We consider a reaction–diffusion system including discontinuous hysteretic relay operators in reaction terms. This system is motivated by an epigenetic model that describes the evolution of a population of organisms which can switch their phenotype in response to changes of the state of the environment. The model exhibits formation of patterns in the space of distributions of the phenotypes over the range of admissible switching strategies. We propose asymptotic formulas for the pattern and the process of its formation.
In this paper, we consider a reaction–diffusion equation that includes discontinuous two-state two-threshold relay operators. This equation was proposed as a model for dynamics of a population of bacteria that can switch their phenotype in response to changes of the environment [17,20]. The model exhibits formation of patterns in the space of distributions of the two phenotypes over an admissible range of the switching thresholds. The convergence of solutions to a stationary pattern observed numerically in [17] was proved in [21]. The objective of this work is to obtain asymptotic formulas for the pattern and the timing of its formation under the assumption of slow diffusion.
The model includes several time scales. The “spatial” variable is a scalar parameter x that measures the distance between the switching thresholds of a bacterium and characterizes its switching strategy (the smaller the x the more responsive is the strategy to variations of the environmental conditions). That is, a solution describes the evolution of the distribution of each of the two phenotypes over possible switching strategies parameterized by x. The diffusion of this distribution models sporadic changes of the switching strategy by individual organisms; it is assumed to be slow compared to the rate of growth and competition precesses. The rate of transition from one phenotype to the other in bacteria is assumed to be much higher than the rate of other processes. Every such transition is modeled by an instantaneous switch of a relay from state 1 to state or vice versa (where each state represents a particular phenotype).
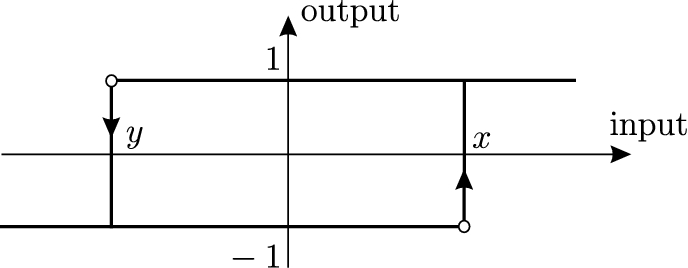
The central assumption that we make is that the switching threshold for the transition of a relay (bacterium) from state 1 to state is different from the switching threshold for the opposite transition (the difference of the thresholds is called the bi-stability range). That is, we assume hysteresis, or multi-stability, in the switching response of bacteria to the exogenous stimulus. This hysteresis can be viewed as form of the persistent memory of past environmental conditions by organisms. Since the seminal work of Max Delbrük [14], epigenetic differences arising in the process of cell differentiation have been attributed to multi-stability of living forms. A classical example of such multi-stability is the behavior of lac-operon in E. coli (lac-operon is a collection of genes associated with transport and metabolism of lactose in the bacterium; expression of these genes can be turned on by molecules called inducers). Experimental studies of the regulation of enzymes in E. coli and yeast that were conducted as early as in the 1950–1960s effectively demonstrated that two phenotypes each associated with “on” and “off” state of lac-operon expression can be obtained from the same culture. The fraction of each phenotype in the total population depended on the history of exposure to the inducer, and each phenotype remained stable through multiple generations of the bacteria [5,10,11,39,40,47,51]. This behavior resembles the two-threshold hysteretic relay shown in Fig. 1. Later, multi-stable gene expression and hysteresis have been well-documented in many natural as well as artificially constructed systems [4,15,19,41,45]. In particular, recent experiments using molecular biology methods permitted a further study of the region of bi-stability of the lac-operon when multiple input variables are used to switch the lac-operon genes on and off.
Non-ideal relay.
A substantial amount of experimental and theoretical work provides an evidence that organisms use various (often randomized) strategies of phenotype switching in order to adapt to changing environmental conditions (see [36] and references therein). It is intuitively clear that phenotypic diversity within the population can help to increase the chances of survival in varying environment. Switching strategies that establish phenotypic diversity are called bet-hedging [3]. Indeed, experimental and theoretical models of adaptation demonstrate that bet-hedging can evolve to maximize the net growth of the total population [1,2,28,38,42,48]. In [18], a simple differential model was used to show that switching strategies with two thresholds where the switching moment depends not only on the state of the environment, but also on the phenotype itself, can further increase Darwinian fitness of species. These strategies exploit hysteresis and are described by a two-threshold relay shown in Fig. 1 in the adiabatic limit when switching is faster than the characteristic rate of the environment variations. It was also shown in [18] that the optimal bi-stability range, that is the optimal separation of thresholds of the relay, which maximizes the net growth rate of the population is different for different environmental inputs. For this reason, the model that is considered in this work includes the distribution of bacteria (relays) over a set of switching strategies which have different separation of switching thresholds with x taking the values from an interval . The diffusion process included in the model acts as a factor that diversifies the switching thresholds (strategies) in the population, while the competition provides a mechanism of selection that may favor a subpopulation with a specific x for a given law of variation of the state of environment. More detailed discussion of the model assumptions and the biological background can be found in [17,18,20].
