
Abstract
Lectin-like oxidized low-density lipoprotein receptor-1 (LOX-1) has been identified as a major receptor for oxidatively modified low density lipoprotein (ox-LDL) in endothelial cells and smooth muscle cells (SMCs). MicroRNAs are small non-coding RNAs that regulate gene expression. Very few studies have reported regulation of LOX-1 expression by microRNAs in SMCs. The present study demonstrates that the microRNA hsa-let-7g can inhibit LOX-1 expression in a time- and dose-dependent fashion, and subsequently inhibit ox-LDL uptake in and proliferation of human aortic SMCs. We also show that hsa-let-7g can reduce SMC apoptosis by down-regulation of cytochrome c and Smac/Diablo and upregulation of Bcl-xL and Bcl-2 expression. Furthermore, we show that hsa-let-7g reduces ox-LDL-induced increase in the expression of NADPH oxidase (p22phox and p47phox subunits) and subsequent intracellular reactive oxygen species generation, phosphorylation of p44/42 mitogen-activated protein kinase and nuclear factor-kappaB p65 expression. These observations suggest that hsa-let-7g is a critical regulator of SMC apoptosis, and may be suitable for therapeutic intervention in disease states characterized by LOX-1 over-expression.
Introduction
Atherosclerosis is a chronic inflammatory disease characterized by lipid accumulation and cellular modifications within the arterial wall. 1 Oxidatively modified-low density lipoprotein (ox-LDL) plays a potent role in the genesis and progression of atherosclerosis. 2 Ox-LDL upregulates its own receptor, the lectin-like ox-LDL receptor-1 (LOX-1), a 40–50-kDa type II membrane protein. Among many receptors for ox-LDL, LOX-1 appears to be the dominant receptor on endothelial cells (ECs) and smooth muscle cells (SMCs). 3 LOX-1 exhibits a strong activity in binding, internalizing and proteolytically degrading ox-LDL. 4 A critical role of LOX-1 in atherogenesis has been shown by use of LDLR/LOX-1 double-knockout mice. 5
MicroRNAs (miRNAs) are endogenous, small, non-coding RNAs that negatively regulate gene expression by binding to their target messenger RNAs for degradation and/or translational repression. 6 MiRNAs are critical modulators for vascular functions, e.g. cell differentiation, contraction, migration, proliferation and apoptosis. 7 Furthermore, altered expression of miRNAs has been associated with inflammation, oxidative stress and angiogenesis. 8,9
Let-7 family members were first observed in Canenorhabditis elegans and nine members of the let-7 family have been identified in humans. 10 Let-7 family expression is usually noted in tissues during embryonic stages and significantly increases towards maturity, which suggests that it plays a pivotal role in the developmental process. 11 The Let-7 family has been shown to inhibit cancer cell proliferation through repressing oncogenes, including RAS and HMGA2, via binding to the 3′UTR of their mRNAs, thus it has also been reported as a tumor suppressor family. 12 The Let-7 family may also play a key role in cell proliferation and inflammation, steps that are critical to the evolution of atherosclerosis. 13–15
Our focus was to study the effect of let-7 family member, hsa-let-7g. This miRNA was predicted to have a putative target site in the LOX-1 3′-untranslated region (UTR) by bioinformatic tools RNA22. 15 We hypothesized that if hsa-let-7g would inhibit LOX-1 expression, it might also inhibit proliferation, ox-LDL uptake and apoptosis of SMCs. The present study was designed to test this hypothesis. We used a hsa-let-7g mimic for let-7g overexpression and its inhibitor in SMCs, and studied LOX-1 expression and function.
Materials and methods
Cell culture and miRNA transfection
Human aortic SMCs, CRL-1999, were obtained from ATCC (Manassas, VA, USA). SMCs were maintained in F-12 K medium containing 0.05 mg/mL ascorbic acid, 0.01 mg/mL insulin, 0.01 mg/mL transferring, 10 ng/mL sodium selenite, 0.03 mg/mL endothelial cell growth supplement, fetal bovine serum to a final concentration of 10%, hydroxyethyl piperazineethanesulfonic acid to a final concentration of 10 mmol/L and N-Tris(hydroxymethyl)methyl-2-aminoethanesulfonic acid buffer to a final concentration of 10 mmol/L. The cells were incubated at 37°C in a humidified atmosphere with 5% CO2.
