
Abstract
Leptin is stated to be an important mediator between obesity and cardiovascular disease. However, whether leptin location in the aorta is dependent on diet and its atherogenic character is still unknown. This study examined the relationship between a high-fat diet with or without cholesterol and the expression of leptin in aortic layers. Forty male rats were fed a high-fat diet with fish or grape seed oil as a dietary fat source, with or without cholesterol, to enhance the atherogenic character of their diet. The experiments lasted for three and six weeks, respectively. Plasma lipid profile, plasma leptin concentration and leptin expression in the endothelium (E), myometrium (M) and adventitia (A) were examined. The length of feeding was a key factor which influenced both the lipid profile and leptin expression in the aorta. Leptin concentration positively correlated with body weight and plasma triglycerides only in the three-week experiment, which suggests that the physiological actions of leptin can be disturbed by prolonged consumption of a high-fat diet. Short-term intake of a high-fat diet with fish oil, increasing high density lipoprotein cholesterol (HDL) level and decreasing total cholesterol (TC)/HDL ratio, enhanced leptin expression in E in comparison to the group fed grape seed oil. However, in the group with the highest diet intake, leptin expression in each layer was lowest. Generally, leptin expression was most common in E; however, an extension of the period of feeding in groups fed a diet with grape seed oil with or without additional cholesterol increased leptin presence in M and A in comparison to the group fed fish oil. Significantly higher values of TC and HDL in the same groups may suggest that leptin changes in the aorta and the atherogenic impact of grape seed oil can be connected when the consumption of a high-fat diet is excessive.
Introduction
Atherosclerosis is a chronic inflammatory disease which is closely connected to the metabolic syndrome. Lifestyle patterns, such as inappropriate dietary habits and physical inactivity, promote the development of excessive body mass, which with hypertension, hypertriglyceridemia and insulin resistance is a risk factor for cardiovascular events. 1 However, mechanisms and mediators connecting obesity and cardiovascular dysfunction are still poorly understood. Adipose deposition as a source of cytokines and adipokines has been suggested to impose an inflammatory reaction in the vascular wall. 2 One of these adipokines is leptin, which is stated to be one of the most important mediators between obesity and cardiovascular disease (CVD).3,4
Leptin is a 167-amino acid peptide hormone produced mainly by adipocytes (white adipose tissue). Initial research only connected leptin's effects with food intake and energy balance. 5 However, it is currently being investigated as an important cardiovascular factor; as its functional receptors, mostly the long isoform OB-Rb, were found in vascular smooth muscle cells 6 and endothelial cells 7 as well as in atherosclerotic lesions. 8 As was reviewed by several authors,9,10 leptin accelerates the pathogenesis of atherosclerosis by several mechanisms, such as enhancing pro-inflammatory cytokine production in macrophages and monocytes (interleukin [IL]-6, -12, -18, and tumor necrosis factor – TNF-α), increasing oxidative stress in endothelium cells, promoting vascular smooth muscle cell migration and proliferation, increasing obesity-associated hypertension, and stimulating platelet aggregation.
As leptin's main role is connected with food intake and satiety, its plasma concentration should be dependent on diet composition. One of the most important diet components responsible for changes in this adipokine's balance is dietary fat, both its quality and quantity. It is well documented that a high-fat diet increases leptin plasma concentration, 11 but the consequences of fatty acid modifications in the diet on this adipokine's level are still uncertain. 12 However, it is generally known that fatty acid composition in the diet plays an important role in the prevention and treatment of cardiovascular events. 13 Inflammatory disease, such as atherosclerosis, can be either intensified or moderated by dietary components, which would be acting as pre- or pro-inflammatory molecules such as polyunsatu-rated fatty acids (PUFA). The pro-inflammatory eicosanoids such as prostaglandin E2 (PGE2) and leukotriene B4 (LTB4) are derived from n-6 fatty acid – arachidonic acid (AA – C20:4) by action of cyclooxygenase and 5-lipoxygenase, respectively. Also, the same enzymes can convert n-3 fatty acids such as eicosapentaenoic acid (EPA – C22:5), intensifying production of 5 series leucotriens (LTB5) and series 3 eicosanoides (PGE3) which demonstrate little inflammatory potency. Moreover, a novel family of EPA- and DHA (docosahexaenoic acid, C22:6)-derived lipid mediators called resolvins (E- and D-series) impose anti-inflammatory activity, reducing acute inflammation states. Additionally, inclusion of n-3 fatty acids in the diet can suppress the production of pro-inflammatory cytokines such as TNF-α and IL-1 β. However, in the modern Western diet there is an abundance of n-6 PUFA, especially linoleic acid (LA, C18:2), from soy, corn, sunflower or grape seed oil. LA elongated to 20-carbon acids is a main source of AA. Consumption of n-3 fatty acids is much lower because EPA and DHA appears only in fish, and their precursor α-linolenic acid (ALA, C18:3) is only in leafy green vegetables and flaxseed oil.
Other positive actions of n-3 fatty acids, such as normalization of lipidemic profile, reduction of blood coagulability and improvement of endothelium function by enhancing activity of NO synthetase, demonstrate them as factors which probably reduce cardiovascular events and so are recommended for dietary prevention.14–17
Knowing what was mentioned above, we have formulated a hypothesis that leptin localization profile in aortic wall layers (endothelium, myometrium and adventitia) will be altered by dietary fatty acid composition when a diet is high-fat, with or without supplementation of cholesterol. Furthermore, this change will be dependent on the duration of the experiment.
Materials and methods
Animals, diets and experimental design
The experiment was conducted on 48 male adult Wistar rats with an initial body weight of 272 ± 20 g. Animals were kept individually in polypropylene cages in stable conditions (temperature 22°C; humidity 50%; 12:12 light:dark cycle). They were given free access to food and water.
During the first week of adaptation, animals were fed a standard rodents’ feed Labofeed H (Andrzej Morawski Feed Production Plant, Kcynia, Poland). Subsequently animals were divided into two main groups with different periods of feeding – three and six weeks. Within these groups, rats were split into four subgroups with various dietary fat sources with or without an extra 3% addition of cholesterol. All experimental diets were high-fat (20% w/w) and semi-synthetic, with either fish oil (rich in n-3 PUFAs) or grape seed oil (rich in PUFA n-6) as a fat source. The characteristic of diets and use of nomenclature are presented in Table 1 while the composition of diets is shown in Table 2. According to diet composition caloric content of the basic diets was 463.2 kcal/100 g and the basic diets with cholesterol – 451.2 kcal/100 g. The analysis of the fatty acid profile in used fish oil and grape seed oil was performed by the Industrial Chemistry Research Institute in Warsaw, and the results are shown in Table 3.
