Chronic granulomatous disease (CGD) causes impaired hydrogen peroxide (H2O2) generation. Consequently, neutrophils in patients with CGD fail to kill infecting pathogens. We expected that supplementation with H2O2 would effectively restore the bactericidal function of neutrophils in CGD. Here, we used polyethylene glycol-conjugated d-amino acid oxidase (PEG-DAO) as an H2O2 source. The enzyme DAO generates H2O2 by using d-amino acid and oxygen as substrates. PEG-DAO plus d-amino acid indeed exerted bacteriostatic activity against Staphylococcus aureus via H2O2in vitro. Furthermore, use of PEG-DAO plus d-amino acids, which increased the amount of intracellular H2O2, restored bactericidal activity of neutrophils treated with diphenylene iodonium, in which nicotinamide adenine dinucleotide phosphate (NADPH) oxidase was defective. This restoration of bactericidal activity was mediated by myeloperoxidase, with concomitant production of H2O2 by PEG-DAO plus d-Ala. We also confirmed that PEG-DAO treatment restored bactericidal activity of congenitally defective neutrophils from patients with CGD. These results indicate that PEG-DAO can supply additional H2O2 for defective NADPH oxidase of neutrophils from patients with CGD, and thus neutrophils regain bactericidal activity.
Chronic granulomatous disease (CGD) is a genetic disorder characterized by chronic and recurrent pyogenic infections. Patients with CGD have a defect in nicotinamide adenine dinucleotide phosphate (NADPH) oxidase that results in dysfunctional production of hydrogen peroxide (H2O2).1,2 H2O2 plays a pivotal role in the antibacterial function of neutrophils, mediated by myeloperoxidase (MPO), so the impaired H2O2 production means failure of bactericidal activity against pathogenic organisms such as Staphylococcus aureus.2,3
d-Amino acid oxidase (DAO) is an enzyme containing flavin adenine dinucleotide (FAD).4 The biochemical function of DAO involves oxidative deamination of d-amino acids, which yields the corresponding α-keto acids, a process in which molecular oxygen is used as an electron acceptor and H2O2 is generated.5
We previously prepared polyethylene glycol (PEG)-conjugated DAO (PEG-DAO) with comparable enzyme activity to native DAO.6,7 More importantly, PEG-DAO had a longer circulation time in the blood, and preferential accumulation in inflamed sites, as a result of the enhanced permeability and retention (EPR) effect.8,9 Our previous report showed that PEG-DAO exhibited selective cytotoxicity against various cancer cells via production of H2O2in vivo and in vitro.6,7
We therefore anticipated that PEG-DAO would function as an alternative supplier of H2O2 for neutrophils in patients with CGD. In this study, we therefore investigated the effect of PEG-DAO on bactericidal activity of neutrophils from mice in which NADPH oxidase was inhibited, and from a patient with CGD, and analyzed the mechanism of bactericidal activity, in addition to investigating MPO-inhibited neutrophils.
Materials and methods
Materials
S. aureus strain ATCC25923 was used in these studies. ICR mice were purchased from Japan SLC, Inc., Shizuoka, Japan. Trypticase soy (SCD) broth was purchased from Nissui Seiyaku Co., Tokyo, Japan. Flavin adenine dinucleotide was purchased from Sigma-Aldrich Chemical Co. (St Louis, MO, USA). Trypticase soy agar, isopropyl-β-d-thiogalactopyranoside, carbenicillin, Tween-20, ammonium sulfate, casein sodium salt and other reagents were from Wako Pure Chemical Industries, Ltd, Osaka, Japan. 4-aminobenzoic acid hydrazide (4-ABH) was from Merck KGaA, Frankfurt, Germany. Diphenylene iodonium (DPI) was purchased from Tokyo Chemical Industry Co., Ltd, Tokyo, Japan. Succinimide-activated PEG (MEC-50HS), with an average molecular size of Mr 5000, was purchased from Nippon Oil & Fat Co. (Tokyo, Japan).
Preparation of PEG-DAO
Recombinant porcine DAO was prepared as described previously.7 Briefly, Escherichia coli BL21 (DE3) bacteria harboring the pET3c plasmid encoding porcine DAO were cultured in LB medium containing 50 μg/mL carbenicillin, and porcine DAO expression was achieved by adding 10 μmol/L isopropyl-β-d-thiogalactopyranoside to the medium with E. coli. After culture of the bacteria at 37°C for 20 h, bacterial pellets were sonicated (150 W, 30 min) in 17 mmol/L pyrophosphate buffer (pH 8.2), and porcine DAO was obtained by heat denaturation at 59°C for three minutes, followed by ammonium sulfate precipitation at 35% saturation, and then diethylaminoethyl cellulose column chromatography (L = 10 cm × ϕ = 1.6 cm). The purity of DAO (>90%) was determined by using sodium dodecylsulfate polyacrylamide gel electrophoresis after staining with Coomassie brilliant blue. PEGylation of DAO was conducted as described previously.6 In brief, to the DAO solution (2.0 mg/mL protein in 50 mmol/L sodium phosphate buffer, pH 7.4), succinimide-activated PEG was added at a 3.5 mol/L excess of PEG/mol of free amino groups in DAO and was allowed to react for one hour at 4°C. The reaction mixture containing PEG-DAO thus obtained was then purified to remove free PEG and other low-molecular-weight reactants by ultrafiltration with the YM-10 membrane (Millipore) using 10 times the volume of 10 mmol/L phosphate-buffered saline (PBS). PEG-DAO was stored in PBS containing 0.1 mmol/L FAD at 4°C. Approximately 30% of the amino groups on DAO was reacted with PEG.