The human hsa-let-7g mimic and inhibitor were synthesized by Applied Biosystems (Carlsbad, CA, USA). SMCs were seeded onto six-well plate at a density of 2 × 105 cells/well the day before transfection. Cells were transfected with the hsa-let-7g mimic or inhibitor using Lipofectamine 2000® (Invitrogen Corp., Grand Island, NY, USA) according to the manufacturer's instructions. The medium was replaced six hours after transfection. Trypsin-ethylenediaminetetraacetic acid was obtained from Gibco (Invitrogen).
High thiobarbituric acid reactive ox-LDL (90 nmoles MDA/mg protein) was purchased from Biomedical Technologies Inc. (Stoughton, MA, USA).
Western blot
Primary and secondary antibodies were purchased from Abcam (San Francisco, CA, USA) and Santa Cruz Biotechnology (Santa Cruz, CA, USA). Protein was extracted from SMCs using RIPA (radioimmunoprecipitation assay) buffer (Thermo Fisher Scientific Inc., Rockford, IL, USA). Proteins were separated by 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis, and transferred to polyvinylidene fluoride membrane (Millipore, Bedford, MA, USA). After blocking with phosphate-buffered saline (PBS) solution with Tween containing 5% bovine serum albumin or non-fat milk, membranes were sequentially incubated with primary antibodies (overnight at 4°C), washed with PBS solution with Tween (3 times, 10 min each), incubated with horseradish peroxidase-conjugated secondary antibodies for one hour at room temperature, washed, treated with the ECL Western blotting substrate (Promega, Madison, WI, USA) and imaged. The relative expression of proteins was evaluated in relation to β-actin.
Quantitative realtime polymerase chain reaction
Total RNA was extracted using TRIZOL reagent (Invitrogen). For hsa-let-7g detection, cDNA was synthesized by TaqMan MicroRNA assays (Applied Biosystems). To determine hsa-let-7g expression levels, U6 was used as internal control.
Dil-ox-LDL uptake
For visualization of specific ox-LDL uptake, triplicate cultures of SMCs in 24-well format were incubated with Dil-ox-LDL 5 μg/mL for two hours at 37°C. After incubation, cells were gently washed with PBS three times and imaged using fluorescent microscopy. 16
MTT (3-(4,5-dimethylthiazolyl-2)-2,5-diphenyltetrazolium bromide) analysis of cell proliferation
To measure the effects of hsa-let-7g on SMC proliferation, cells were seeded at 1 × 105 cells/well density on to a 24-well plate with the complete medium. After a 24-h incubation, cells were transfected with the hsa-let-7g mimic and inhibitor using Lipofectamine 2000®. The medium was removed after six hours of transfection and the cells washed two times with PBS. Eighteen hours later, cells were further treated with 20 μg/mL ox-LDL for another 24 h. SMC proliferation was performed by MTT (3-(4,5-dimethylthiazolyl-2)-2,5-diphenyltetrazolium bromide) Cell Proliferation Assay (ATCC) according to the manufacturer's protocol. Briefly, 10 μl MTT reagent was added followed by incubation for two to four hours until purple precipitate was visible, and then 100 μl detergent reagent was added and left at room temperature in the dark for two hours. Absorbance was recorded at 570 nm in a microplate reader.
Measurement of intracellular reactive oxygen species
Intracellular reactive oxygen species (ROS) was measured with the use of the fluorescent signal dihydroethidium (DHE), a cell-permeable indicator for ROS generation. SMCs cultured in a six-well plate were incubated with 10 μmol/L DHE in PBS for 30 min. The ROS-mediated fluorescence was observed under a Nikon, Eclipse E600 fluorescent microscope (Nikon Instruments Inc., Melville, NY, USA) with excitation set at 530 nm and emission set at 610 nm. DHE fluorescence intensity was measured using Image J 1.34 software (National Institutes of Health, Bethesda, MD, USA).
Analysis of apoptosis
Apoptosis was analyzed by Caspase Assay Detection Kit (ImmunoChemistry Technologies, Bloomington, MN, USA) according to the manufacturer-supplied protocols. The caspase activities were assessed by flow cytometry (Becton Dickinson, Franklin Lakes, NJ, USA), and the results were analyzed with WinMDI29 software (Becton Dickinson).