Parabolic equations with distributed non-ideal relays were previously used for modeling spatial structures that were observed in spatially distributed colonies of bacteria, for example concentric rings in Petri dish experiments [9,22,23,26,27,37]. In these models, all the relays have the same fixed separation of thresholds. The mechanism of pattern formation discussed in this work is based on the long-term memory of the hysteretic relays and is related to dynamics of free boundary separating the domain where relays are in state 1 from the domain where relays are in state on the interval . This mechanism is different from the mechanism found in [9,26,27]. Other important aspects of dynamics of parabolic and hyperbolic differential equations with distributed relays were studied in [6,8,12,16,24,25,29,30,33,49,50] in relation to multiple applied problems (not related to patterns) such as phase transitions, population dynamics, multi-phase flows in porous media, thermostat control and others.
In the following Sections 2 and 3 we present the model and a statement on its well-posedness that has been obtained earlier. Section 4 contains the main statement on the asymptotics of the patterns and its proof.
Model description
In this paper, we consider a class of models, which attempt to account for a number of phenomena, namely (a) switching of bacteria between two phenotypes (states) in response to variations of environmental conditions; (b) hysteretic switching strategy (switching rules) associated with bi-stability of phenotype states; (c) heterogeneity of the population in the form of a distribution of switching thresholds; (d) bet-hedging in the form of diffusion between subpopulations characterized by different bi-stability ranges; and (e) competition for nutrients. The resulting model is a reaction–diffusion system including, as reaction terms, discontinuous hysteresis relay operators and the integral of those. This integral can be interpreted as the Preisach operator [7,13,31,32,34,35,43,44,46,50] with a time dependent density (the density is a component of the solution describing the varying distribution of bacteria). In [21], we have shown that fitness, competition and diffusion can act together to select a non-trivial distribution of phenotypes (states) over the population of thresholds. The main goal of this paper is to give a quantative description of this distribution for small diffusion.
We assume that each of the two phenotypes, denoted by 1 and , consumes a different type of nutrient (for example, one consumes lactose and the other glucose). The amount of nutrient available for phenotype i at the moment t is denoted by where . The model is based on the following assumptions (see [20] for further discussion).
Each bacterium changes phenotype in response to the variations of the variable , which measures the deviation of the relative concentration of the first nutrient from the value in the mixture of the two nutrients.
The input is mapped to the (binary) phenotype (state) of a bacterium , where is the non-ideal relay operator with symmetric switching thresholds x, with ; see Fig. 1 and the rigorous definition (3.1) in Section 3.
The population includes bacteria with different bi-stability ranges , where the threshold value x varies over an interval . We will denote by the density of the biomass of bacteria with given switching thresholds at a moment t.
There is a diffusion process acting on the density u.
At any particular time moment t, for any given x, all the bacteria with the switching threshold values are in the same state (phenotype). That is, is the total density of bacteria with the threshold x at the moment t and they are all in the same state. This means that when a bacterium with a threshold sporadically changes its threshold to a different value x, it simultaneously copies the state from other bacteria which have the threshold x. In particular, this may require a bacterium to change the state when its threshold changes.
With these assumptions, we obtain the following model of the evolution of bacteria and nutrients,
where and are the derivatives of the population density u; is the diffusion coefficient; dot denotes the derivative with respect to time; and all the non-ideal relays , , have the same input . We additionally assume the growth rate based on the mass action law for bacteria in the phenotype . The rate of the consumption of nutrient in the equation for is proportional to the total biomass of bacteria in the phenotype i, hence to the integral; and are the lower and upper bounds on available threshold values, respectively.
We assume that a certain amount of nutrients is available at the initial moment; the nutrients are not supplied after that moment. We assume the Neumann boundary conditions for u, that is no flux of the population density u through the lower and upper bounds of available threshold values.
Rigorous setting of a well-posed model
Rigorous setting
Throughout the paper, we assume that .
We begin with a rigorous definition of the hysteresis operator (non-ideal relay) with fixed thresholds . This operator takes a continuous function defined on an interval to the binary function of time defined on the same interval, which is given by
where is either 1 or (initial state of the non-ideal relay ). Since may take different values for different x, we write . The function of taking values is called the initial configuration of the non-ideal relays. In what follows, we do not explicitly indicate the dependence of the operator on .
