
Abstract
This study was undertaken to evaluate the prevalence of GB virus C (GBV-C) viraemia and anti-E2 antibody, and to assess the effect of co-infection with GBV-C and HIV during a 10-year follow-up of a cohort of 248 HIV-infected women. Laboratory variables (mean and median CD4 counts, and HIV and GBV-C viral loads) and clinical parameters were investigated. At baseline, 115 women had past exposure to GBV-C: 57 (23%) were GBV-C RNA positive and 58 (23%) were anti-E2 positive. There was no statistical difference between the groups (GBV-C RNA + /anti-E2 − , GBV-C RNA − /anti-E2 + and GBV-C RNA − /anti-E2 − ) regarding baseline CD4 counts or HIV viral loads (P = 0.360 and 0.713, respectively). Relative risk of death for the GBV-C RNA + /anti-E2 − group was 63% lower than that for the GBV-C RNA − /anti-E2 − group. Multivariate analysis demonstrated that only HIV loads ≥ 100,000 copies/mL and AIDS-defining illness during follow-up were associated with shorter survival after AIDS development. It is likely that antiretroviral therapy (ART) use in our cohort blurred a putative protective effect related to the presence of GBV-C RNA.
INTRODUCTION
GB virus C (GBV-C) or hepatitis G virus (HGV) was discovered by two independent groups of investigators while studying cases of non-A, non-B, non-E hepatitis. 1–3 This virus derives its name from the ‘GB-agent’ described by Deinhardt et al. in 1967. After inoculation with the serum from a surgeon with active hepatitis (who had the initials GB), marmosets developed hepatitis. 4 The GBV-C genome consists of a single-stranded positive sense RNA of approximately 9.4 kb, which has characteristics of a flavivirus-like genome similar to hepatitis C virus (HCV). 5 It appears to encode two glycoproteins predicted to compose the virus envelope proteins E1 and E2. Anti-E2 antibody is directed against the envelope 2 protein of GBV-C and indicates previous exposure to the virus. 6 It is known that the appearance of this antibody is mostly associated with viral clearance.
GBV-C RNA is detected by nucleic acid amplification tests and quantification has been described using terminal dilution methods, realtime (TaqMan) polymerase chain reaction (PCR) and branched chain DNA assays.
Transmission of GBV-C occurs by parenteral, sexual and materno-fetal exposure to the virus; infection is extremely common in humans, being found in 2% and up to 17% of healthy blood donors from industrialized and developing countries, respectively. 7
The prevalence of GBV-C RNA among Brazilian healthy blood donors varies between 5–10%. 8,9 Levi et al. 10 demonstrated 9.7% prevalence of GBV-C RNA in 545 serum samples from blood donors in the state of São Paulo, Brazil.
Although GBV-C was initially thought to cause hepatitis, extensive epidemiological studies found no association between the virus and either acute or chronic liver disease, nor for any other disease entity studied to date. 11 Infection with GBV-C is very common and currently most researchers agree that GBV-C probably replicates primarily in lymphocytes. Furthermore, it does not appear to be hepatotropic and does not replicate effectively in hepatocytes. 12
Since there appears to be no association between GBV-C and any identifiable disease, this virus has been considered non-pathogenic to humans, and pertinent research has slowed down considerably. However, two surprising reports published in 1998 renewed interest in this virus. 13,14 Studies carried out with HIV/GBV-C-infected patients have suggested that GBV-C infection is associated with reduced morbidity with slower progression to AIDS. 15–20 GBV-C modulates lymphocyte functions, which may explain the beneficial associations between GBV-C and HIV-related survival.
The aim of this study was to evaluate the prevalence of GBV-C viraemia and anti-E2 antibody and assess the effect of the interaction between GBV-C and HIV during a 10-year follow-up of women based on mean and median CD4 + cell counts, and HIV and GBV-C quantitative viral loads. We also examined the correlation between the GBV-C loads, CD4 + counts and HIV loads.
PATIENTS AND METHODS
Samples
We evaluated 248 plasma samples from HIV-infected women enrolled in studies on human papillomavirus (HPV) and cervical disease, which were collected between 1996 and 1999. 19 Replicate samples were stored at − 70°C and kept at this temperature, with no thawing. Clinical data from all patients were reviewed and updated by checking their medical records.