Diets characterization and used symbols
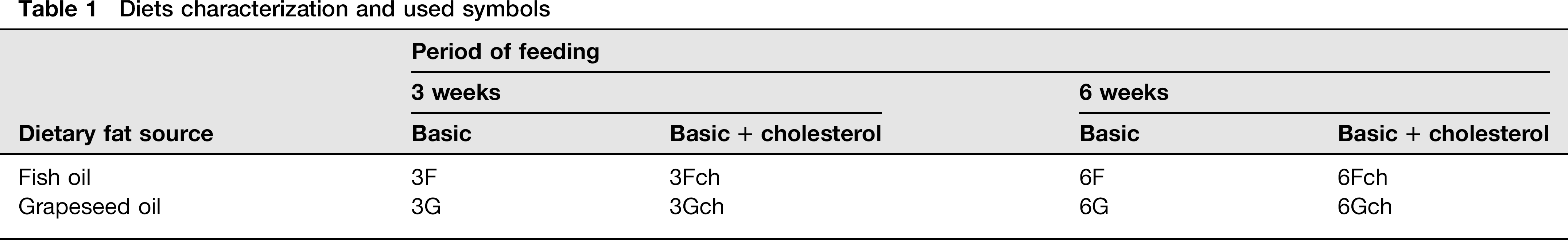
Composition of diets (g/100 g of diet)
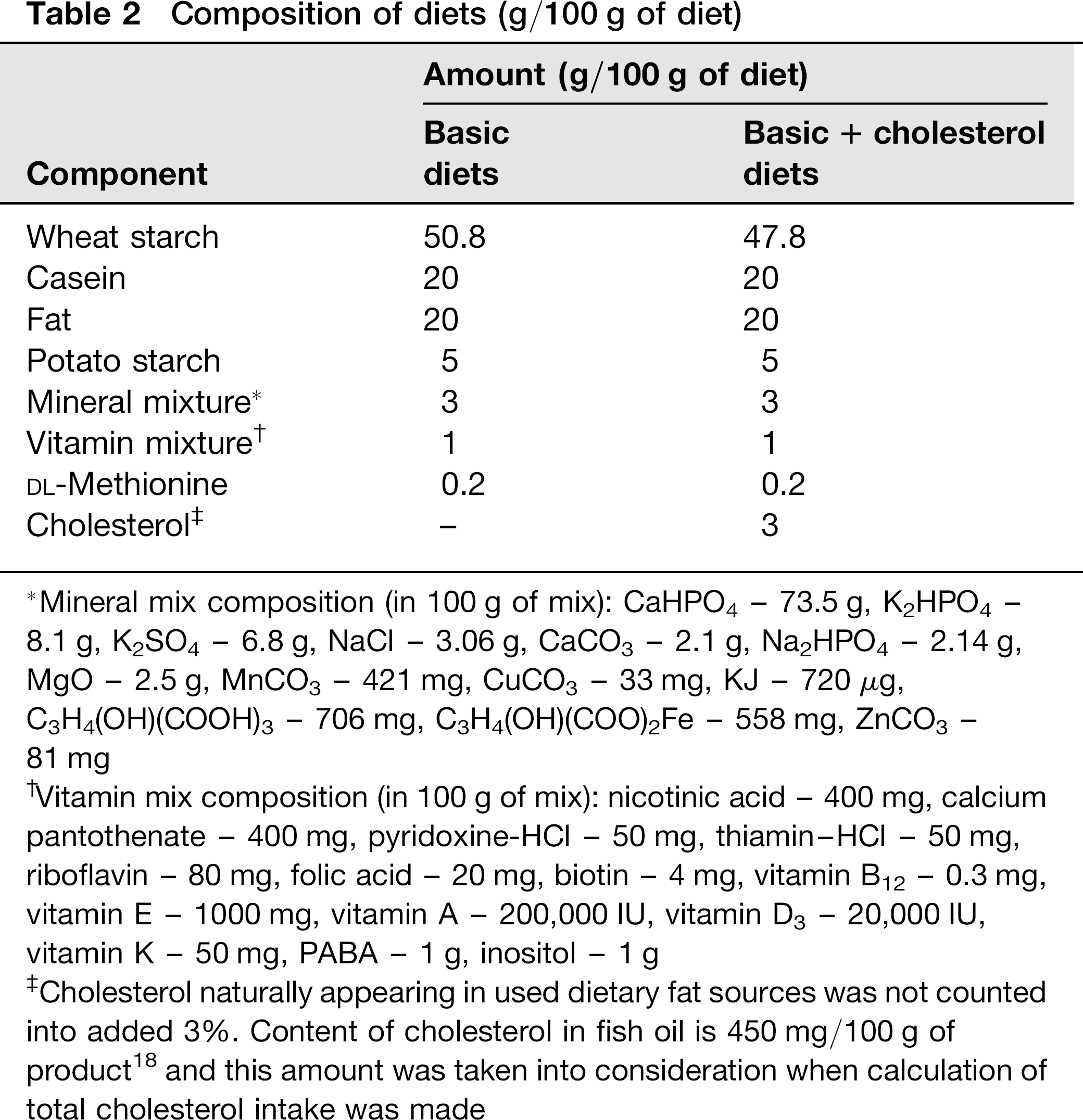
Mineral mix composition (in 100 g of mix): CaHPO4 - 73.5 g, K2HPO4 -8.1 g, K2SO4 - 6.8 g, NaCl - 3.06 g, CaCO3 - 2.1 g, Na2HPO4 - 2.14 g, MgO - 2.5 g, MnCO3 - 421 mg, CuCO3 - 33 mg, KJ - 720 μg, C3H4(OH)(COOH)3 - 706 mg, C3H4(OH)(COO)2Fe - 558 mg, ZnCO3 -81 mg
Vitamin mix composition (in 100 g of mix): nicotinic acid - 400 mg, calcium pantothenate - 400 mg, pyridoxine-HCl - 50 mg, thiamin-HCl - 50 mg, riboflavin - 80 mg, folic acid - 20 mg, biotin - 4 mg, vitamin B12 - 0.3 mg, vitamin E - 1000 mg, vitamin A - 200,000 IU, vitamin D3 - 20,000 IU, vitamin K - 50 mg, PABA - 1 g, inositol - 1 g
Cholesterol naturally appearing in used dietary fat sources was not counted into added 3%. Content of cholesterol in fish oil is 450 mg/100 g of product 18 and this amount was taken into consideration when calculation of total cholesterol intake was made
Content of main fatty acids in dietary fat sources (g/100 g of product)

Fatty acids profile was determined with AOCS Ce 2-66 method by Industrial Chemistry Research Institute in Warsaw; SFA, saturated fatty acids; MUFA, monounsaturated fatty acids; PUFA, polyunsaturated fatty acids; C16:0, palmitic acid; C18:0, stearic acid; C18:1, oleic acid; C18:2, linoleic acid; C18:3, α-linolenic acid; C20:4, arachidonic acid; C20:5, eicosapentaenoic acid; C22:6, docosahexaenoic acid
Rats’ body weight was monitored once each week and the food intake was counted by daily measurement of uncon-sumed diet through the whole experiment. Body weight gain was counted as: (final body weight - initial body weight)/number of days (21 or 42, respectively to the period of feeding).