In this paper, we assume that is simple, which means the following. There is a partition of the interval such that the function , which satisfies for all , is constant on each interval and has different signs on any two adjacent intervals:
where the second relation holds if .
We define the distributed relay operator taking functions to functions by
The function will be referred to as the configuration (state) of the distributed relay operator at the moment t.
We set
Here the first integral is the total mass of bacteria; is the so-called Preisach operator with the time dependent density function u. Further, we replace the unknown functions and in system (2.1) by the functions (total mass of the two nutrients) and (deviation of the relative concentration of the first nutrient from the value ). Substituting
into the first equation of (2.1), we obtain the equation
Furthermore, summing the second and the third equations of system (2.1) and using the relationships , and the notation (3.4), we get
Finally, from the second and the third equations of (2.1), it follows that
Thus, the resulting system, which is equivalent to equations (2.1), takes the form
where we assume the Neumann boundary conditions
and the initial conditions
Well-posedness
The problems (3.5)–(3.7), which contains a discontinuous distributed relay operator, was shown in [20] to be well posed. We briefly summarize this result before proceeding with the analysis of long time behavior.
Set for . We will use the standard Lebesgue spaces and ; the Sobolev spaces , ; the anisotropic Sobolev space with the norm
and the space
Assume that . We say that is a (global) solution to problems (3.5)–(3.7) if, for any , , is a continuous -valued function for , and relations (3.5)–(3.7) hold in the corresponding function spaces.
The stateof the distributed relay operator is simple for all;
We havewhere the convergence takes place as.
The behavior given by (3.8)–(3.10) is to be expected. Indeed, as we assume no supply of nutrients after the initial moment, the total amount of nutrients converges to zero. When the density of nutrients vanishes as a result of consumption by bacteria, the equation for the density u approaches the homogeneous heat equation with zero flux boundary conditions, which explains why the density of bacteria converges to a uniform distribution over the interval as a result of the diffusion.
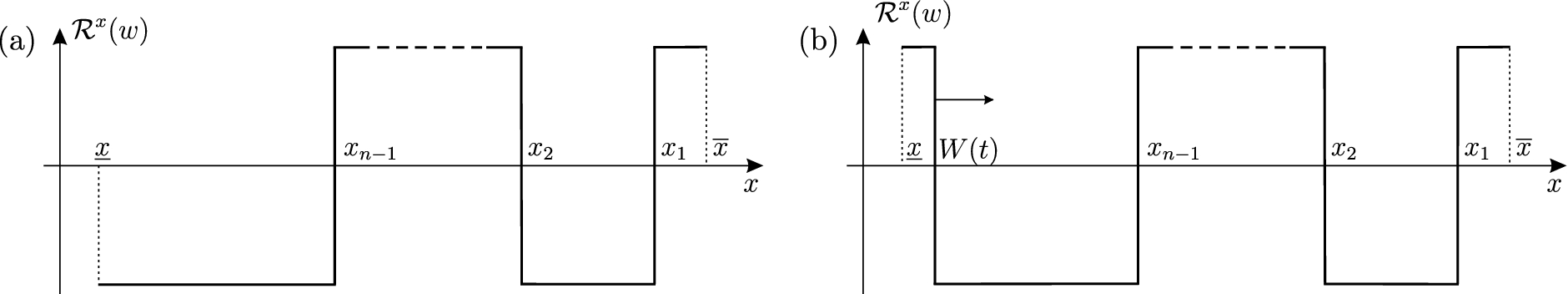
Below we are interested in the limiting behavior of the configuration function given by (3.3), or, in other words, in the limiting distribution of phenotypes over the population of thresholds. In [21], we have proved that converges to a step like profile as , see Fig. 2. In the next section, we give a quantative description of this phenomenon under the assumptions that the initial amount of nutrient and the diffusion coefficient D tend to zero, while the initial density tends to the delta function.
Spatial configuration of the relays at a moment . (a) The input satisfies . Hence, the relays do not switch and all the fronts are steady. (b) The input satisfies . Hence, the relays may switch and the nth front can move.
Fronts asymptotics
Terminology
A point which separates an interval on which the relays are in state 1 from an interval on which the relays are in state is called a front; cf. (3.2). The total number of fronts can vary, but, by Theorem 3.1, remains finite at all times (equivalently, the state of the distributed relay operator remains simple at all times). A front can either stay (a steady front) or move right. That is, any is a non-decreasing function on the time interval of its existence. A front disappears if it hits another front or is hit by another front. Assume a front exists at a moment . It is called immortal up to time T, , if it does not disappear on the time interval .
Our goal is to give asymptotic formulas for the time moments at which steady immortal (up to time T) fronts appear as well as for their positions. This will be done as , where D is the diffusion coefficient in (3.5) responsible for random fluctuations of hysteresis thresholds of individual bacteria.