Bacteriostatic assay
S. aureus bacteria were cultured until the mid-log phase of growth in SCD broth with reciprocal shaking at 37°C. S. aureus were washed twice in saline and 1 × 106 CFU/mL of S. aureus were incubated with various concentrations of PEG-DAO, d-Ala, and with or without catalase in SCD broth at 37°C for five hours. The relative total numbers of bacteria were measured at turbidity at 570 nm and were correlated with the numbers of viable bacteria.
Preparation of neutrophils
Peritoneal neutrophils were elicited in 10-week-old female ICR mice by intraperitoneal injection of 3 mL per mouse of 6% casein sodium salt dissolved in physiological saline. At six hours after injection, neutrophils were harvested via peritoneal lavage with 5 mL of PBS, pH 7.4. Contaminating erythrocytes were removed by incubating in hypotonic saline solution (0.2% NaCl) for 30 s to cause erythrocytes to burst, after which isotonicity was restored via a rebalancing solution (1.9% NaCl) followed by centrifugation. Approximately 1 × 107 neutrophils were obtained from 10-week-old female ICR mice. The purity of the neutrophils (>90%) was checked by using Giemsa staining and examination of cell morphology with a conventional microscope (ECLIPSE TS100; Nikon, Tokyo, Japan). Human peripheral neutrophils were collected from a patient with CGD and a healthy volunteer using Polymorphprep™ (Cosmo Bio, Tokyo, Japan) according to the manufacturer's instruction. Briefly, 5 mL of human blood sample was carefully layered on the top of 5 mL of Polymorphprep™, followed by centrifugation with a swing-out rotor for 30 min at 450 × g. The neutrophil fraction was collected and mixed with 0.45% NaCl, and then centrifuged for 10 min at 400 × g. Neutrophil pellets were then resuspended in PBS (−) and used for further experiments.
Bactericidal activity of neutrophils and preparation of CGD neutrophil mimics
Mouse peritoneal neutrophils were preincubated with 10 DPI or 10 μmol/L 4-ABH at 37°C for 15 min. S. aureus, which were cultured in SCD broth until the mid-log phase growth, were treated with 10% pooled mouse serum for effective neutrophilic endocytosis of S. aureus. Bacteria were added to neutrophils at the bacteria-to-neutrophil ratio of 10:1 (1 × 106 neutrophils/mL), and incubation proceeded at 37°C with reciprocal shaking at 0.5 Hz. After 30 min of incubation, non-phagocytosed bacteria were removed by swing-out centrifugation (at 110 × g, 4 min) and neutrophils were washed three times with PBS (+) containing 10 μmol/L DPI. Phagocytosed bacteria were precipitated with neutrophils, but non-phagocytosed bacteria were retained in the supernatant. Neutrophils that ingested the bacteria were incubated at 37°C for 30 min with shaking, with increasing concentrations of PEG-DAO (10, 50 and 100 mU/mL) in the presence of 10 mmol/L d-Ala and PBS (+) containing 10 μmol/L DPI. Samples were diluted with 0.2% Tween-20, incubated at room temperature for five minutes to release phagocytosed bacteria, and vortexed vigorously, after which duplicate 100-μL aliquots were plated on 15 mL plates of SCD agar gel followed by overnight culture at 37°C. The numbers of viable bacteria were counted as described above.