Statistical analysis
Data from at least three sets of samples were used for statistical analysis. Results are shown as mean ± SD. Multiple means were compared using one-way analysis of variance. Paired Student's t-test was used to assess significant differences. A P value < 0.05 was considered significant.
Results
Ox-LDL induces LOX-1 expression
As expected, treatment of SMCs with ox-LDL (0–40 μg/mL) resulted in a concentration-dependent increase in LOX-1 expression (Figure 1a). Importantly, hsa-let-7g expression decreased with increasing ox-LDL concentration (Figure 1b).
(a) Oxidatively modified-low density lipoprotein (ox-LDL) increases lectin-like oxidized low-density lipoprotein receptor-1 (LOX-1) protein in a concentration-dependent manner in human aortic smooth muscle cells; (b) ox-LDL decreases hsa-let-7g expression; (c) hsa-let-7g mimic decreases LOX-1 protein in a concentration-dependent manner; (d) while the hsa-let-7g inhibitor increases LOX-1 protein. The expression levels of LOX-1 and hsa-let-7g were measured by Western blot and quantitative realtime polymerase chain reaction. Bar graphs represent data in mean ± SD based on three–five experiments, *P < 0.05. (A color version of this figure is available in the online journal)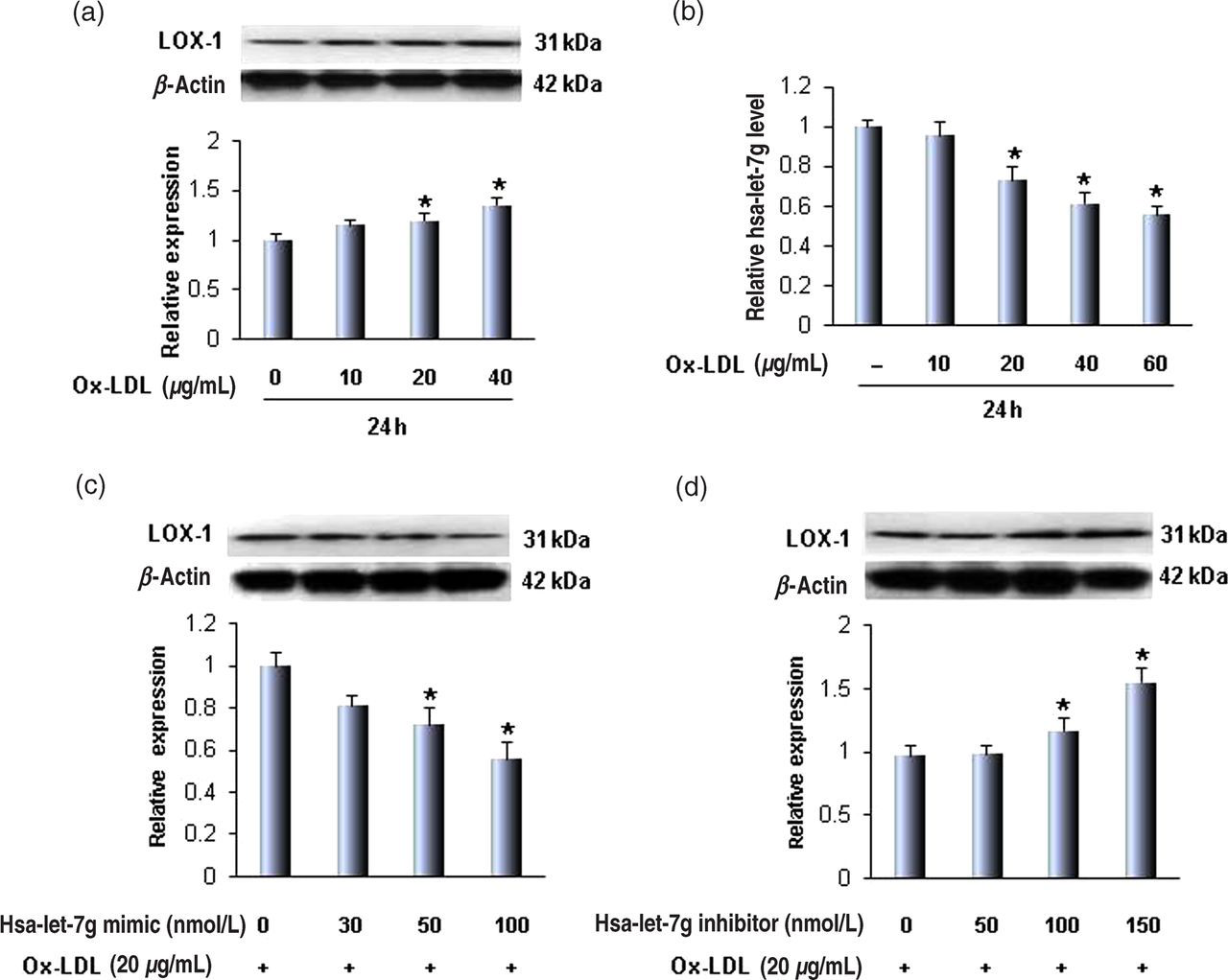
Hsa-let-7g mimic inhibits and its inhibitor enhances LOX-1 expression
In order to examine if hsa-let-7g regulates ox-LDL-mediated LOX-1 expression, different concentrations of a hsa-let-7g mimic and inhibitor were transfected into SMCs, and cells treated with ox-LDL. We observed that the hsa-let-7g mimic induced a decrease in LOX-1 expression in a concentration-dependent manner (Figure 1c). The hsa-let-7g inhibitor had the opposite effect on LOX-1 expression compared with the hsa-let-7g mimic (Figure 1d). These observations suggest a negative feedback regulation between LOX-1 and hsa-let-7g.