After the proper feeding period, rats were anesthetized via the peritoneum by Thiopental and bled by cardiac puncture. Blood was centrifuged (20 min, 3000 rpm, 4°C) and the collected plasma was stored at -20°C until further analysis. The aorta was removed and fixed in buffered 10% formalin for 24 h.
The study was approved by the Third Local Animal Care and Use Committee in Warsaw (Poland).
Plasma lipid profile colorimetric assays
Concentrations of plasma total cholesterol (TC), triglycerides (TG) and high-density lipoprotein cholesterol (HDL) were measured using enzymatic-colorimetric methods. Kits containing ready to use liquid reagents were purchased from PTH Hydrex (Warsaw, Poland). The analyses were held according to kits manuals. Concentrations were expressed as milligrams per deciliter of plasma.
Leptin radioimmunological assay
Plasma leptin concentration was measured using Rat Leptin RIA Kit (Cat. # RL-83K, LINCO Research, St. Charles, MO, USA). The intra- and inter-assay precision was 2.4% and 4.8%, respectively. The sensitivity of the test was 0.5 ng/mL. The assay was conducted according to the kit manual and the leptin concentration was expressed as nano-grams per milliliter of plasma.
Leptin immunohistochemical assay in aorta layers
Aorta specimens were analyzed immunohistochemically using the rabbit polyclonal antibody Anti-OB (AB-20) from Santa Cruz Biotechnology Inc. (Santa Cruz, CA, USA, cat. no. sc-842). Three-micrometer sections of formalin-fixed, paraffin-embedded aorta specimens were deparaffinized with xylene in two changes of 10 min each, and rehydrated using graded alcohols (99.8%, 96% and 70%) twice for five minutes each. Antigens were retrieved by heating the sections in 0.01 mol/L citrate buffer (pH 6.0) for five minutes at 700 W in a microwave oven four times. Afterwards, specimens were cooled, rinsed with distilled water, kept in phosphate-buffered saline (pH 7.4) for five minutes and then incubated for 15 min in 10% bovine serum albumin to block non-specific tissue reactivity.
The sections were then incubated with the primary antibody Anti-OB (AB-20) in a 1:1000 dilution overnight at 4°C. After incubation with the primary antibody, the slides were incubated with biotinylated secondary goat anti-rabbit IgG (1:1500, cat. no. 830, Immunotech, Marseille, France) for 30 min. Endogenous peroxidase activity was inactivated by a five-minute incubation with 3% H2O2, and the sections were then incubated with a peroxidase-conjugated streptavidin (1:500, cat. no. 309, Immunotech) for 45 min. The complex was then detected with 3.3'-diaminobenzidine (DAB) (Sigma-Aldrich, St. Louis, MO, USA) as a chromogen, whereby the sections were lightly counterstained with hematoxylin and mounted in DPX mounting medium (BD Biosciences, Bedford, MA, USA).
Normal rabbit IgG of the same concentration as the primary antibody was used as a negative control.
Leptin localization was identified in one 3-μm section per animal, separately in endothelium (E), myometrium (M) and adventitia (A).
Results were calculated according to Ball et al.
1
One hundred cells per each layer of aorta were assigned to an intensity category of 0 (absent), 1 (weak), 2 (moderate) or 3 (strong) (intensity of each reaction is presented in Figure 1). The percentage of cells in each intensity category was determined as N0, N1, N2 and N3, respectively. A weighted average (ID score) was then calculated as
Leptin expression in the layers of the wall of the aorta after three weeks (a) and six weeks (b) (microscope photo, × 400). 
The ID score therefore ranged from 0 (absent staining in all cells) to a maximum of 3 (100% cells having a staining intensity of 3).
Statistical analysis