Results
Bacteriostatic activity of PEG-DAO
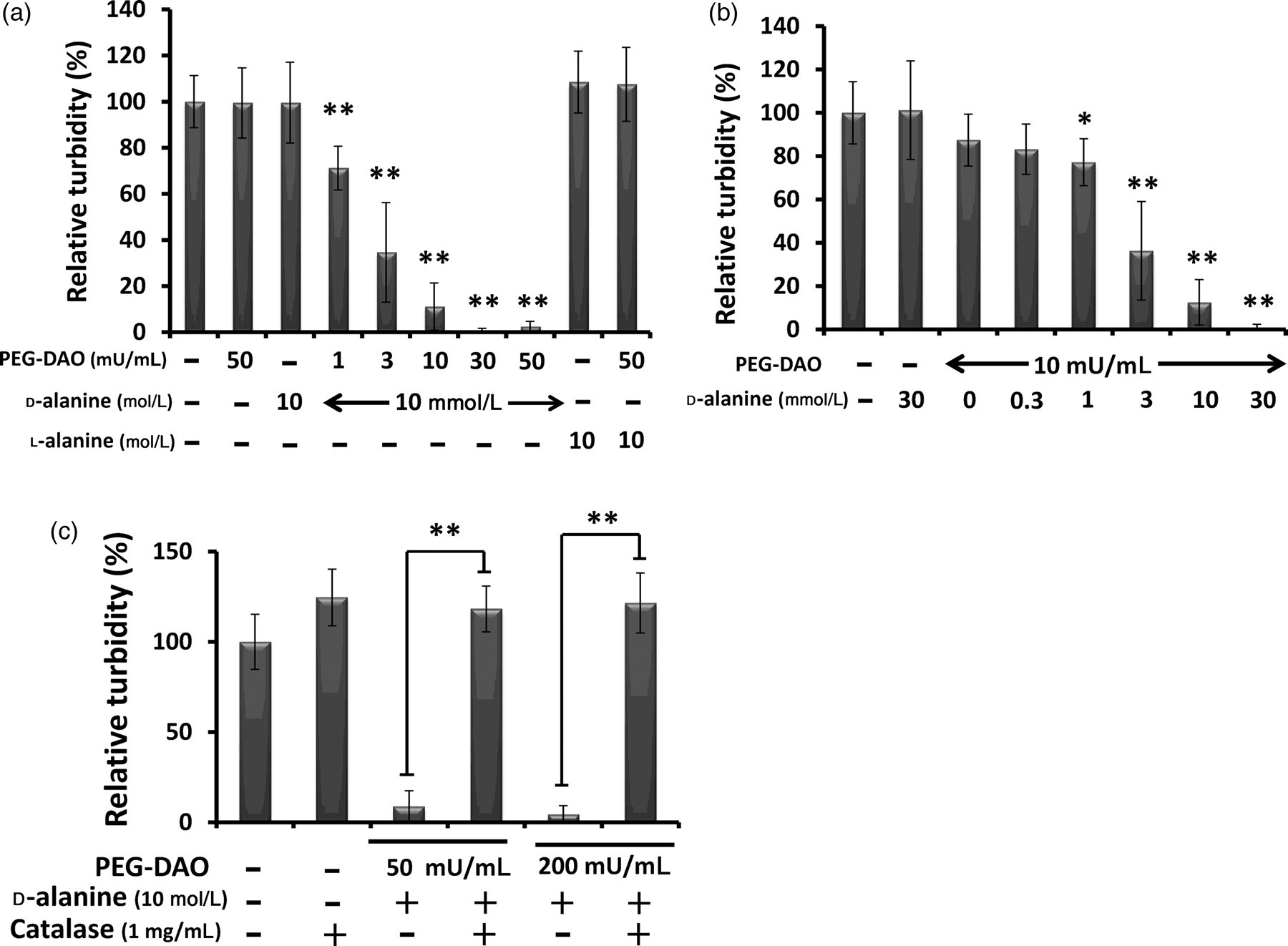
We first examined the bacteriostatic activity of PEG-DAO against S. aureus. In the presence of 10 mmol/L d-Ala, PEG-DAO showed bacteriostatic activity in a dose-dependent manner (Figure 1a). Different concentrations of d-Ala also demonstrated dose-dependent activity (Figure 1b). Bacteriostatic activity was not observed with treatment of PEG-DAO and l-Ala (Figure 1a). Adding 1 mg/mL catalase (5000–15,000 U/mL) to samples with PEG-DAO and d-Ala nullified the bacteriostatic activity of PEG-DAO (Figure 1c).