Assumptions
From now on, we fix a time interval , . Along with conditions (3.7), we assume that the initial density satisfies the relation
where will be chosen small enough and depending on D.
We introduce the function
and set
Set
Using [21, Lemmas 4.1 and 5.4] and the fact that (see (3.7) and (3.8)), we can choose positive functions and such that as and
provided that satisfies (4.1) with and that .
The main assumptions will be as follows.
The initial density satisfies (4.1) with , where is the above function.
The initial amount of food is small: , where is the above function.
The initial value of the input is close to : , where is an arbitrary function such that and as .
The initial configuration is .
Our goal is to determine the consecutive time moments at which the moving fronts become steady and their positions () at these moments.
We begin with the following recursive algorithm for determining (finitely or infinitely many) positive numbers
Algorithm
Basis. Set , and let be the (unique) root of the equation , .
Set . Note that and for all .
Inductive conjecture. Fix . Assume that we have defined the sequences and such that the function
satisfies
As we have seen, relations (4.8) hold for .
Below we will also use the equivalent recursive representation of :
Inductive step. Now we will determine , , and the function satisfying relations (4.9) with n replaced by .
Consider the equation
Due to (4.8) and the fact that as , the left-hand side is a strictly increasing function of s that tends to ∞ as . Therefore, Eq. (4.10) has a unique root, which we denote by .
Consider the function
Relations (4.8) imply that
On the other hand, due to (4.9),
and, hence,
due to the inductive conjecture (see (4.8) with n replaced by ).
Thus, we see that the equality uniquely defines a smooth positive function , , such that
Now we have several possibilities depending on the geometry of the curve defined by (4.14):
for . In this case, we set .
There is such that for and for .
Otherwise, we terminate the process at the finite sequences and .
In these two cases, it remains to check that the function
satisfies relations (4.8) with n replaced by . This will guarantee that we can repeat the inductive step of the algorithm from Section 4.3 with n replaced by .
Combining (4.16) with (4.11), (4.14) and (4.15), we have
On the other hand, in cases (3)(a) and (3)(b), we have for . Therefore, relations (4.11), (4.15) and (4.16) imply that
Main result
Formulation of the main result
Assume that, for some , we have the sequences and constructed according to the above algorithm, and assume that , where T was fixed in Section 4.2.
For all, the nth moving front becomes steady and immortal (up to time) at a momentand its position at this moment isherestands for functions of D that tend to 0 as. Furthermore,.
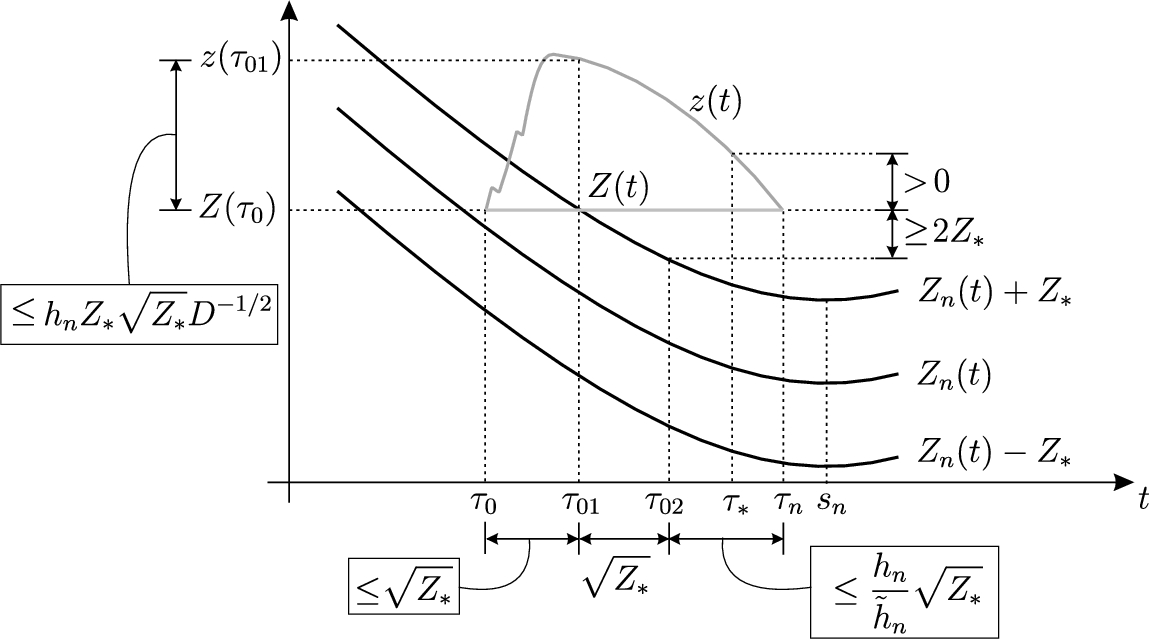
Preliminary discussion
In Section 5, we will prove Theorem 4.1. Assume we have constructed fronts that became steady at the moments at the positions given by (4.17) and (4.18), respectively. We will consider the formation of the nth front on the time interval . We consider the case of an even . The case of odd n is analogous; see also Remark 5.1 below concerning .