Statistical analysis was made using STATISTICA v. 8.00 (StatSoft Polska Sp. z o.o., Cracow, Poland). Results of simple regression, two-way variance analysis ANOVA, followed by post hoc Fisher's test and Student's T-test were considered statistically significant at P ≤ 0.05. All data are showed as means ± standard deviation (SD).
Results
Body weight gain, food and energy intake
It was shown that final body weight gain (Table 4) depended on the period of feeding (ANOVA, P ≤ 0.0001) and it was higher after three weeks in all four groups (F, Fch, G and Gch). When rats were fed for three weeks, final body weight gain depended on fat type and cholesterol addition (ANOVA, P ≤ 0.004 and P ≤ 0.0001, respectively) and it was highest in group 3Fch. After six weeks of feeding no such association was observed.
Body weight gain (g/d) and intake of diet, total fat, SFA, MUFA, PUFA, n-3, n-6 (g/d/100 g body weight), cholesterol (mg/d/100 g body weight) and energy (kcal/d/100 g body weight)
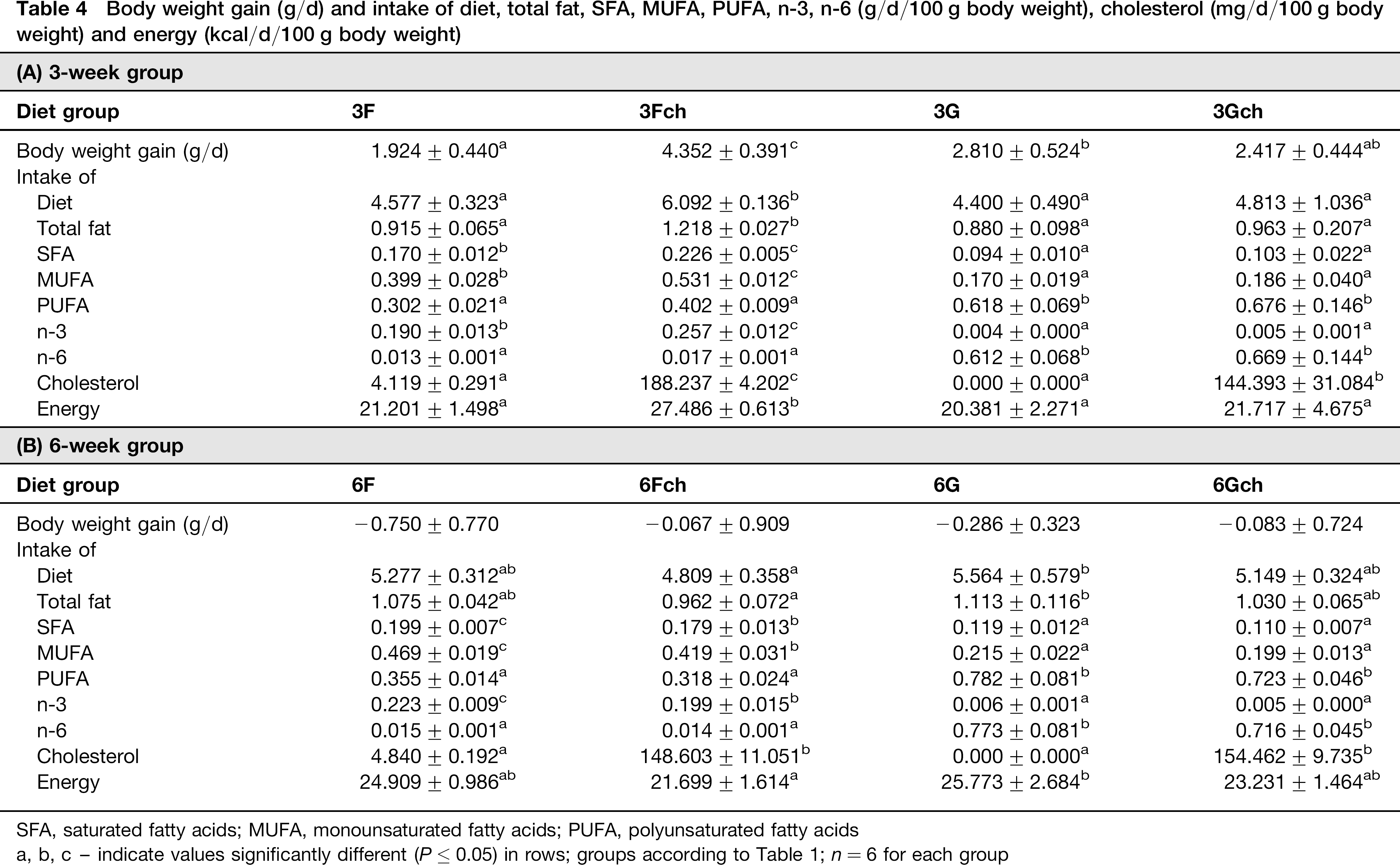
SFA, saturated fatty acids; MUFA, monounsaturated fatty acids; PUFA, polyunsaturated fatty acids
a, b, c - indicate values significantly different (P ≤ 0.05) in rows; groups according to Table 1; n = 6 for each group
Food consumption and energy intake was not dependent on the period of feeding, but after three weeks these parameters were dependent on cholesterol supplementation and fat type (ANOVA, ≤ 0.002 and P≤ 0.02, respectively for both parameters) and were highest in group 3Fch. After six weeks, animals fed a diet with no additional cholesterol consumed a higher amount of the diet (ANOVA, P ≤ 0.01).
Consumption of particular types of fatty acids depended on the dietary fat source used. As a result of the fatty acid composition, higher consumption of saturated fatty acids (SFA), monounsaturated fatty acids (MUFA) and PUFA n-3 was found in groups fed a diet with fish oil, and higher consumption of PUFA n-6 in groups fed a diet with grape seed oil (ANOVA for each fatty acids group P ≤ 0.0001).
Cholesterol intake was greater in groups fed fish oil than grape seed oil, because of naturally appearing cholesterol in this oil (ANOVA, P ≤ 0.001). Also, statistically important higher values of cholesterol intake were observed in groups with supplementation of this compound (ANOVA, P ≤ 0.0001).
Plasma lipid profile
The plasma lipid profile depended on the period of feeding, with all three parameters (TC, HDL, TG) lower after three weeks of feeding (ANOVA, P ≤ 0.0001 for all three parameters) (Table 5).
Plasma lipid profile: TC – total cholesterol (mg/dL), HDL – high-density lipoprotein cholesterol (mg/dL), TG – triglycerides (mg/dL) and TC/HDL ratio after (A) 3 weeks and (B) 6 weeks
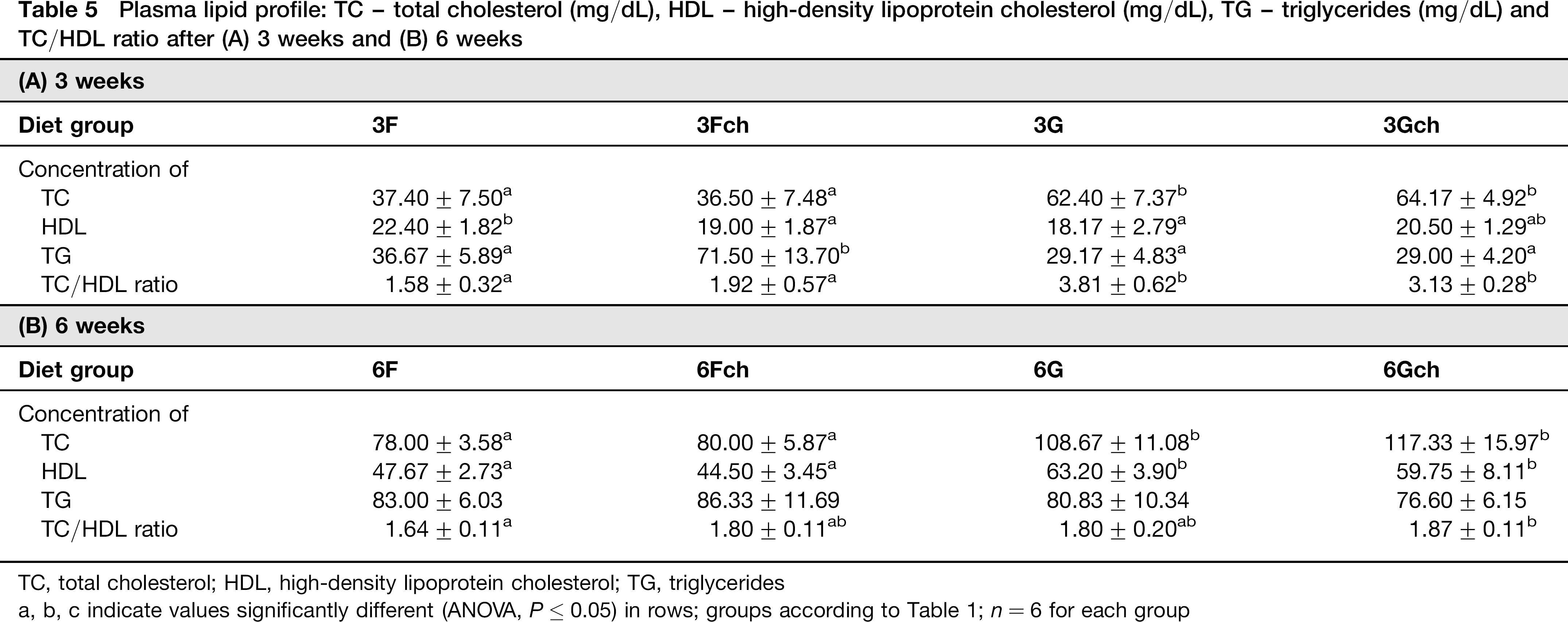
TC, total cholesterol; HDL, high-density lipoprotein cholesterol; TG, triglycerides
a, b, c indicate values significantly different (ANOVA, P < 0.05) in rows; groups according to Table 1; n = 6 for each group
TC was lower in groups fed diets with fish oil after both three and six weeks (ANOVA, P ≤ 0.0001 for both periods). However HDL cholesterol level was higher in groups fed a diet with grape seed oil only after six weeks (ANOVA, P ≤ 0.0001). Therefore, the ratio of TC to HDL was lower in groups fed diets with fish oil for three weeks (ANOVA, P ≤ 0.0005). Interestingly, the highest values of TC/HDL ratio were observed in groups fed grape seed oil for three weeks; after a longer period of feeding these values decreased and did not differ from groups fed diet 3Fch. TG level was dependent on fat type and cholesterol supplementation, but only after three weeks. Higher values were observed in animals from the group with fish oil in their diets and when there was cholesterol supplementation. This showed that TG level was highest in group 3Fch. After six weeks no such influence was observed.