Bacteriostatic activity of polyethylene glycol-conjugated d--amino acid oxidase (PEG-DAO) against Staphylococcus aureus. (a) Increasing concentrations of PEG-DAO in the presence of 10 mmol/L d-Ala or (b) increasing concentrations of d-Ala in the presence of 10 mU/mL PEG-DAO were incubated with S. aureus bacteria (1 × 106 CFU/mL) for five hours, after which turbidity at 570 nm was measured. (c) S. aureus bacteria (1 × 106 CFU/mL) were incubated for five hours with PEG-DAO plus d-Ala with or without 1 mg/mL bovine catalase, and after which turbidity at 570 nm was measured. ** and * indicate statistically significant differences (P< 0.01) and (P< 0.05), respectively, by Student's t-test. Values are means ± SD (n = 12)
PEG-DAO restored antibacterial activity of NADPH oxidase-deficient mouse neutrophils
As H2O2 is a highly cell permeable oxidant, we examined H2O2 generated by PEG-DAO plus d-Ala in the medium to determine whether H2O2 could penetrate cell membranes and enter the cells. H2O2 treatment increased oxidative stress inside neutrophils, as shown by analysis using fluorescence flow cytometry in the presence of the fluorescent reactive oxygen species probe dichlorofluorescein diacetate (DCFH-DA) (Figure 2a). In the presence of 10 mmol/L d-Ala, treatment with PEG-DAO increased the intracellular H2O2 concentration in a dose-dependent manner as judged by this fluorescent probe (Figures 2b and c). However, PEG-DAO alone did not induce oxidative stress (data not shown). To mimic defective CGD neutrophils, mouse peritoneal neutrophils were treated with 10 μmol/L DPI to inhibit NADPH oxidase. DPI treatment increased the numbers of viable bacteria inside the treated neutrophils, as seen by the colony forming assay (Figure 2d). However, PEG-DAO treatment in the presence of 10 mmol/L d-Ala restored bactericidal activity of DPI-treated defective neutrophils; the number of viable bacteria inside neutrophils almost recovered to the normal, non-CGD control. No enhancement of antibacterial activity by PEG-DAO in normal neutrophils was observed (Figure 2d).
Increase in intracellular hydrogen peroxide (H2O2) by means of treatment with polyethylene glycol-conjugated d-amino acid oxidase (PEG-DAO) plus d-Ala and restoration of bactericidal activity of chronic granulomatous disease-like neutrophils. Mouse peritoneal neutrophils were pretreated with 10 μmol/L dichlorofluorescein diacetate, a fluorescent molecular probe for H2O2, and were then incubated with (a) increasing concentrations of H2O2 or (b) increasing concentrations of PEG-DAO (1–100 mU/mL) plus a fixed amount of 10 mmol/L d-Ala, followed