The institutional review boards of both recruiting institutions, SEAP – USP and Hospital Emílio Ribas – approved this study and written informed consent was obtained from all participants. The patients were divided into three groups: those who tested positive for GBV-C RNA, those who tested positive for anti-E2 antibodies, and those without evidence of exposure to GBV-C, who were GBV-C RNA/anti-E2 negative.
RNA extraction and cDNA synthesis
RNA was extracted from 140 µL of plasma using the QIAamp Viral RNA mini kit (Qiagen, Hilden, Germany) according to the manufacturer's protocol. RNA was eluted in 60 uL of AVE buffer and stored at − 70°C or directly submitted to cDNA synthesis as described. 21
Measurement of anti-E2 antibodies in serum
Anti-E2 antibodies to GBV-C envelope protein E2 were detected by using the μPlate anti-HG env test (Roche Diagnostics, Penzberg, Germany) according to the manufacturer's instructions.
Nested PCR for GBV-C
For first-round PCR, 5 µL of cDNA was directly used in a 25 µL final volume PCR reaction generating a fragment of 367 bp. Five microlitres of the first-round product was used as template for the second round. A fragment of 208 bp is expected for samples containing GBV-C RNA. 20 All reactions were run in a Mastercycler Gradient thermocycler (Eppendorf, Hamburg, Germany).
Quantitative realtime PCR for GBV-C: TaqMan assay
The GBV-C 5′ non-coding region was chosen as a target for primer design since it was shown by several groups to be the most conserved region of GBV-C among different isolates. 5 This region was submitted to the Primer Express software (Applied Biosystems [ABI], Foster City, CA, USA) and several combinations of primers and probes were retrieved. Each possibility was analysed visually in order to choose the set containing less nucleotide variability, checked against an alignment of GBV-C 5′NCR sequences. We checked as well the primer/probe set where those differences, if unavoidable, would be placed at the first 5′ positions of the oligonucleotides, where they are less prone to interfere with the hybridization to the template. Finally, the following set was selected: sense primer RTG1 (5′GTGGTGGATGGGTGATGACA3′); antisense primer RTG2 (5′GACCCACCTATAGTGGCTACCA3′) and probe (5′FAM-CCGGGATTTACGACCTACC-NFQ3′) where NFQ stands for ‘non-fluorescent quencher’. ABI provided this assay as a 20× concentrated mix. We added 9 µL of cDNA to 10 µL of TaqMan Universal PCR Master Mix 2X and 1 µL of the 20X TaqMan assay that contained primers at 360 µmol/L and the probe at 100 µmol/L. Cycling conditions were 95°C/10 minutes, followed by 40 cycles of 95°C/15 seconds, 60°C/1 minute.
Reference material
A plasma bag previously characterized as containing high titres of GBV-C RNA was kindly provided by the Hospital Sírio Libanês Blood Bank (São Paulo, Brazil) and used to generate a standard curve, as described. 22
Statistical analysis
For descriptive analyses, the means and standard deviations, or the medians and interquartile ranges, were reported. A comparison of the mean age between groups was performed by analysis of variance test. Comparisons of proportions among groups were assessed by chi-square test or likelihood ratio, when appropriate. Continuous and ordinal data were compared by Kruskal–Wallis and Wilcoxon signed rank sum. The Spearman's rank correlation coefficient was used to assess correlations between the continuous variables. Differences in survival rates were assessed by Kaplan–Meier analysis. Equality of survival distributions was evaluated by log-rank test. A Cox proportion-hazards regression model of variables such as CD4 + cell count and HIV load at baseline, number of antiretroviral therapy (ART) regimens, opportunistic infections and GBV-C status (RNA-positive, anti-E2 positive, or without a marker of exposure to GBV-C) was used to examine the effect of these covariates on survival. A significance level of 0.05 was used and all of the P values are two-sided. All the analyses were conducted in SPSS version 12 (2002).