Plasma leptin concentration
It was shown that the plasma leptin level was not dependent on the period of feeding. However, analysis with the Student's T-test showed that leptin concentration was higher when rats were fed diets 6F (3F versus 6F, P ≤ 0.0001) and 6Gch (3Gch versus 6Gch P ≤ 0.0001), and were lower in groups 6Fch (3Fch versus 6Fch P ≤ 0.0002) and 6G (3G versus 6G P ≤ 0.0001) (Figure 2).
Plasma leptin concentration (ng/mL). a and b indicate values significantly different (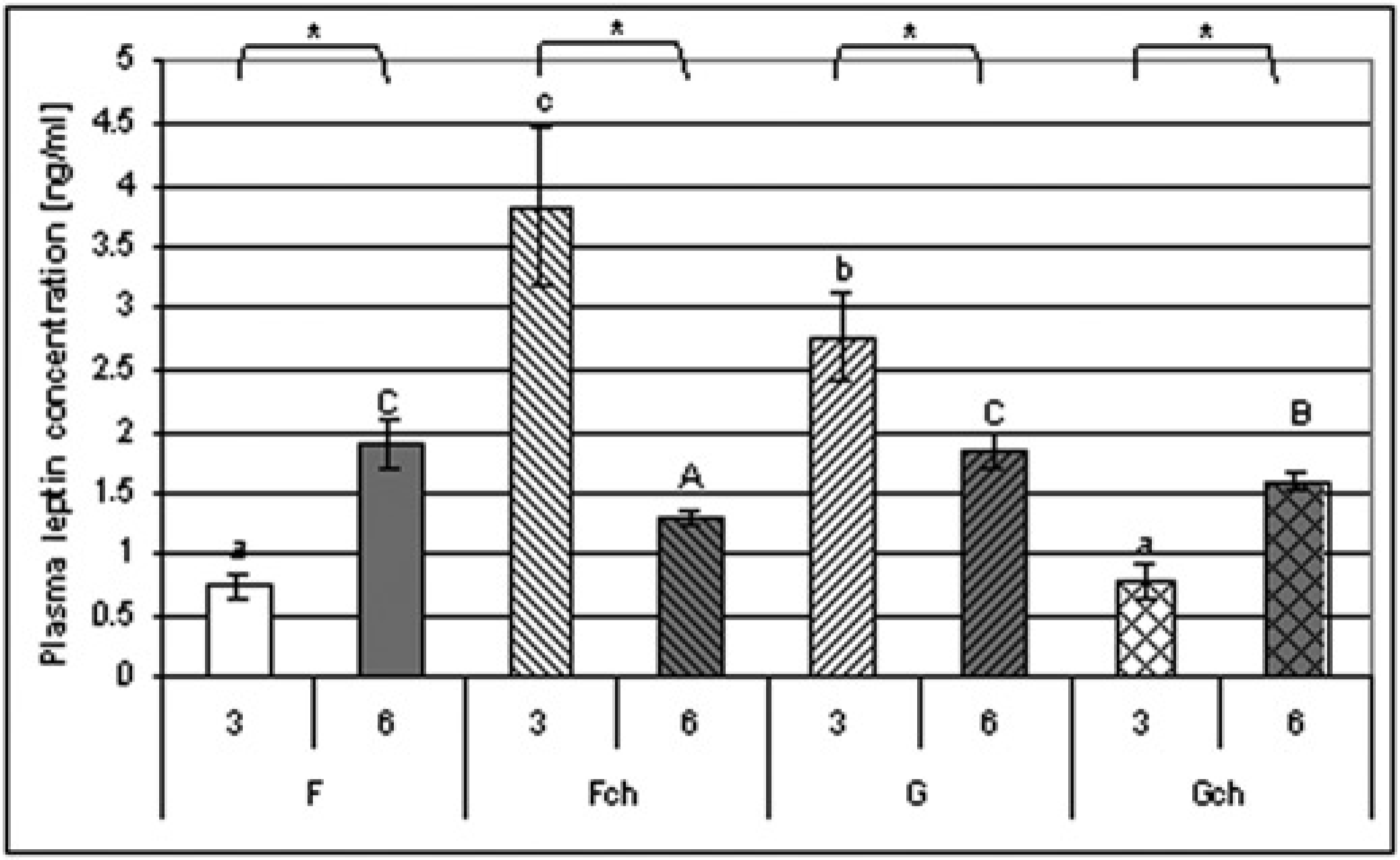
When rats were fed the diet for three weeks, the plasma leptin concentration depended on both fat type (ANOVA, P ≤ 0.02) and cholesterol supplementation (ANOVA, P ≤ 0.02), with the highest value in group 3Fch. Also, interaction between these two factors could be observed. Additional cholesterol caused an increase in leptin concentration in the group fed a diet with fish oil, but decreased leptin in the group fed a diet with grape seed oil (ANOVA, P ≤ 0.0001).
On the other hand, when the experiment lasted for six weeks plasma leptin concentration depended only on cholesterol supplementation (ANOVA, P ≤ 0.0001) with lower values when cholesterol was added. Interestingly, cholesterol supplementation caused higher decreases of leptin level in the group fed a diet with fish oil than the group fed a diet with grape seed oil.
After the three-week experiment, plasma leptin concentration was positively correlated with body weight gain (P ≤ 0.0003, r = 0.97) and plasma TG level (P ≤ 0.005, r = 0.72), but negatively with HDL concentration in plasma (P ≤ 0.007, r= -0.73). However, after a six-week feeding period there was an observed positive correlation between leptin level and HDL concentration in plasma (P ≤ 0.05, r = 20.51).
Leptin expression in the layers of the aorta
The parameter that statistically influenced leptin expression in the aorta was the aortic layer involved. The highest expression of leptin was observed in endothelium after both periods of feeding: three and six weeks (ID score: 0.49 ± 0.2 and 0.87 ± 0.3) with almost equal amounts in both myometrium (ID score: 0.26 ± 0.16 and 0.40 ± 0.15) and adventitia (ID score: 0.20 ± 0.14, 0.46 ± 0.18).