by incubation for 30 min at room temperature. The fluorescence intensity of neutrophils was measured by means of a flow cytometer. (c) Normalized fluorescence intensity of cells in (b) representing intracellular reactive oxygen species. (d) Mouse peritoneal neutrophils were pretreated with 10 μmol/L diphenylene iodonium (DPI) and were then incubated with opsonized Staphylococcus aureus to allow phagocytosis for 30 min. After removal of non-phagocytosed bacteria in the supernatant, neutrophils were treated with PEG-DAO plus 10 mmol/L d-Ala for 30 min. Viable bacteria inside the neutrophils were counted as described in Materials and methods. Values are means ± SE (n = 4). **Indicates statistically significant differences (P< 0.01) by Student's t-test
PEG-DAO induced MPO-dependent bactericidal activity of CGD-equivalent mouse neutrophils
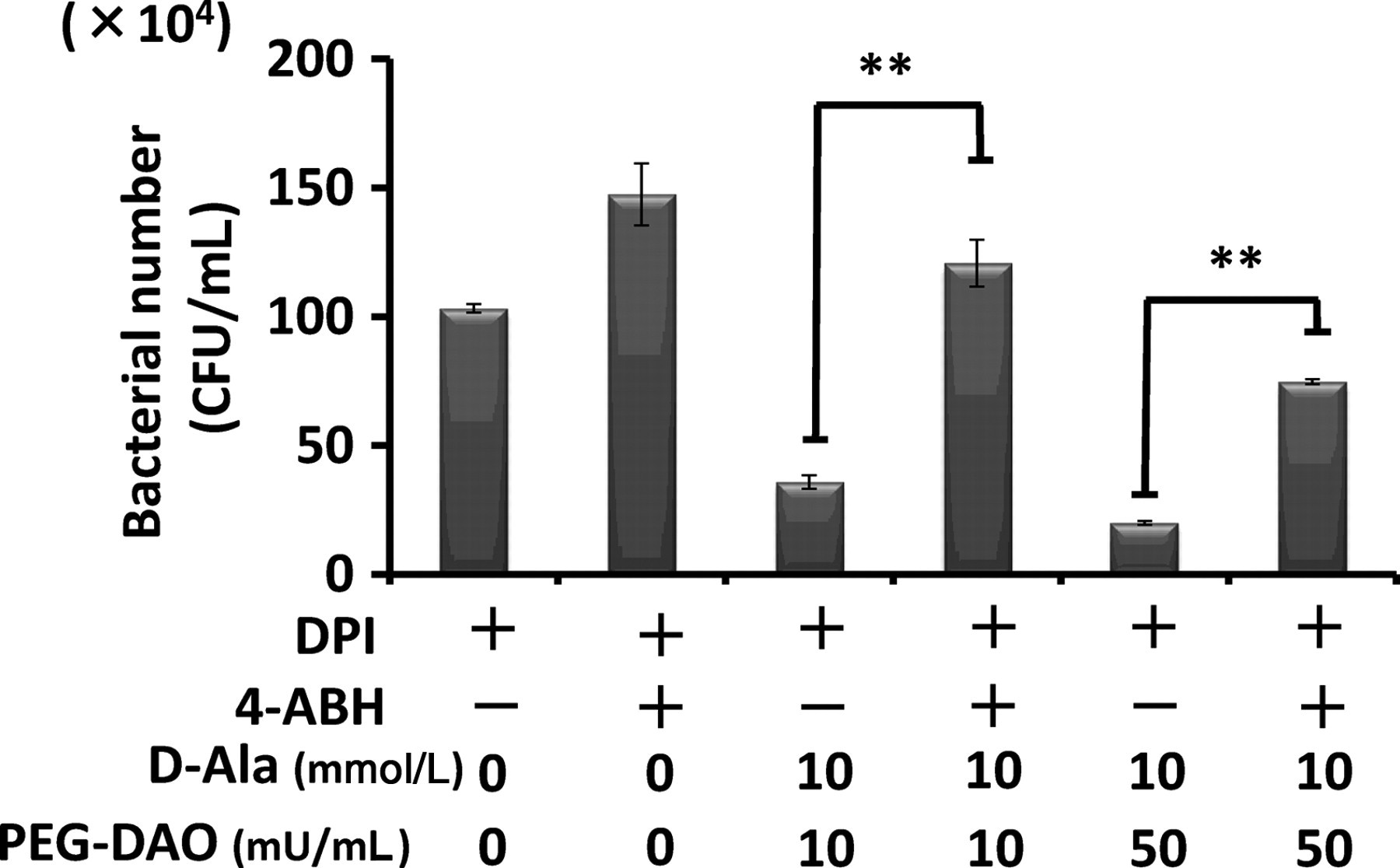
Consistent with the results shown in Figure 2, the bactericidal activity of mouse peritoneal neutrophils, which had been pretreated with DPI and thus had no NADPH oxidase activity, was restored by adding PEG-DAO and d-Ala (Figure 3). Treatment with 4-ABH, a specific inhibitor of MPO, clearly suppressed the bactericidal activity of normal mouse neutrophils. An interesting finding was the significant suppression of bactericidal activity of neutrophils by 4-ABH treatment, even with added PEG-DAO and d-Ala. This result demonstrates the important role of MPO in PEG-DAO-mediated bacterial killing (Figure 3).
Polyethylene glycol-conjugated d-amino acid oxidase (PEG-DAO) restored the bactericidal activity of diphenylene iodonium (DPI)-treated neutrophils, which depended on myeloperoxidase (MPO) activity. Mouse peritoneal neutrophils were treated with both DPI, an inhibitor of nicotinamide adenine dinucleotide phosphate oxidase, and 4-aminobenzoic acid hydrazide (4-ABH), an inhibitor of MPO. Cells were then incubated with opsonized Staphylococcus aureus to allow phagocytosis. After removal of non-phagocytosed bacteria, neutrophils were treated with PEG-DAO for 30 min. The number of viable bacteria was derived from the count of colonies on agar plates, as described in Materials and methods. Values are means ± SE (n = 4). **Indicates statistically significant differences (P< 0.01) by Student's t-test
Restoration of bactericidal activity of human CGD neutrophils by PEG-DAO treatment
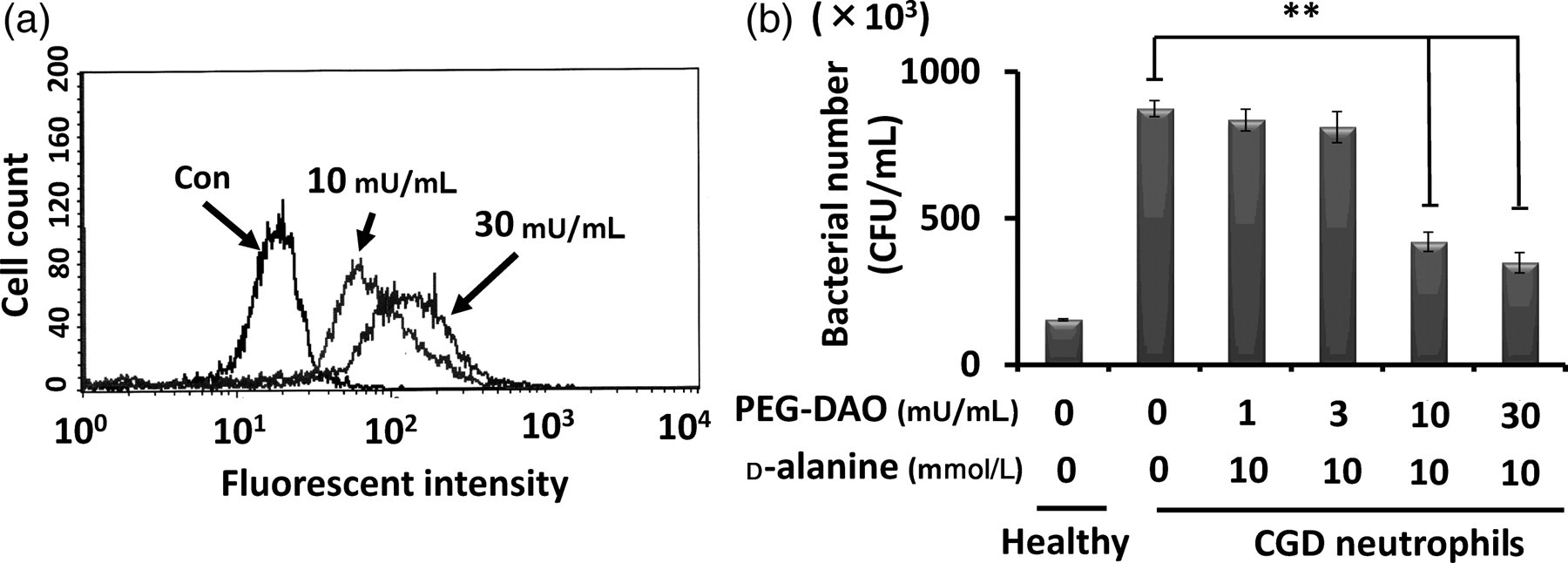