Below we will consecutively consider time moments
which are characterized by the following properties:
The function is increasing and the relays do not switch during the time interval ; furthermore,
The function is increasing and some relays switch during the time interval ; further, for some (which will be chosen below to satisfy relations (5.15)), we have
Note that, although the distance between and is already , it is still much larger than the desired distance due to (5.12). Also the moment , which is of order , may be much smaller than the desired moment, which is of order .
The function may oscillate and the relays may, though not necessarily, switch during the time interval . The moment is already of the correct order, but the value may still have a wrong asymptotics. However, it satisfies
i.e., it is bounded from above by the correct asymptotics.
The function may oscillate and the relays may, though not necessarily, switch during the time interval . The moment is of the same (correct) order as , but now, additionally, the value has the correct asymptotics
The function may oscillate and the relays may, though not necessarily, switch during the time interval . The moment is of the same (correct) order as and , and the function does not oscillate any more after the moment . More precisely, is decreasing and the relays do not switch during the time interval . Moreover, the nth immortal front is formed at the moment in the sense that
Proof of the main result
In this section, we will prove Theorem 4.1. Recall that, without loss of generality, we assume that n is even. In Sections 5.1–5.5, we will consider in detail the respective time intervals from items (1)–(5) of Section 4.4.2.
Dynamics for as
We will see below that is increasing for and achieves the value . Let be the moment when this happens: . Note that if and , then the relays do not switch.
Assume we have proved at the previous step (for n replaced by ) that
for some not depending on D (we shall do this for in the end; see (5.33)). As long as remains positive and holds, the relays do not switch, see Fig. 2(a). Hence, using (4.5) and (4.18), we can write the third equation in (3.5) as follows (we omit “” for brevity):
(here and below, we consider only).
We set
Taking into account (5.2), definition (4.2) of , definition (4.7) of , and the fact that n is even, we see that satisfies
where
Due to (4.8) and (5.5), the right-hand side in (5.3) is positive for , provided D is small enough. Together with (5.1), this means that the solution to (5.3), (5.4) increases until a moment (if it exists) at which it achieves the value . This proves that (5.3) is equivalent to the third equation in (3.5) not only for but actually for .
The following lemma determines the time moment .
The equationhas a unique root on the interval. Denote it by. Thenand.
Due to (4.8) and the fact that as , Eq. (5.6) has a unique root .
To prove the asymptotics for , we denote by the solutions of
with the same initial data at as in (5.4). Obviously,
Let be the first moment at which . Then, integrating the differential equation for , we obtain
Using the implicit function theorem in a neighborhood of the point , we obtain . Therefore, taking into account (5.7), we have .
The inequality has been proved before the lemma. □
Dynamics for as
By Lemma 5.1, . Therefore, as long as remains positive, the input increases and hence switches the relays. However, if becomes negative, the relays will stop switching. To describe the dynamics of , we introduce the function
where is defined in Lemma 5.1, see Fig. 2(b). Then, similarly to (5.2), we obtain from the third equation in (3.5) and from relation (4.5)
Setting
we have
where
with defined in (4.3). We consider Eq. (5.8) with the initial condition
We begin with the following observation, which shows that the th front is immortal.
The solutionto problem (5.8), (5.10) cannot achieve the value.
Assume, by contradiction, that achieves the value () for the first time at some moment (). At this moment . Since , it follows from (4.8) (with n replaced by ) that . Therefore, using (5.8) and (4.9), we have
for all sufficiently small D, which is impossible. □
To proceed, we will need a specific (positive) function such that
To define it, we introduce the new function and set
Then we consider Eq. (5.8) for , where is the time moment defined in the inductive step (1) of the algorithm from Section 4.3. For such t, it is equivalent to
where is given by (4.11). We choose the desired to satisfy, for all ,
where is the same function as in (5.14) and does not depend on t and D. This can be done because for all .
The following lemma defines a time moment satisfying (4.20).
There is a time momentsuch thatfor alland.
As long as and , we have . Therefore, we can rewrite Eq. (5.8) as follows:
where, due to (5.9) and (5.12),
Due to (4.8) and (5.17), the right-hand side in (5.16) is negative for , provided D is small enough. Hence, for .
Denote by the solution to the equation
with the same initial condition as in (5.10), .