Leptin expression in endothelium, myometrium and adventitia was influenced by the period of feeding (ANOVA P ≤ 0.0009, P ≤ 0.01 and P ≤ 0.0002, respectively), with higher values after six weeks of feeding. Only in groups 3F and 6F did leptin expression not differ; in other pairs higher values were observed in each layer after a longer period of feeding (Figures 3a-c).
Leptin expression in the aorta (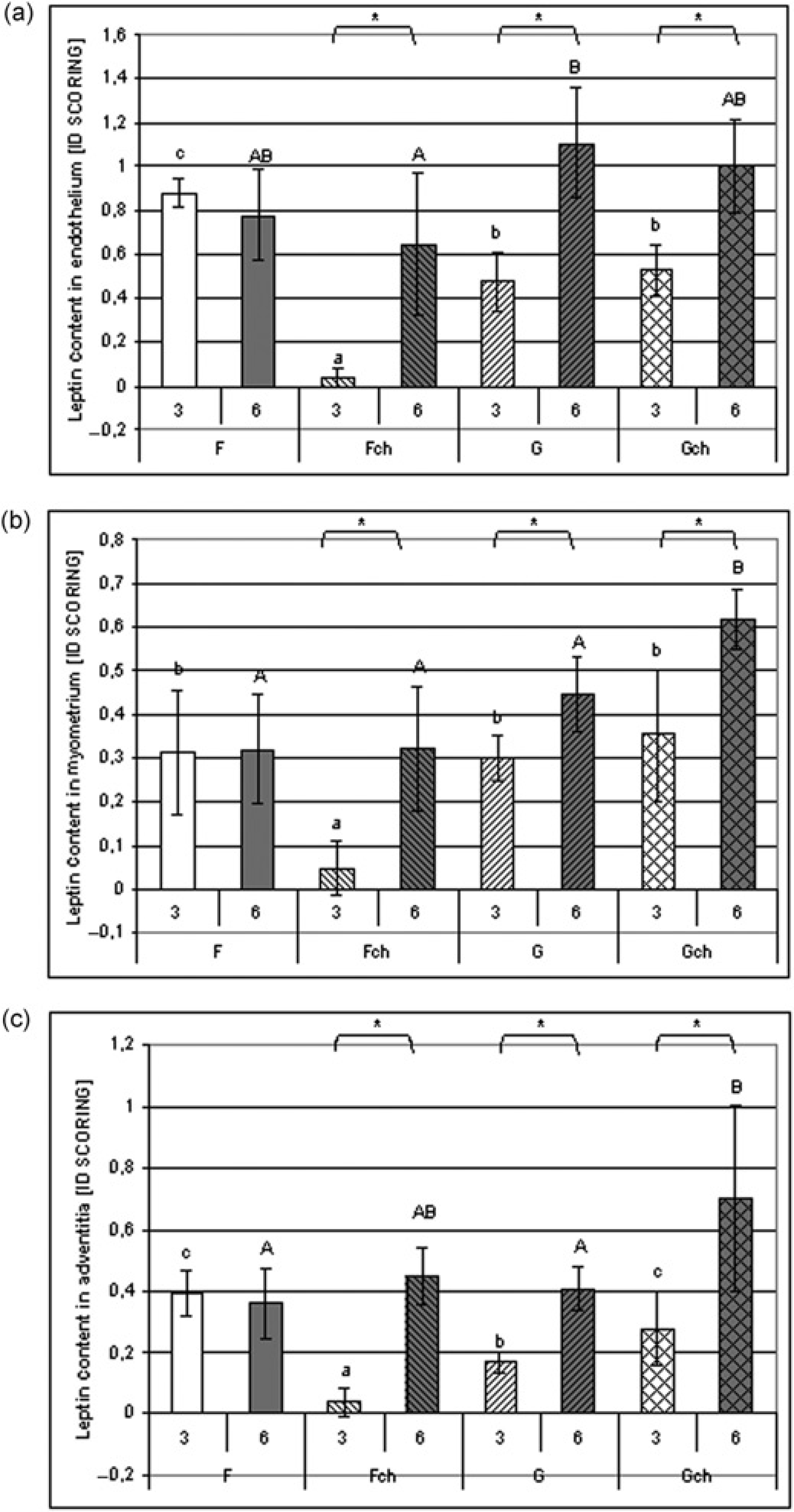
After three weeks, the highest leptin expression in endothelium was observed in group 3F and the expression was lowest in group 3Fch. So, the influence of cholesterol supplementation was observed (ANOVA P ≤ 0.0001), also interacting with fat type (interaction ANOVA P ≤ 0.0001), because when rats consumed a diet with grape seed oil no difference was observed between 3G and 3Gch. The influence of fat type was observed after a longer period of feeding, when higher values were observed after the consumption of a diet with grape seed oil (ANOVA P ≤ 0.01).
Fat type had a significant influence on leptin expression in the myometrium after both periods of feeding (ANOVA, P ≤ 0.02 and P ≤ 0.003 for three and six weeks). After three weeks, the lowest expression of leptin was observed in group 3Fch; and after six weeks the highest value of leptin expression was observed in group 6Gch.
There was a noticeable influence of cholesterol supplementation on leptin expression in the adventitia. However, after six weeks the addition of cholesterol to the diet caused higher values (ANOVA, P ≤ 0.03) but caused lower values after three weeks (ANOVA P ≤ 0.007), mainly because of the very low values of the examined parameters in group 3Fch. The leptin expression was highest in group 6Gch.
Discussion
Undoubtedly one of the main factors promoting abnormal changes in blood vessels is nutrition. A high-fat diet rich in SFA is known to cause atherosclerosis and coronary insufficiency. On the other hand, high-consumption of PUFA, especially n-3, normalizes the function of the cardiovascular system. It was also observed that additional fat in the diet is a reason for leptin increase in the plasma, and because of its proatherogenic actions this adipokine can be treated as a mediator between obesity and atherosclerosis.
The proper state of the cardiovascular system is mainly dependent on the function of the endothelium. It controls blood flow, angiogenesis and permeability of the vessel wall.20,21 Also, this one-layer tissue is responsible for the synthesis of biologically active compounds like NO, endo-thelin, free radicals and cytokines. 22 Any dysfunction of the endothelium can be a trigger for the atherosclerotic process. 23 However, in the development of atherosclerosis, the myometrium and adventitia also play crucial roles. Vascular smooth muscle cell migration, proliferation and calcification cause wall stiffness and thickness. 24 In the adventitia an inflammatory process develops and vascular reactive oxygen radicals are generated. 25
In our experiment, we tried to check whether leptin, which has its receptors in both endothelium and myocytes, will change its concentration in individual layers of the aortic wall (endothelium, myometrium and adventitia) under different modifications of dietary fat type and cholesterol supplementation. All of the animals were fed high-fat (20% w/w) diets, as such an amount highly exceeds rats’ nutritional needs for fat according to American Institute of Nutrition (AIN). 26 We expected that on such diets animals would put on weight and become obese. During the three-week experiment we observed such a tendency, although after a longer period of feeding the animals reduced their food intake and started losing weight. As we observed at autopsy particular the six-week fish oil fed animals had negligible amounts of visceral and subcutaneous fat. Moreover animals on a normal chow diet during the same period gained weight (the average body weight – 2.93 ± 0.369 g/d) and consumed more chow (the average daily diet intake – 6.255 ± 0.188 g/d/100 g body weight)(the unpublished data). Also we can suggest that a high-fat diet, regardless of the fat type, has accelerated the function of a satiety center in the brain, having caused the animals to decrease their food intake. According to Gardiner et al., 27 a high-fat diet can reduce food intake even in ghrelin trans-genic mice and wild-type mice with long-term ghrelin administration. However, inhibition of a hyperphagic effect of ghrelin was not caused by either alterations to food preference or hypothalamic signaling of neuropeptides that control food intake. Also, in our experiment we cannot say exactly which pathway changes resulted in animals lowering their food intake.