We examined the bactericidal activity of PEG-DAO with neutrophils from a patient with CGD, as they are known to have little antibacterial effect. We first checked the intracellular oxidant status before and after PEG-DAO treatment. In agreement with the results shown in Figure 2, treatment with PEG-DAO plus d-Ala increased the intracellular oxidative state (Figure 4a). CGD neutrophils showed less bactericidal activity as compared with healthy neutrophils. However, treatment with 10 or 30 mU/mL PEG-DAO restored the bactericidal activity of these CGD neutrophils to a considerable extent in the presence of 10 mmol/L d-Ala (Figure 4b).
Restoration of bactericidal function of human chronic granulomatous disease (CGD) neutrophils by polyethylene glycol-conjugated d-amino acid oxidase (PEG-DAO) treatment. (a) Human CGD neutrophils were pretreated with 10 μmol/L dichlorofluorescein diacetate, an intracellular marker of oxystress, and were then incubated with PEG-DAO (10 or 30 mU/mL) and 10 mmol/L d-Ala for 30 min at room temperature. The fluorescence intensity of the neutrophils was measured by using a fluorescence flow cytometer. (b) CGD neutrophils were incubated with Staphylococcus aureus to allow phagocytosis. After removal of the non-phagocytosed S. aureus by centrifugation, neutrophils were treated with PEG-DAO and d-Ala for 30 min followed by incubation with 0.2% Tween-20 to liberate bacteria from the neutrophils. Serial dilutions of bacteria were then mixed with Trypticase soy agar in Petri dishes and incubated overnight at 37°C, and bacterial colonies was counted. Values are means ± SE (n= 3). **Indicates statistically significant differences (P< 0.01) by Student's t-test
Discussion
Patients with CGD have a genetic defect in NADPH oxidase, which results in deficient H2O2 generation and hence poor antimicrobial defense. Consequently, neutrophils from patients with CGD fail to kill bacteria, and chronic inflammation may result.2,3 CGD neutrophils continue to evidence normal migration, phagocytosis and MPO enzyme activity; only their H2O2-generating capacity is severely impaired.10 We thus hypothesized that supplementation with H2O2 would restore this function of CGD neutrophils.
H2O2 itself shows bactericidal activity in vitro, but intravenous injection of H2O2 does not result in antibacterial activity against bacteria in vivo because of the presence of excess catalase in the systemic circulation.11 In other words, circulating H2O2in vivo is rapidly cleared by catalase, so no H2O2 is available for targeting to infected or inflamed sites. Furthermore, H2O2 may harm mammalian cells that have no catalase or other antioxidants.11 Thus, new strategies to deliver the H2O2 to the target site are required, which may provide a new strategy of H2O2-dependent treatment of infection and cancer.