Then, we have . Therefore, it suffices to show that achieves the value for the first time at a moment . Equation (5.18) yields
Using the fact that is a root of (4.10) and is a root of (5.6) as well as the definition of in (4.4) and the symmetry of , we have
Hence, the implicit function theorem in a neighborhood of together with (5.11) yield . □
Dynamics for as
Assume that case (3)(b) in the inductive step of the algorithm from Section 4.3 holds with some . If case (3)(a) holds, we omit this step and proceed with Section 5.4.
Dynamics for t close to . Three black curves are the graphs of and . The upper grey curve is the graph of the solution to Eq. (5.14). The lower (horizontal) grey curve is the graph of given by (5.13).
The following lemma defines a time moment satisfying (4.21).
There is a time momentsuch thatand
To follow the proof, we refer the reader to Fig. 3. We consider Eq. (5.14) with the initial data
First, note that for all . Indeed, if the equality is achieved for the first time at some moment , then . Moreover, since is non-increasing, we have at this moment. On the other hand, relations (5.14) and (5.15) imply that . Thus, the first inequality in (5.19) holds with replaced by any .
Furthermore, is non-increasing. Hence, for all by Lemma 5.3. It remains to find such that and .
Next, we introduce a positive function such that and
Consider the time moment
First, assume that
(the case will be considered at the end of the proof). Denote by the time moment preceding such that
Obviously, . Indeed, otherwise, and, due to (5.15) and Eq. (5.14), we would have .
Note that, due to (5.21), we have . Denote by
a moment such that
If , then there exists a desired time moment and the proof is complete.
Let us estimate the velocity for . For such t, we have and, due to (5.24), . Hence, (5.15) implies that for . Therefore,
Set
From it follows that for . If , then there exists a desired time moment and the proof is complete.
Assume that . It follows from (5.21) and (5.25) that
Furthermore, due to (5.15) and Eq. (5.14), we have
Therefore (see (5.26)),
Now inequality (5.27), relations (5.15) and Eq. (5.14) imply that at least as long as , where does not depend on t and D. Therefore, taking into account (5.28), we see that will achieve the value at a time moment . If follows from (5.23) and from the definitions of , , , and that . Hence, is the desired time moment.
Finally, if (5.22) is not valid, then and, as we already know, . The equality implies . If , then we can choose . If , then relations (5.14), (5.15) imply and all the argument following Eq. (5.23) can be repeated for . □
Dynamics for as
There is a time momentsuch thatforand.
Setting , we consider the problem (see Lemma 5.4)
where is given by (4.11). If , then we take and complete the proof.
Assume that . We need to prove that there exist positive functions and that tend to zero as and such that the solution to (5.14) satisfies
To construct the functions and , we consider positive sequences such that . Due to (4.13), (4.14) and (4.15), there exists a sequence such that
Then (5.29) and (5.30) imply that achieves the value at a time moment that satisfies
for all , where is a strictly decreasing sequence with . Therefore, where as . Now we set and for . □
Dynamics for as
To describe the dynamics for , we consider (5.8) with the initial data
(see Lemma 5.5). The following lemma will allow us to justify (5.1) (with replaced by n) and thus to complete the proof of Theorem 4.1.
There existsand() not depending on D such that the solutionto (5.8), (5.31) satisfiesforand.
Since for , it suffices to estimate the expression in brackets in (5.8). Using the definition of , the monotonicity of , and relations (5.31) and (4.8) (with replaced by n), we see that there exists such that
Equation (5.8) and inequality (5.32) imply that for . Obviously, we can choose independent of D such that . □
Finally, we determine the time moment (see (4.23)) at which the nth front becomes steady and immortal and its position as follows:
Due to Lemmas 5.5 and 5.6, and
This justifies (5.1) (with replaced by n). Moreover, since , the value satisfies (due to (5.8) and (5.9))
On the other hand, the definition (4.11) of , the definition (4.14) of and the definition (4.15) of imply that is the root of the equation
Therefore, , which implies . This completes the proof of Theorem 4.1.
An essential ingredient in the proof is the approximation (4.5) of the integral of the solution U by the error function . It is valid for t separated from zero and thus can be used for . In the case , the analog of Eq. (5.3) will be on the interval , where , and . On this time interval, one can use the approximation (see (3.7) and (3.8)) instead of (4.5).