In addition to a high-fat diet, in our experiment the addition of 3% cholesterol was used to intensify the atherogenic effect of the diet. In previous publications, there were two types of diets used to trigger atherogenic changes in vessels. The first type of diet was one with a normal fat level (5%) but with lard as a source of SFA with cholic acid (0.5%) and cholesterol (1%). 28 The second type was a diet which is high-fat (20%) with a higher amount of cholesterol (2–3%). 29 Here we decided to use a high-fat diet that allowed us to change the composition of fatty acids by using different dietary fat sources: grape seed oil (rich in PUFA n-6) and fish oil (rich in PUFA n-3). It was stated that such a diet would cause hypercholesterolemia and hypertriglicerydemia, and finally an increase of lipid peroxi-dation, which can ultimately cause pathological changes in the vessel wall. 30
The results obtained in our experiment showed three aspects which need further investigation: (1) the particularly distinctive group 3Fch, (2) the differences in leptin expression in the aorta between groups 3F and 3G and (3) a diet rich in n-6 fatty acids with additional cholesterol as a proatherogenic factor.
The particularly distinctive group 3Fch
Leptin is an adipokine strictly connected with body weight and energy homeostasis. In accordance to this, in our experiment the highest leptin concentration in plasma observed in group 3Fch occurred simultaneously with the elevated daily diet intake and body weight gain. In addition, plasma TG concentration was also increased in this group, suggesting that any changes in TG can be treated as a predictor of excessive consumption. This can be confirmed by a positive correlation between TG and both body weight gain and diet consumption in the three-week experiment, whereas such correlations were not observed for either TC or HDL. Dependence between TG and leptin concentrations can be confirmed by the suggestion made by Banks et al. 31 about leptin, TG and the blood–brain barrier in which they stated that higher TG concentrations decrease leptin transport to the brain, increasing its plasma concentration. But, what is most important in the description of group 3Fch is that parameters describing leptin expression in all three aorta wall layers were lowest in this group.
All of this might suggest that a short period of feeding with a diet rich in n-3 fatty acids from fish oil with the addition of cholesterol increased diet consumption, triggering a physiological action of leptin directing it not to the aorta wall, but keeping it in blood or redirecting it to brain where this adipokine exerts its effect of diminishing the appetite. So, in the six-week experiment, the group fed fish oil with cholesterol (6Fch) consumed even lower amounts of the diet than group 6F, but aorta leptin expression in each layer did not differ between these groups. Correlation between leptin and body weight gain observed only in the three-week experiment can prove that its physiological actions are connected with energy expenditure and food consumption.
Differences in leptin expression in the aorta between groups 3F and 3G
Although daily food intake and body weight gain were the same in groups 3F and 3G, leptin concentration in the plasma was lower when rats were fed a diet with fish oil. The difference between leptin concentration and dietary fat quantity and quality has been studied in our laboratory for many years. In our previous studies, we observed that when a normal fat (5%) diet was consumed by animals, leptin concentration was lower in the group fed a diet with flaxseed oil rich in n-3 fatty acids than in the group fed a diet with lard (rich in SFA) as a fat source. However, no difference between the groups fed a diet rich in n-3 (flax-seed oil) and n-6 (grape seed oil) was seen. 12 Also, Wang et al. 32 found no difference in leptin concentration in animals fed for seven weeks diets rich in n-3 from fish oil or n-6 from safflower oil alone; however, the addition of beef tallow (rich in SFA and MUFA) to a diet with safflower oil resulted in a higher plasma leptin concentration compared with other groups. In our experiment, the results obtained in a three-week experiment contradict the literature. But when we concentrate on the six-week feeding period, no influence of dietary fat source can be seen (6F versus. 6G).
Another difference between the examined groups 3F and 3G is the lipid profile. When rats were fed a diet with fish oil, HDL cholesterol level in the plasma was higher, whereas the TC level was lower. This resulted in an increase in the TC/HDL-cholesterol ratio when grape seed oil was consumed. The TC/HDL ratio is suggested as an important indicator of cholesterol imbalance, 33 which suggests that the consumption of grape seed oil can impair cholesterol metabolism in rats. However, a longer period of feeding in our experiment did not show differences in the TC/HDL ratio between group 6F versus 6G, but TC and HDL levels were higher in the group fed grape seed oil. Lipidogenic properties of grape seed oil were also seen in our previous studies, 12 even when rats were fed normal-fat (5%) diets, when compared with consumption of a diet with flaxseed oil. The lower values of TC observed in groups fed fish oil in these experiments can be explained by a mechanism proposed by Du et al., 34 who suggested a decrease in 3-hydroxy-3-methyl-glutaryl-CoA reductase activity (a crucial enzyme in cholesterol synthesis) in animals fed a diet rich in DHA and ALA.
On the other hand, the correlation between a high level of HDL and a low level of leptin in animals’ blood from group 3F is similar to the correlation observed in ob/ob mice. It was found that the absence of leptin in these mice caused disturbances in HDL plasma clearance and disturbances in intracellular cholesterol distribution. Changes in catabolism and production of apolipoprotein apoA-I and apoA-II are believed to be the biochemical explanation of such a state. 35 Lundasen et al. 36 also stated that the HDL receptor, scavenger receptor class B type I (SR-BI) and the LDL receptor were also suppressed in ob/ob mice. But in our experiment, it should be noted that increased concentration of HDL in plasma was accompanied by the increased expression of leptin in the endothelium in group 3F, in comparison to 3G. We hypothesize that this influx of leptin to the endothelium can be modulated in two ways, connected with the functionality of lipid rafts in the endothelium cell membrane. Lipid rafts are fluctuating units of sphingolipid, cholesterol and different proteins combined together, forming platforms taking part in membrane signaling and trafficking. 37 Firstly, it was established that HDL and apoA-I can participate in cholesterol efflux from lipid rafts causing disruption and vanishing. 38 Raft functions can also be altered by different fatty acid supplies. As it was shown by Ma et al., 39 n-3 PUFA can alter plasma membrane microdomains causing sphingomyelin expression in lipid rafts and the reduction of cholesterol and caveolin in caveo-lae (another type of microdomain). The two presented mechanisms can ultimately lead to changes in membrane properties and consequently to an alteration in the affinity of leptin to endothelium cells. In group 3F, both presented reasons of lipid raft modification could have taken place, as we fed animals with high amounts of n-3 PUFA and observed higher HDL cholesterol concentration with a low leptin level.