Based on the data in these first set of experiments, we used the hsa-let-7g mimic and hsa-let-7g inhibitor in 50 and 100 nM concentrations, respectively, in subsequent experiments.
Hsa-let-7g regulates ox-LDL uptake and proliferation in SMCs
Next, we examined if hsa-let-7g would influence ox-LDL uptake by SMCs. As shown in Figure 2a, ox-LDL enhanced Dil-ox-LDL uptake, and the hsa-let-7g mimic blocked it while hsa-let-7g inhibitor increased it. These changes paralleled the changes in LOX-1 expression (Figures 1c and d).
Hsa-let-7g regulates oxidatively modified-low density lipoprotein (ox-LDL) uptake and smooth muscle cell (SMC) proliferation. (a) Representative images of Dil-ox-LDL uptake by SMCs. (b) SMC proliferation. (c) Hsa-let-7g regulates ox-LDL-induced apoptosis. The apoptosis rate was measured by flow cytometry using the Caspase Assay Detection Kit. Left panels are representative experiments and right panels are values in mean ± SD based on three independent experiments. *P < 0.05. (A color version of this figure is available in the online journal)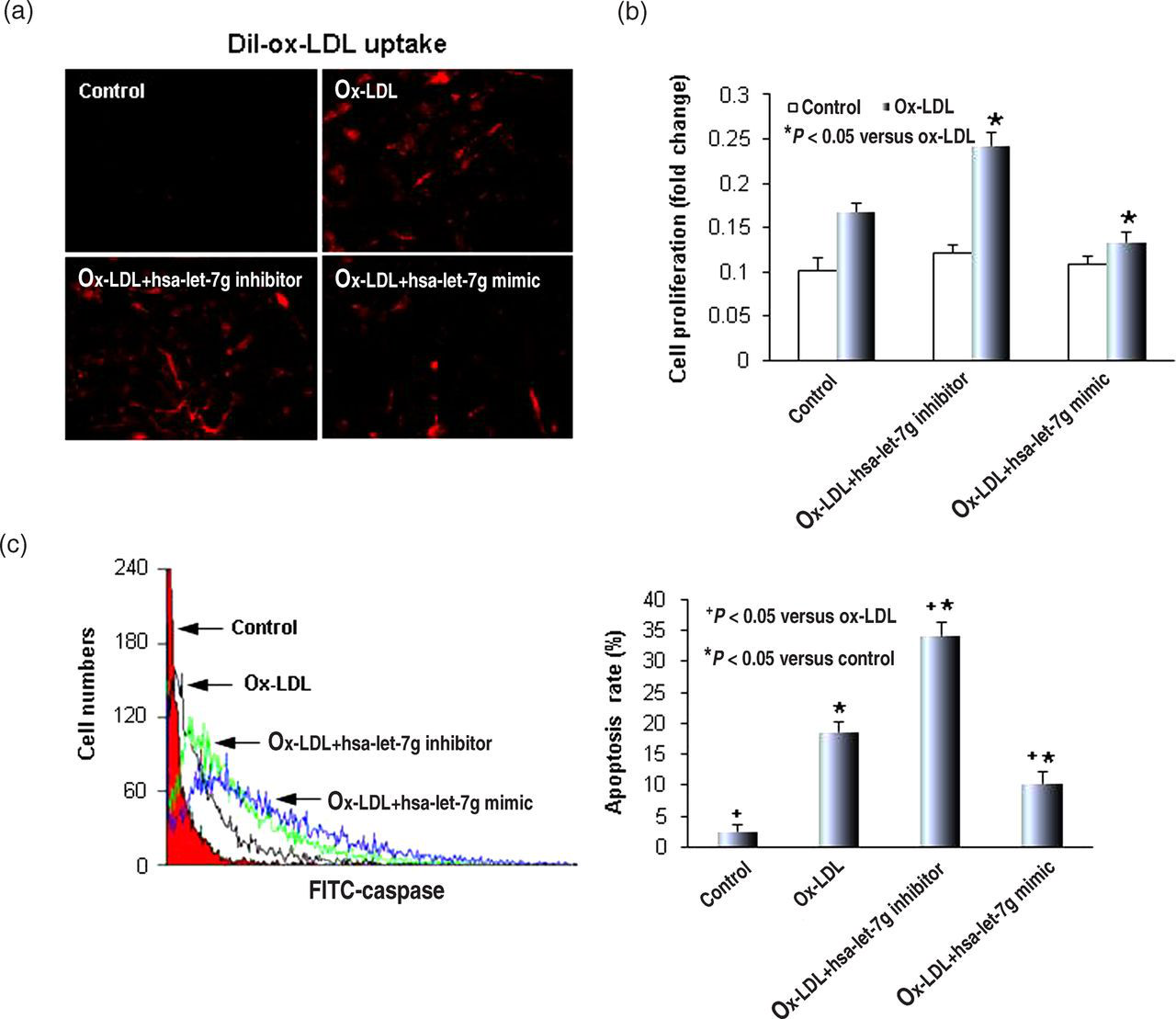
In order to examine the effect of hsa-let-7g on ox-LDL-mediated SMC proliferation, the hsa-let-7g mimic and hsa-let-7g inhibitor were transfected into SMCs for 24 h, and then the cells were treated with 20 μg/mL ox-LDL for another 24 h. Consistent with the data on LOX-1 expression and Dil-ox-LDL uptake, the hsa-let-7g inhibitor induced cell proliferation, while the hsa-let-7g mimic inhibited cell proliferation (Figure 2b). In the control group of cells, not treated with ox-LDL, neither the hsa-let-7g inhibitor nor the mimic had any effect on cell proliferation (data not shown).