Numerics
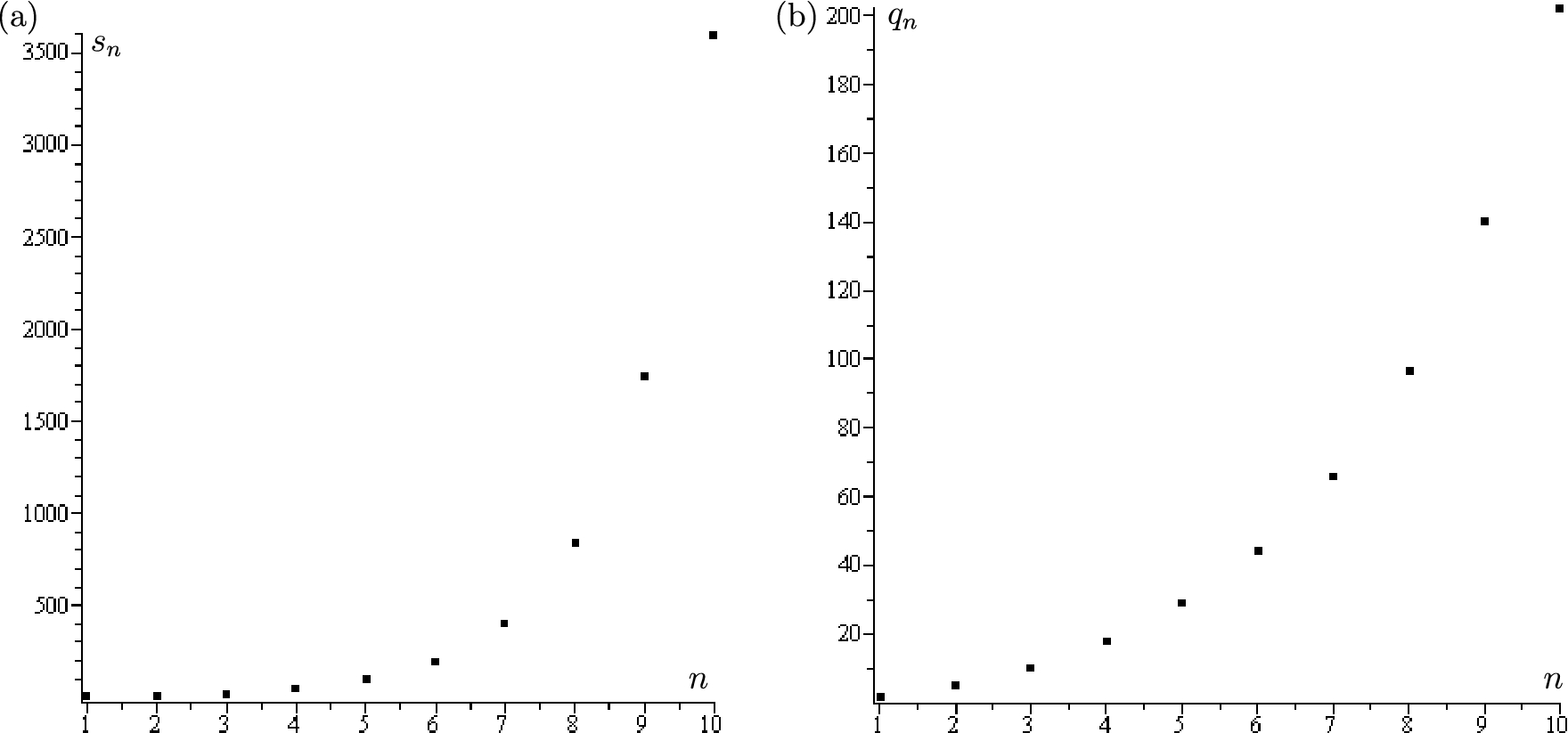
The two graphs indicate the values of (a) and (b) for from Theorem 4.1. The maximal and minimal values for admissible thresholds are and , respectively.
Figure 4 illustrates the values of and for from Theorem 4.1 found numerically. We note that for (i.e., case (3)(a) from the inductive step of the algorithm from Section 4.3 takes place) and for (i.e., case (3)(b) from the inductive step takes place). In general, it is an open question whether one of the two cases (3)(a) and (3)(b) takes place for all n, or both may fail for some n and hence the resulting sequences (4.6) are finite.
Footnotes
Acknowledgements
Pavel Gurevich acknowledges the support of the DFG through the Collaborative Research Center 910 and the Heisenberg fellowship. Dmitrii Rachinskii was supported by National Science Foundation, Grant DMS-1413223. The authors are grateful to Sergey Tikhomirov who created a software for a number of numerical experiments.
References
1.
M.Acar, A.Becskei and A.van Oudenaarden, Enhancement of cellular memory by reducing stochastic transitions, Nature435 (2005), 228–232.
2.
M.Acar, J.T.Mettetal and A.van Oudenaarden, Stochastic switching as a survival strategy in fluctuating environments, Nature Genetics40(4) (2008), 471–475.
3.
H.J.E.Beaumont, J.Gallie, C.Kost, G.C.Ferguson and P.B.Rainey, Experimental evolution of bet hedging, Nature462 (2009), 90–93.
4.