A diet rich n-6 fatty acids with additional cholesterol as a proatherogenic factor
The consumption of a diet rich in n-6 fatty acids from grape seed oil for longer periods with or without cholesterol, as well as diet rich in n-3 fatty acids with cholesterol, caused an increase in leptin expression in all three layers. This was opposite to the group fed a diet with fish oil alone. Comparing group 6G and 6Gch, it seems interesting that leptin expression in the adventitia and myometrium was highest in group 6Gch; while in the endothelium it did not differ from group 6G. While plasma leptin expression in the group 6Gch was not either the lowest or the highest among the six-week groups, it is worth discussing here the ways by which leptin gets to the aortic wall. When released from visceral and subcutaneous adipose tissue, leptin easily reaches the systemic circulation, exerting para-crine and endocrine effects on tissues (such as endothelium) and the central nervous system. However, it can also be secreted from perivascular adipose tissue (PVAT), acting directly on vessels like the aorta as there is no anatomical barrier between the adventitia and adipocytes. By this means, leptin can also reach the smooth muscle more quickly.40–42 In our experiment, after both periods leptin expression was generally highest in the endothelium; which could suggest that leptin mainly reaches the aorta from the circulation. However, the lack of difference in endo-thelial leptin expression between groups 6G and 6Gch with the statistical change in the adventitia implies that leptin enters the aortic wall not only from the vessel lumen but also from the opposite site of the aortic wall (Figure 4). Consumption of a diet rich in n-6 fatty acids and cholesterol could enhance leptin production in PVAT, directing it to the aortic wall (adventitia and myometrium) where leptin can act as a proatherogenic factor promoting, for example, smooth muscle proliferation, migration 43 and calcification. 44 Leptin may also contribute to the development of an inflammatory process in the adventitia, but there are no studies devoted to this subject.
Leptin pathways to aortic layers (microscope photo, ×200). 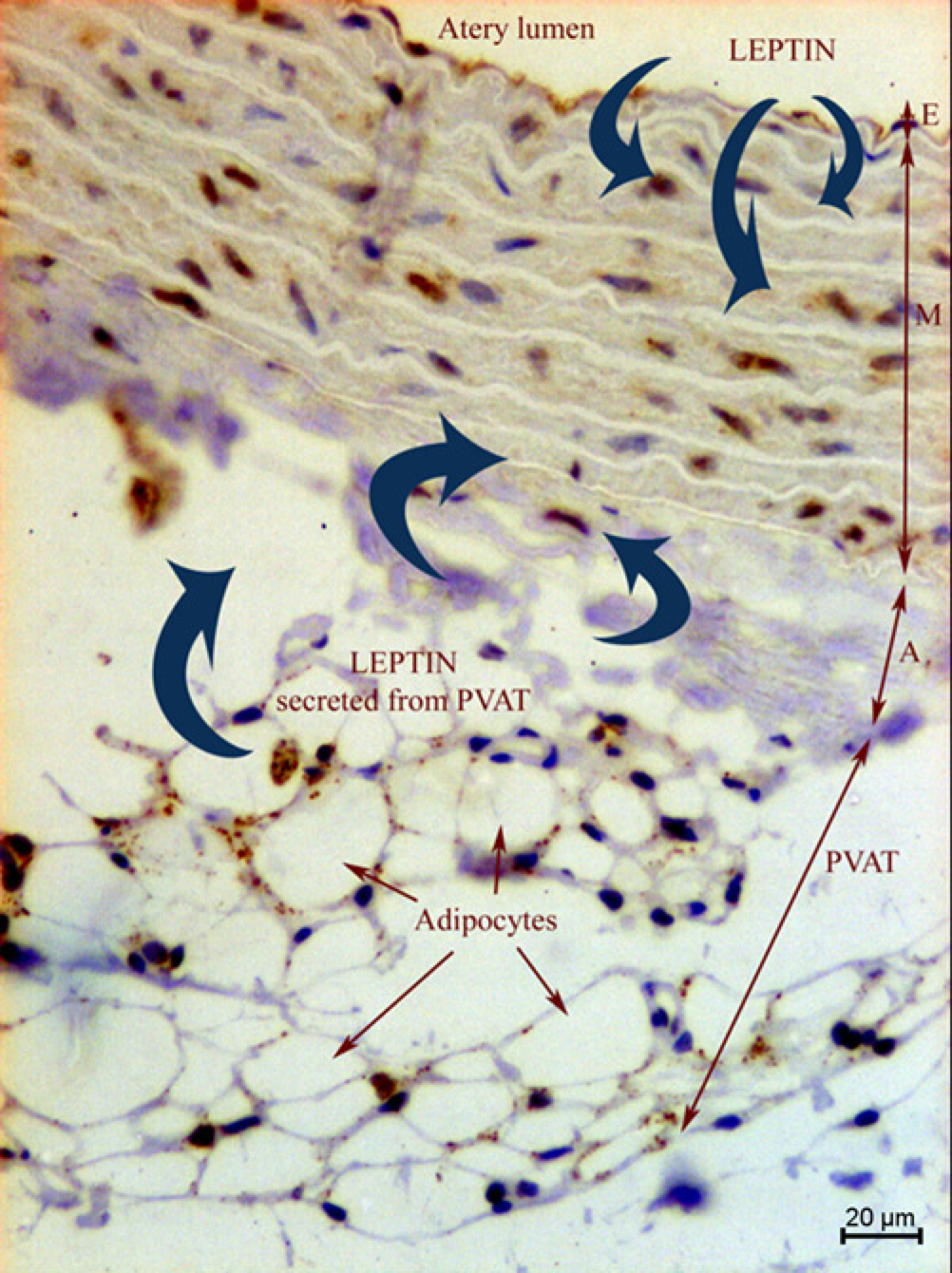
The physiological actions of leptin, which depend on body weight and food intake, can be disturbed by a prolonged consumption of high-fat diet. This may result in leptin being redirected to vessel walls where it can act as a proatherogenic factor. Leptin expression in the aorta is the most common in the endothelium; however, dietary fatty acids or the amount of cholesterol in the diet can enhance its presence in the myometrium and adventitia. Diet-induced changes in the plasma lipid profile in favor of HDL may preferentially sensitize the endothelium to the influx of leptin into the cells, regardless of its concentration in the plasma.
Footnotes
Acknowledgements
This project was supported by grants from the Ministry of Science and Higher Education of Poland (No. 2 P06T 031 27).