Hsa-let-7g regulates apoptosis
Previous studies have shown that ox-LDL induces apoptosis in SMCs via LOX-1. 4 To examine if hsa-let-7g influences ox-LDL-induced apoptosis, SMCs were transfected with the hsa-let-7g mimic or inhibitor and then treated with 20 μg/mL ox-LDL for another 24 h.
We analyzed SMC apoptosis by caspase-3 assay (flow cytometry). As shown in Figure 2c, the apoptosis rate increased in cells treated with ox-LDL. Transfection with the hsa-let-7g mimic significantly attenuated ox-LDL-induced apoptosis of SMCs. In contrast, knockdown of hsa-let-7g was found to significantly enhance the rate of apoptosis.
It is well known that apoptosis is regulated by various proapoptosis-related proteins, such as cytochrome c, Smac/Diablo and antiapoptosis-related proteins Bcl-xL and Bcl-2.
17–19
Therefore, we examined the influence of the hsa-let-7g mimic and inhibitor on the expression of these proteins. As shown in Figures 3a and b, the hsa-let-7g mimic decreased cytochrome c expression in a concentration-dependent fashion. In contrast to the effect of the mimic, the hsa-let-7g inhibitor increased cytochrome c expression. Similar to the effect on cytochrome c expression, hsa-let-7g was noted to regulate Smac/Diablo expression (Figures 3c and d). In keeping with the data on cytochrome c and Smac/Diablo, hsa-let-7g mimic enhanced Bcl-xL and Bcl-2 expression, while its inhibitor decreased Bcl-xL (Figures 3e and f) and Bcl-2 (Figures 3g and h) in a concentration-dependent fashion.