To achieve the delivery of H2O2 to an infected or inflamed site, we prepared PEG-DAO which retained a degree of H2O2-generating activity that was comparable with that of native DAO.6,7 Also, DAO derived from a porcine source can be used in humans, because PEGylation reduces the antigenicity of this enzyme. PEG-DAO accumulates preferentially in tumor tissue because of the EPR effect, the mechanism of which is based on the highly enhanced extravasation of macromolecules in the tumor and inflamed tissues. In addition, impaired lymphatic clearance of such macromolecules from interstitial space makes this effect more distinct. This increased vascular permeability is induced partly by overproduction of inflammatory vascular mediators such as bradykinin, nitric oxide and many others.2,8
The concentrations of d-amino acids, which are substrates of PEG-DAO, are extremely low in mammalian blood plasma, so H2O2 generation by PEG-DAO alone in systemic circulation is quite limited. However, we can induce H2O2 generation with an intravenous injection of d-amino acids. PEG-DAO at first accumulates predominantly at the inflamed site because of the EPR effect. After several hours of PEG-DAO infusion via an intravenous route allowing PEG-DAO to accumulate more selectively at the disease site, at that time, the PEG-DAO concentration in blood is very low, and d-amino acid is infused subsequently via the intravenous route. Thus, PEG-DAO is preferable for delivery of H2O2 to the inflamed sites or cancer tissue, and avoids systemic generation of H2O2.6
PEG-DAO plus d-Ala, as mediated by H2O2, showed bacteriostatic activity against S. aureus in a dose-dependent manner via production of H2O2 (Figure 1). These results indicate that PEG-DAO can serve as an antibacterial agent if it is selectively delivered to an infected site. This delivery became a possibility as a result of the prolonged plasma half-life of PEG-DAO and the EPR effect.
When PEG-DAO plus d-Ala was supplied to NADPH oxidase-deficient neutrophils, it increased the amount of H2O2 inside the cells (Figures 2 and 4). The fluorescent oxystress probe DCFH-DA, which effectively enters neutrophils, contains a diacetate group that is quickly hydrolyzed, so DCFH-DA becomes reactive to H2O2 and then fluorescent.12 The higher fluorescence intensity of DCFH-DA thus indicates a higher oxidative state inside cells, but does not reflect the oxidative state outside cells. As expected, PEG-DAO treatment or H2O2 treatment increased the intracellular level of H2O2 in a dose-dependent manner (Figure 2a). Furthermore, in the presence of d-Ala, addition of PEG-DAO also increased the amount of intracellular H2O2 in a dose-dependent manner (Figures 2a and b). These results clearly indicate that PEG-DAO treatment can supply exogenous H2O2 efficiently to neutrophils and that this restored level of intracellular H2O2, as H2O2 is converted to the hypochlorite ion by MPO, would facilitate potent bactericidal activity of neutrophils from a CGD patient.
We also examined whether PEG-DAO treatment would restore the bactericidal activity of CGD-like neutrophils. We prepared neutrophil mimics, which were similar to neutrophils in patients with CGD, by pretreatment with 10 μmol/L DPI, which achieves its effects by inhibiting NADPH oxidase and thus suppressing H2O2 generation.13 In this experiment, we used PEG-DAO plus d-Ala and examined the effect of this treatment on phagocytosed bacteria. We observed a significant increase in the number of viable bacteria inside these neutrophils after DPI treatment (Figure 2d). In this setting, the bactericidal activity of DPI-treated neutrophils was similar to that of neutrophils from a CGD patient. However, PEG-DAO treatment of DPI-treated neutrophils greatly suppressed the number of viable bacteria inside neutrophils, almost to the number in healthy neutrophils (Figure 2d).
Most MPO exists in vacuoles in neutrophils,14 and MPO oxidizes the chloride ion, with H2O2, to produce hypochlorous acid, one of the most potent bactericidal molecules in biological systems. We therefore hypothesized that restoration of bactericidal activity of DPI-treated neutrophils by treatment of PEG-DAO and d-Ala was mediated by the function of MPO. Consistent with our hypothesis, 4-ABH, an MPO inhibitor, suppressed the bactericidal activity of the neutrophils, even in the treatment of PEG-DAO plus d-Ala (Figure 3).
As a more important result, we found that PEG-DAO plus d-Ala treatment restored the bactericidal activity of neutrophils from a CGD patient. Although the CGD neutrophils showed decreased bactericidal activity, in great contrast to healthy neutrophils (Figure 4), treatment with PEG-DAO plus d-Ala restored the bactericidal activity of these CGD neutrophils (Figure 4).
Our results thus demonstrated that H2O2 supplementation via PEG-DAO plus d-Ala would protect against bacterial infection. In our experiments, we used porcine DAO to prepare PEG-DAO. For porcine DAO, the Kcat and Km values for d-proline are 43.3 s−1 and 2 mmol/L, respectively, whereas the corresponding values for d-Ala are 6.4 s−1 and 3.1 mmol/L.15 Thus, using d-proline may be preferable to using d-Ala to treat CGD neutrophils, although this issue requires additional investigation. Furthermore, H2O2 can enter the cytosol of neutrophils and be converted to hypochlorous acid by MPO. These observations suggest that H2O2 supplementation via enzymatic action may become a plausible approach for treatment of patients with CGD. Less useful therapeutic strategies for CGD exist as yet, despite the great advances in the development of antimicrobial agents. The previously reported pharmacokinetics of PEG-DAO indicated an effective targeting ability to solid tumors as a result of the EPR effect. Bacterial components such as bacterial proteases and endotoxin facilitate the vascular permeability, and thus leakage of blood components such as albumin from circulating blood. Consequently, these macromolecules will accumulate at the infected or inflamed site.16,17 Although the accumulation property of macromolecular proteins at the tumor and inflamed tissue is similar, accumulation of PEG-DAO at the inflamed tissue in the CGD mouse model of human patients is yet to be determined. PEG-DAO pharmacokinetics may be similarly beneficial for targeting to the inflamed granuloma tissue in patients with CGD, although more studies are needed to confirm this possibility.