RESULTS
Of the 248 HIV-positive patients studied, 115 had been exposed to GBV-C, 57 (23%) being GBV-C RNA + /anti-E2 − and 58 (23%) GBV-C RNA − / anti-E2+. The overall prevalence of GBV-C infection in this population was 46%. Concomitant presence of GBV-C RNA and anti-E2 antibodies was not found in any of the patients. The majority of patients had acquired HIV infection by sexual contact. There were no significant differences regarding age and route of HIV transmission among groups (P > 0.05). When compared with subjects who were GBV-C RNA + /anti-E2 − , GBV-C RNA − /anti-E2+ and GBV-C RNA − /anti-E2 − , median baseline CD4 + counts and plasma HIV viral loads were similar among groups, as shown in Table 1. When we analysed the association of CD4 + count, HIV load and GBV-C load at baseline, we found no significant association between GBV-C viral load and CD4 + count (r s = 0.097, P = 0.129), nor a significant inverse correlation between GBV-C viral load and HIV viral load (r s= − 0.058, P = 0.366).
Baseline characteristics of the 248 HIV-infected women according to GBV-C status
GBV-C = GB virus C; IDU = injecting drug user; IQR = interquartile range (P 25 − 75%)
*Kruskal-Wallis test
†Chi-square test
‡Likelihood ratio
Follow-up
By the end of the follow-up period, we had observed a total of 43 deaths. Only five of the 57 subjects positive for GBV-C RNA and 31 of the 133 subjects negative for GBV-RNA/anti-E2 had died from AIDS-related events. We calculated the relative risk of death considering GBV-C status using the Cox model. There was a statistically significant difference when comparing groups GBV-C RNA − /anti-E2 − and GBV-C RNA + / anti-E2 − (P = 0.038). The relative risk of death for the GBV-C RNA positive group was 63% lower than for the unexposed group. The comparison between GBV-C RNA − /anti-E2 − and GBV-C RNA − /anti-E2+ subjects was not statistically significant (P = 0.089).
We observed a higher prevalence of tuberculosis cases (18.9%) as compared with oro-esophageal candidiasis (14%) and toxoplasmosis (17.3%). However, there was no statistical difference on the occurrence of these opportunistic diseases between the GBV-C groups. Nineteen patients acquired hepatitis C, all confirmed by PCR. Three patients acquired hepatitis B (HBV) and five human T-cell lymphotropic virus (HTLV) 1/2. There was no statistical difference between the GBV-C RNA groups and chance of HCV, HBV and HTLV acquisition, P = 0.864, 0.987 and 0.39, respectively.
Considering ART, the median number of different ART regimens used during follow-up was four, and it was similar between groups (Table 2, P = 0.583). Regarding the reason for change of ART (immunological/virological failure and side-effects related to medication), there was no significant difference between the GBV-C groups (data not shown).
Mean and median of antiretroviral regimens used during follow-up according to GBV-C status
GBV-C = GB virus C; IQR = interquartile range
*Kruskal–Wallis test
Survival analysis
Of the 248 patients followed in our analysis, an outcome of AIDS-related death was observed in 43 patients. Among the remaining 205 patients, 164 completed the follow-up period up to January 2008, whereas 41 were lost to follow-up and data were censored at the time of their last visit. The mean survival time achieved by the Kaplan–Meier analysis stratified by GBV-C groups considering the outcome of death related to AIDS was 9.45 years (95% confidence interval [CI] 8.95–9.95) for GBV-C RNA + /anti-E2 − ; 9.32 years (95% CI 8.78–9.85) for GBV-C RNA − /anti-E2+ and 8.65 years (95% CI 8.19–9.10) for GBV-C RNA − /anti-E2 − . This analysis exhibited a log-rank with P = 0.034, with a statistically significant difference in survival rates in 10 years of follow-up between the GBV-C groups as depicted in Figure 1.