Hsa-let-7g regulates cytochrome c, Smac/Diablo, Bcl-xL and Bcl-2 expression. (a, c) The hsa-let-7g mimic downregulated cytochrome c and Smac/Diablo. (b, d) In contrast, the hsa-let-7g inhibitor upregulated cytochrome c and Smac/Diablo expression. (e, g) The hsa-let-7g mimic also increased anti-apoptosis proteins Bcl-xL and Bcl-2 in a concentration-dependent fashion and (f, h) the hsa-let-7g inhibitor decreased Bcl-xL and Bcl-2. Bar graphs represent data in mean ± SD, based on three independent experiments. *P < 0.05; **P < 0.01. (A color version of this figure is available in the online journal)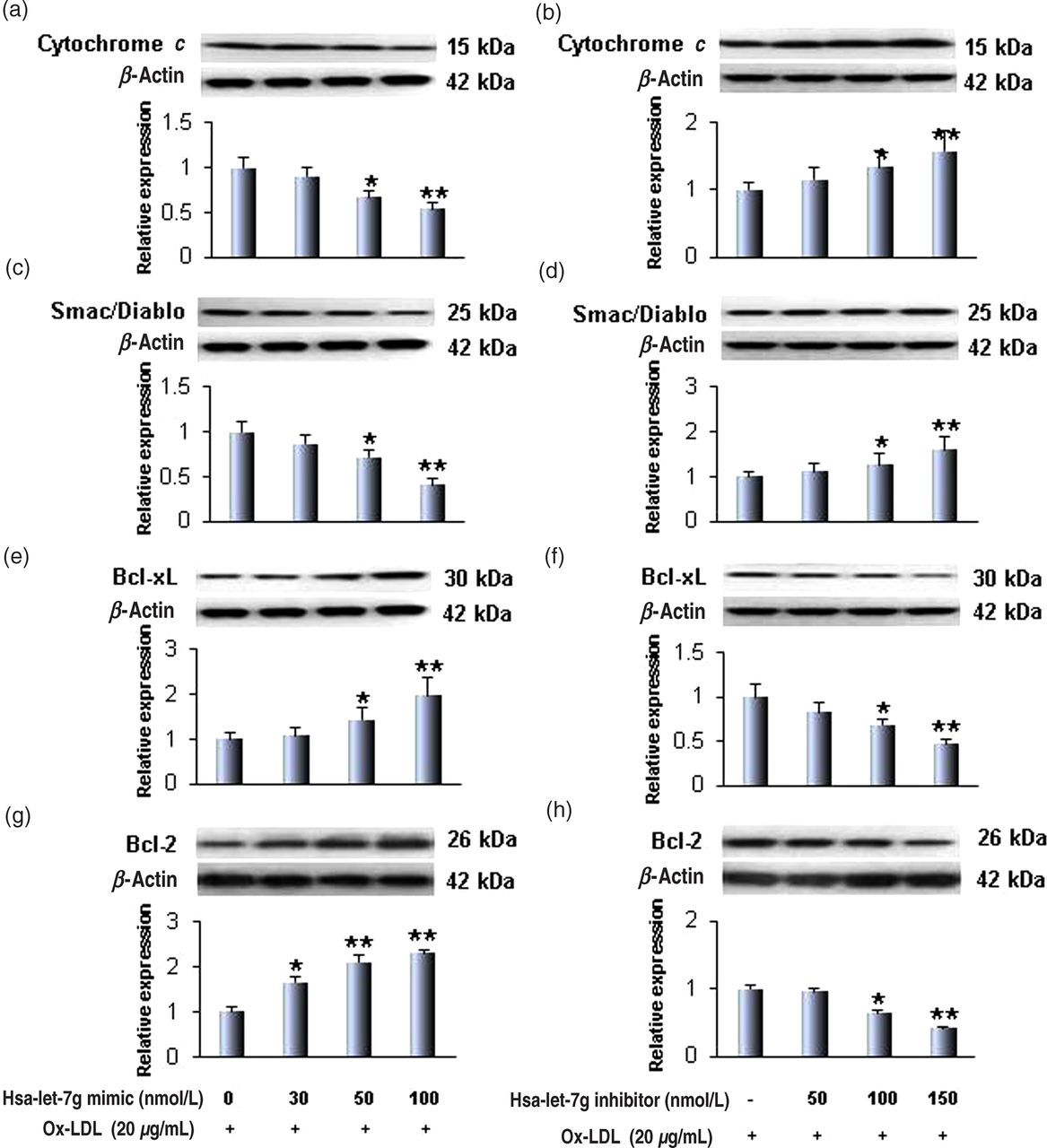
Hsa-let-7g, LOX-1 and intracellular signaling
The formation of ROS can be stimulated by LOX-1 activation, and NADPH oxidase activation is a major source of ROS in ECs and SMCs.
