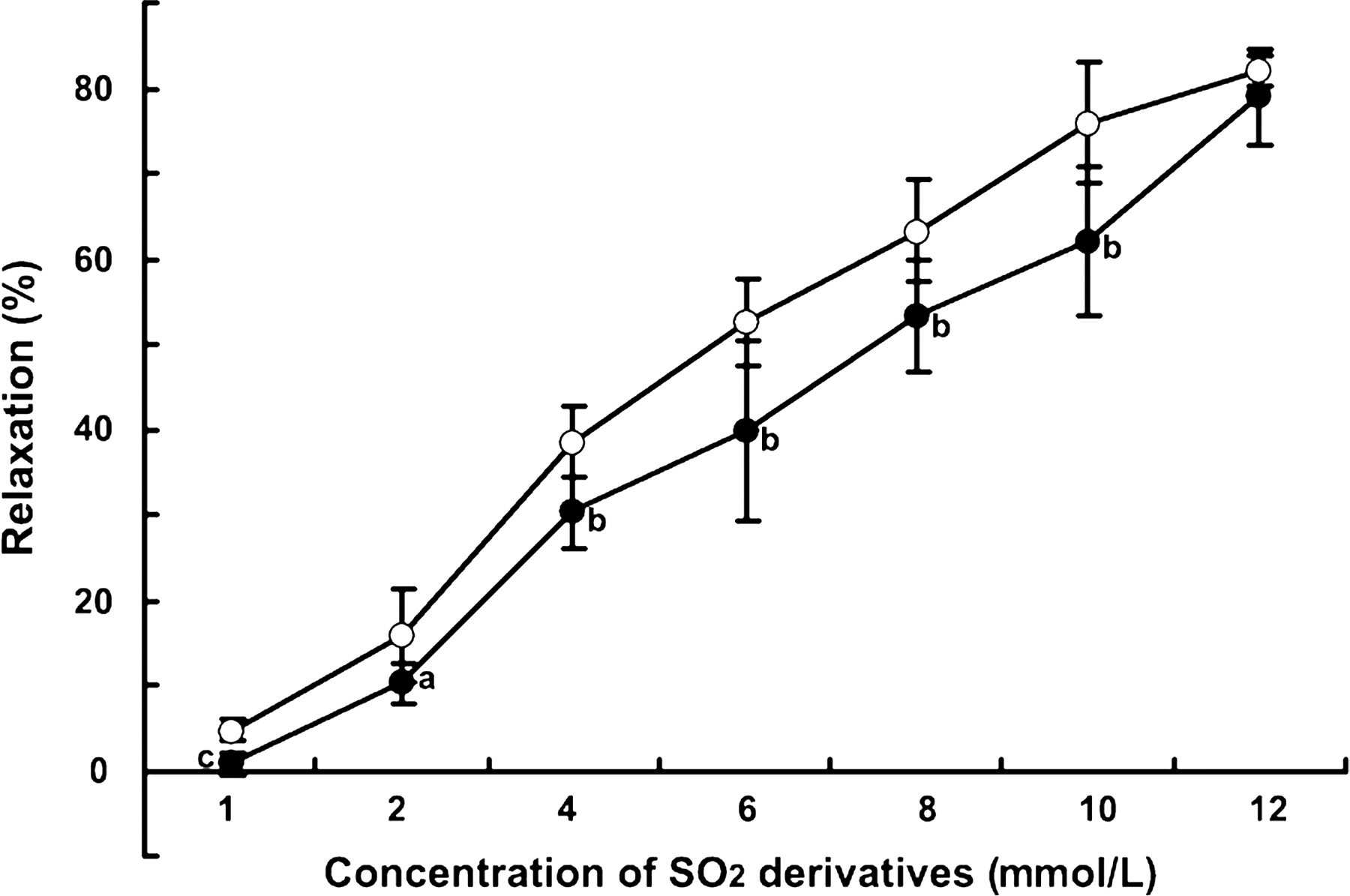
We also examined vasorelaxation in aortic rings from Wistar rats after pre-incubation with the NO inhibitor
In summary, our study indicates that SO2 reduces blood pressure and increases vasorelaxation in the arteries of SHRs. SO2 could produce these effects by enhancing the vasorelaxant response to NO in isolated aortic rings and increasing the NO level in aortic tissues.
Footnotes
ACKNOWLEDGEMENTS
This study was supported by the Major Basic Research Program of China (Nos 2012BA103B03 and 2011CB503904), the National Natural Science Foundation of China (Nos 81121061, 81070111 and 31130030) and the Beijing Natural Science Foundation (No. 7112130).