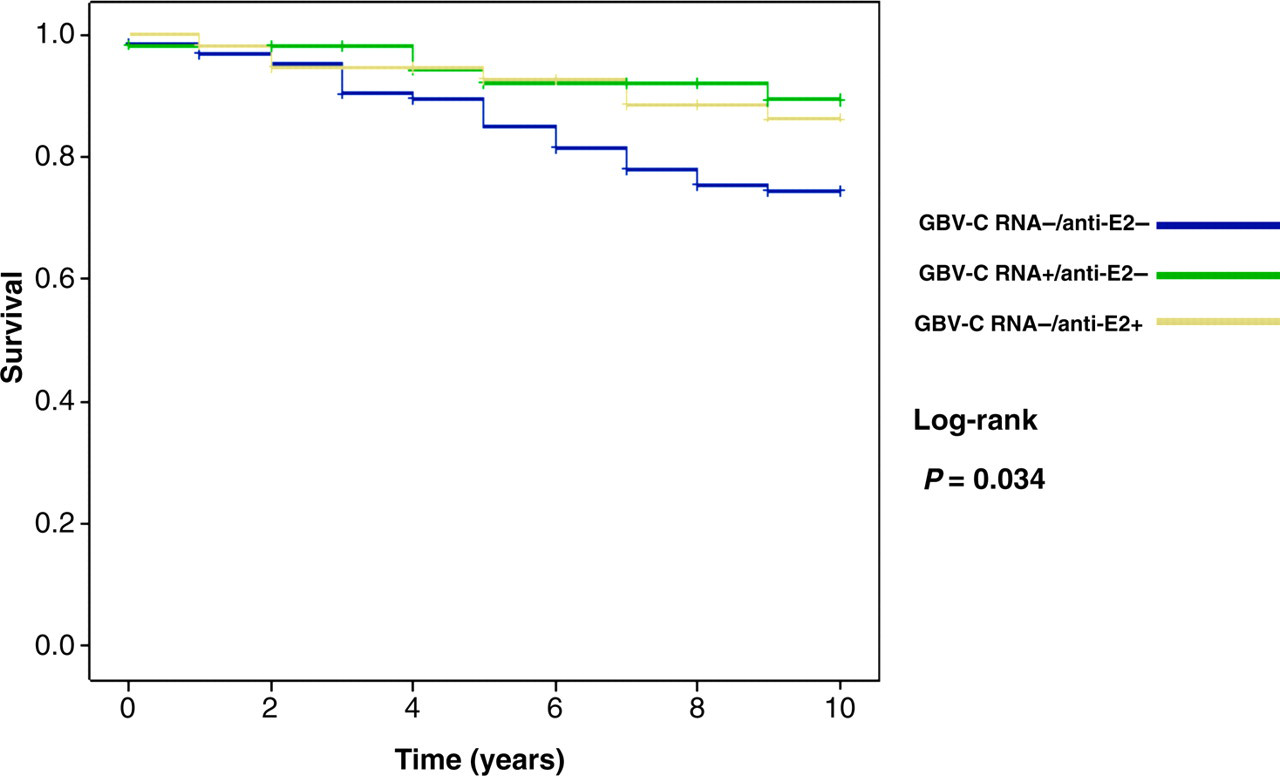
Survival analysis according to GBV-C status (Kaplan–Meier). GBV-C = GB virus C
Multiple Cox regression analysis was applied to the variables CD4 + count and HIV load at baseline, number of ART regimens, AIDS-defining illness and GBV-C groups. Only the variables HIV viral load ≥100,000 copies/mL and presence of AIDS-defining illness during follow-up demonstrated a statistically significant negative association with survival. HIV load ≥100,000 copies/mL compared with low baseline HIV load (<400 copies/mL), and/or the presence of an AIDS-defining illness during follow-up had a relative risk of death 9.34 and 3.76 times higher than the referent group, respectively, regardless of the GBV-C group in question (Table 3, P > 0.05).