3
Therefore, NADPH oxidase expression and intracellular ROS generation were measured in SMCs transfected with the hsa-let-7g mimic and its inhibitor in the presence of ox-LDL. Consistent with the data on LOX-1 expression (Figure 1a), the expression of NADPH oxidase (p22phox and p47phox subunits) was observed to be clearly regulated by hsa-let-7g (Figures 4a and b). Furthermore, the hsa-let-7g mimic markedly reduced ox-LDL-mediated DHE florescence, reflecting reduction in intracellular ROS generation, while the hsa-let-7g inhibitor enhanced DHE florescence in SMCs (Figure 4c). These data suggest that hsa-let-7g influences ox-LDL-mediated ROS generation by regulating NADPH oxidase expression.
Hsa-let-7g regulates NADPH oxidase (p22phox and p47phox subunits) and reactive oxygen species (ROS) generation in smooth muscle cells. (a, b) The hsa-let-7g mimic downregulated, while the hsa-let-7g inhibitor upregulated p22phox and p47phox. Bar graphs represent data in mean ± SD from three independent experiments. *P < 0.05 and **P < 0.01. (c) Representative images of ROS generation, with quantitation of ROS generation data on the right. (A color version of this figure is available in the online journal)
It has been suggested that LOX-1 expression in vascular cells initiates a cascade of redox-sensitive signaling events including activation of the mitogen-activated protein kinase (MAPK) pathway.
20
To determine if hsa-let-7g participates in the regulation of the MAPK pathway, the expression of p44/42 components of MAPKs was measured by Western blot. As shown in Figure 5a, the expression of phospho-p44/42 MAPK was significantly suppressed by the hsa-let-7g mimic; on the other hand, the hsa-let-7g inhibitor induced phosphop42/44 MAPK expression (Figure 5b). Effects of the mimic and inhibitor were concentration-dependent. It is of note that the unphosphorylated form of MAPK was not affected by the hsa-let-7g mimic or its inhibitor (data not shown).
Hsa-let-7g regulates MAPK p44/42 and NF-κB p65 in smooth muscle cells. (a, c) The hsa-let-7g mimic downregulated p44/42 and NF-κB p65 activation. (c, d) The hsa-let-7g inhibitor upregulated MAPK p44/42 and NF-κB activity activation. Bar graphs represent data in mean ± SD, based on three independent experiments. *P < 0.05; **P < 0.01. (A color version of this figure is available in the online journal)
Next, we measured the expression of redox-sensitive transcription factor nuclear factor-kappaB (NF-κB) p65 with hsa-let-7g transfection in the presence of ox-LDL. As shown in Figure 5c, the expression of phospho-NF-κB p65 induced by ox-LDL was inhibited by the hsa-let-7g mimic, and the hsa-let-7g inhibitor enhanced phospho-NF-κB p65 expression (Figure 5d). These effects of the hsa-let-7g mimic and inhibitor were concentration-dependent. Similar to p44/42 MAPK, unphosphorylated NF-κB p65 was not affected by the hsa-let-7g mimic or its inhibitor (data not shown).
Discussion
Apoptosis (programed cell death) and proliferation of vascular SMCs has been identified as an important process in a variety of human vascular disease states, including atherosclerosis, arterial injury and restenosis after angioplasty. 21 All these pathological states are associated with oxidant stress and LOX-1 over-expression. 3–5 In the present study, we identified that the miRNA hsa-let-7g is a critical regulator of LOX-1 expression and apoptosis in human aortic SMCs.