Author contributions: All authors participated in the design, interpretation of the studies, analysis of the data and review of the manuscript. HNa conducted the experiments and wrote the manuscript with HM; TM and HNu supplied the critical sample; and JF and HM discussed the content constructively for the experiments and revised the manuscript.
Footnotes
ACKNOWLEDGEMENTS
The use of blood samples from a patient with CGD was approved by the Institutional Review Board of University of Miyazaki, Japan. This work was supported by Grant-in-Aid for Scientific Research 21791016 from the Ministry of Education, Culture, Sports, Science and Technology of Japan.
JohnstonRBJr, KeeleBBJr, MisraHP, LehmeyerJE, WebbLS, BaehnerRL, RajagopalanKV. The role of superoxide anion generation in phagocytic bactericidal activity. Studies with normal and chronic granulomatous disease leukocytes. J Clin Invest1975;55:1357–72
3.
RepineJE, ClawsonCC. Quantitative measurement of the bactericidal capability of neutrophils from patients and carriers of chronic granulomatous disease. J Lab Clin Med1977;90:522–8
4.
BurtonK. The stabilization of D-amino acid oxidase by flavin-adenine dinucleotide, substrates and competitive inhibitors. Biochem J1951;48:458–67
5.
MosebachKO. [Mechanism of action of some factors affecting D-amino acid oxidase. III. Reactions and significance of nascent hydrogen peroxides.]. Hoppe Seylers Z Physiol Chem1957;309:206–18
6.
FangJ, SawaT, AkaikeT, MaedaH. Tumor-targeted delivery of polyethylene glycol-conjugated D-amino acid oxidase for antitumor therapy via enzymatic generation of hydrogen peroxide. Cancer Res2002;62:3138–43
7.
FangJ, DengD, NakamuraH, AkutaT, QinH, IyerAK, GreishK, MaedaH. Oxystress inducing antitumor therapeutics via tumor-targeted delivery of PEG-conjugated D-amino acid oxidase. Int J Cancer J Int Cancer2008;122:1135–44
8.
MatsumuraY, MaedaH. A new concept for macromolecular therapeutics in cancer chemotherapy: mechanism of tumoritropic accumulation of proteins and the antitumor agent smancs. Cancer Res1986;46:6387–92
9.
FangJ, NakamuraH, MaedaH. The EPR effect: unique features of tumor blood vessels for drug delivery, factors involved, and limitations and augmentation of the effect. Adv Drug Deliv Rev2011;63:136–51
10.
Gougerot-PocidaloMA, ElbimC, DangPM, El BennaJ. [Primary immune deficiencies in neutrophil functioning]. Presse Med2006;35:871–8
11.
ShenepJL, StokesDC, HughesWT. Lack of antibacterial activity after intravenous hydrogen peroxide infusion in experimental Escherichia coli sepsis. Infect Immun1985;48:607–10
12.
LebelCP, BondySC. Sensitive and rapid quantitation of oxygen reactive species formation in rat synaptosomes. Neurochem Int1990;17:435–40
13.
EllisJA, MayerSJ, JonesOT. The effect of the NADPH oxidase inhibitor diphenyleneiodonium on aerobic and anaerobic microbicidal activities of human neutrophils. Biochem J1988;251:887–91
14.
RosenH, CrowleyJR, HeineckeJW. Human neutrophils use the myeloperoxidase-hydrogen peroxide-chloride system to chlorinate but not nitrate bacterial proteins during phagocytosis. J Biol Chem2002;277:30463–8
15.
MollaG, SacchiS, BernasconiM, PiloneMS, FukuiK, PolegioniL. Characterization of human D-amino acid oxidase. FEBS Lett2006;580:2358–64
16.
XingJ, MoldobaevaN, BirukovaAA. Atrial natriuretic peptide protects against Staphylococcus aureus-induced lung injury and endothelial barrier dysfunction. J Appl Physiol2011;110:213–24
17.
KamataR, YamamotoT, MatsumotoK, MaedaH. A serratial protease causes vascular permeability reaction by activation of the Hageman factor-dependent pathway in guinea pigs. Infect Immun1985;48:747–53