Multiple regression analysis of selected Cox-adjusted variables in 248 patients. Outcome: risk of death
CI = confidence interval; RR = relative risk
*Chi-square test
DISCUSSION
Prevalence studies have demonstrated that GBV-C is a virus present in most HIV-positive patients, being detected at rates greater than those described for healthy blood donors. This viral co-infection can be explained by shared modes of transmission, the parenteral and sexual routes being the most important. In Brazil, few investigations have been carried out on the presence of GBV-C among the HIV-infected. Our study showed a prevalence rate of 23% for both GBV-C RNA and anti-E2, giving a 46% exposure rate to this virus. These results are similar to the findings described in North American and European studies. 23
Our group of patients was composed exclusively of women, mostly in their mid-thirties, and whose exposure to HIV was primarily sexual (67.2%). Most of these patients reported stable HIV-positive partners. The literature confirms the dominance of parenteral and sexual routes as the main modes of HIV transmission. 21
In our study, the presence of GBV-C viraemia in HIV patients was not associated with higher median baseline CD4 + counts or with lower median baseline HIV loads when compared with GBV-C RNA-negative HIV patients. Similarly, Giret et al. 22 studied a Brazilian cohort of 175 HIV-infected patients and did not find a significant correlation between GBV-C RNA and CD4 + count at baseline.
Co-infection with HBV and HCV occurs frequently due to shared modes of transmission (parental, sexual, vertical). Since we lacked information on the presence of hepatitis B and C at baseline, it was not possible to establish how many patients were carriers of these viruses at the time we started the study. However, we were able to detect the acquisition of viral hepatitis during the follow-up, with a higher incidence of HCV (11.7%) than HBV (1.8%), although there was no predominance of either hepatitis associated with a specific GBV-C status. The majority of AIDS-defining illnesses observed during follow-up were disseminated tuberculosis, oro-esophageal candidiasis and cerebral toxoplasmosis, and the frequency of these infections was similar among the GBV-C groups. This result, however, should be interpreted with caution because we compared a laboratory variable (GBV-C RNA) with a clinical outcome (AIDS-defining illness). We must also consider that our patients were on ART, which in itself should have a beneficial effect on the control of opportunistic infections regardless of GBV-C status. In addition, persistent co-infection with GBV-C RNA appears to be necessary for the beneficial effects of this virus during ART. Also, clearance of GBV-C RNA early in the course of HIV was found not to promote maintenance of the beneficial effects. 11 Williams et al. 24 demonstrated that HIV-infected patients in whom GBV-C RNA was cleared during follow-up showed a higher mortality rate than those with persistent GBV-C RNA.
Relatively little is known about the effect of GBV-C co-infection on the response to ART in HIV-infected subjects. 11 In the analysis of exposure to ART regimens, we found that our population made use of a similar number of ART regimens and was heavily treatment experienced during their follow-up. We also obtained information on the reasons for change of ART from medical records of 129 patients, and we defined reason for change as immunological/virological failure or adverse events. However, we found no statistically significant association between the GBV-C groups and reason for change of ART.
The GBV-C RNA-positive group had a better survival, related to AIDS, by Kaplan-Meier analysis. However, using multiple Cox regression, we demonstrated that only patients with HIV load ≥100,000 copies/mL and/or presence of AIDS-defining illness during follow-up had a higher relative risk of death, regardless of GBV-C viraemia. In other words, we conclude that in our study the protective effect of GBV-C on patient survival was not an independent factor responsible for increased CD4 + cell count and reduced HIV load, which are the surrogate markers widely used in clinical management as reliable prognostic factors for HIV disease progression.
We concur with the concept that ART has been an important determinant of survival in the natural history of HIV infection and are aware that, in our study, ART probably blurred any putative protective effect of GBV-C viral infection on HIV viral load and CD4 + counts.
One limitation of our study is that the survival analysis of the 248 patients investigated began in 1996, covering a period marked by the introduction of highly active ART in several reference centres for treatment of sexual transmitted infections and HIV/AIDS in Brazil.
Further studies on the effect of GBV-C/HIV co-infection on the response to ART in treatment-experienced HIV-infected patients are required. It is possible that GBV-C acts as a marker for the presence of other factors that delay HIV progression or, alternatively, GBV-C may act directly on the reduction of HIV replication. However, this effect is difficult to be proven in vivo, due to the many variables involved in the rise and fall of HIV viral load.
Footnotes
ACKNOWLEDGEMENTS
This study was supported by Fundação de Amparo a Pesquisa do Estado de São Paulo (FAPESP), Grant number 05/01072-9.