Chen et al. 22 have recently demonstrated that ox-LDL can reduce let-7g expression, and showed that there is a let-7g binding site on the 3′-untranslated region of LOX-1 mRNA. We significantly expand on these observation by showing that the hsa-let-7g mimic inhibited ox-LDL-mediated LOX-1 expression, which resulted in a decrease in ox-LDL uptake in SMCs as well as proliferation of these cells. We also show that hsa-let-7g influences ROS generation by regulating NADPH oxidase expression. Activation of the p44/42 MAPKs-NF-κB pathway which is downstream of ROS generation was also inhibited by hsa-let-7g. Lastly, we observed that this miRNA inhibited SMC apoptosis by regulating apoptosis-related proteins. In essence, the present findings confirm that hsa-let-7g is a potent regulator of LOX-1 and resultant apoptosis on SMCs.
MiRNAs are highly conserved, non-coding RNA molecules that generally bind to the target sequence localized in the 3′-untranslated region of their target miRNAs and regulate inflammation, oxidative stress, apoptosis and angiogenesis in atherosclerotic plaques. 12 LOX-1 is a major receptor for ox-LDL expressed in vascular ECs, macrophages and vascular SMCs 3,5,16,18 and serves as a potent pro-oxidant and proinflammatory signal. The regulation of LOX-1 expression is also important for many physiological processes, such as vascular cell proliferation, apoptosis and vascular remodeling. 17,22,23
The Let-7 family has been shown to play a pivotal role in cell proliferation, migration and certain cancers. 12–15 The present study shows that hsa-let-7g modulates internalization of ox-LDL in vascular SMCs via LOX-1 (Figure 2a). Ox-LDL can also induce SMC proliferation at low concentrations, 24 which was also blocked by the hsa-let-7g mimic and exacerbated by the hsa-let-7g inhibitor (Figure 2b). We also studied the development of apoptosis in SMCs. Using multiple markers of cell apoptosis (caspase-3 assay, cytochrome c, Smac/Diablo, Bcl-xL and Bcl-2), we showed that hsa-let-7g inhibited ox-LDL-mediated SMC apoptosis. The role of hsa-let-7g in the regulation of SMC apoptosis was verified with the use of its inhibitor.
An increasing body of evidence indicates that ROS, as secondary messengers, play key roles in many cellular events including cell proliferation, apoptosis, hypertrophy and inflammation. 3,25–27 The NADPH oxidase system is the most potent ROS-generating system in vascular cells, including SMCs. 28,29 The prototype phagocytic NADPH oxidase is composed of membrane-bound gp91phox and p22phox, as well as cytosolic subunits p47phox, p67phox and small GTPase Rac. In the present study, we found that hsa-let-7g inhibited p22phox and p47phox subunits of NADPH oxidase and subsequent generation of ROS (Figure 4). We did not study other components of NADPH oxidase, but changes in other subunits generally parallel the changes in p22phox and p47phox subunits. 3 The cytotoxic effect of ROS is thought to involve intracellular activation of MAPKs and NF-κB. 3,30,31 Our study showed that hsa-let-7g inhibits activation of both p42/44 MAPK and NF-κB p65. Not unexpectedly, the hsa-let-7g inhibitor had an opposite effect on NADPH-MAPK-NF-kB signaling and development of apoptosis. We focused on the p44/42 component of MAPK as this component relates to cell proliferation. 3
It has been reported that ox-LDL at higher concentrations (>60 μg/mL) induces cell apoptosis while it causes cellular proliferation at lower concentrations (<20 μg/mL) in bovine aortic SMCs.19 The present study showed that ox-LDL induces both cell proliferation and apoptosis at 20 μg/mL; these apparently contradictory phenomena can be explained as follows: though ox-LDL at this concentration causes apoptosis in about 19% of cells (Figure 2c), the cell proliferation is the dominant feature (Figure 2b), which is consistent with our previous data. 17
In summary, the present study provides evidence that the miRNA hsa-let-7g inhibits LOX-1 expression/activation, proliferation and development of apoptosis in vascular cells. Since LOX-1 activation is involved in a number of disease states, such as atherosclerosis (that are characterized by SMC proliferation and apoptosis), it is possible that hsa-let-7g might become the target for further investigation in these disease states.
Footnotes
ACKNOWLEDGEMENT
This study was supported in part by funds from the Department of Veterans Affairs.