A.Becskei, B.Seraphin and L.Serrano, Positive feedback in eukaryotic gene networks: Cell differentiation by graded to binary response conversion, EMBO J.20 (2001), 2528–2535.
5.
S.Benzer, Induced synthesis of enzymes in bacteria analyzed at the cellular level, Biochim. Biophys. Acta11(3) (1953), 383–395.
6.
M.Brokate, N.D.Botkin and O.A.Pykhteev, Numerical simulation for a two-phase porous medium flow problem with rate independent hysteresis, Physica B: Condensed Matter407(9) (2012), 1336–1339.
7.
M.Brokate, A.Pokrovskii and D.Rachinskii, Asymptotic stability of continuum sets of periodic solutions to systems with hysteresis, J. Math. Anal. Appl.319 (2006), 94–109.
8.
M.Brokate and J.Sprekels, Hysteresis and Phase Transitions, Springer, 1996.
9.
C.Chiu, F.C.Hoppensteadt and W.Jäger, Analysis and computer simulation of accretion patterns in bacterial cultures, J. Math. Biol.32 (1994), 841–855.
10.
M.Cohn and K.Horbita, Inhibition by glucose of the induced synthesis of the beta-galactoside-enzyme system of Escherichia coli. Analysis of maintenance, J. Bacteriol.78 (1959), 601–612.
11.
M.Cohn and K.Horbita, Analysis of the differentiation and of the heterogeneity within a population of Escherichia coli undergoing induced beta-galactosidase synthesis, J. Bacteriol.78 (1959), 613–623.
12.
P.Colli, M.Grasselli and J.Sprekels, Automatic control via thermostats of a hyperbolic Stefan problem with memory, Appl. Math. Optim.39 (1999), 229–255.
13.
R.Cross, H.McNamara, A.Pokrovskii and D.Rachinskii, A new paradigm for modelling hysteresis in macroeconomic flows, Physica B403(2,3) (2008), 231–236.
14.
M.Delbrück, Discussion, in: Unités Biologiques Douées de Continuité Génétique, Editions du Centre National de la Recherche Scientifique, Paris, 1949, pp. 33–35.
15.
D.Dubnau and R.Losick, Bistability in bacteria, Mol. Microbiol.61 (2006), 564–572.
16.
A.Friedman and L.-S.Jiang, Periodic solutions for a thermostat control problem, Commun. Partial Differ. Equ.13(5) (1988), 515–550.
17.
G.Friedman, P.Gurevich, S.McCarthy and D.Rachinskii, Switching behaviour of two-phenotype bacteria in varying environment, J. Phys.: Conf. Ser.585 (2015), 0121012.
18.
G.Friedman, S.McCarthy and D.Rachinskii, Hysteresis can grant fitness in stochastically varying environment, PLoS ONE9(7) (2014), e103241.
19.
T.S.Gardner, C.R.Cantor and J.J.Collins, Construction of a genetic toggle switch in Escherichia coli, Nature403 (2000), 339–342.
20.
P.Gurevich and D.Rachinskii, Well-posedness of parabolic equations containing hysteresis with diffusive thresholds, Tr. Mat. Inst. Steklova283 (2013), 92–114; English transl.: Proc. Steklov Inst. Math.283 (2013), 87–109.
21.
P.Gurevich and D.Rachinskii, Pattern formation in parabolic equations containing hysteresis with diffusive thresholds, J. Math. Anal. Appl.424 (2015), 1103–1124.
22.
P.Gurevich and S.Tikhomirov, Uniqueness of transverse solutions for reaction–diffusion equations with spatially distributed hysteresis, Nonlinear Anal. A: Theory, Methods and Applications75 (2012), 6610–6619.
23.
P.Gurevich and S.Tikhomirov, Systems of reaction–diffusion equations with spatially distributed hysteresis, Mathematica Bohemica139(2) (2014), 239–257.
24.
P.L.Gurevich, Periodic solutions of parabolic problems with hysteresis on the boundary, Discrete Cont. Dynam. Systems A29(3) (2011), 1041–1083.
25.
P.L.Gurevich and S.B.Tikhomirov, Symmetric periodic solutions of parabolic problems with discontinuous hysteresis, J. Dynamics and Differential Equations23(4) (2011), 923–960.
26.
F.C.Hoppensteadt and W.Jäger, Pattern formation by bacteria, in: Biological Growth and Spread, Lecture Notes in Biomathematics, Vol. 38, Springer, 1980, pp. 68–81.
27.
F.C.Hoppensteadt, W.Jäger and C.Pöppe, A hysteresis model for bacterial growth patterns, in: Modelling of Patterns in Space and Time, Lecture Notes in Biomathematics, Vol. 55, Springer, 1984, pp. 123–134.
28.
B.B.Kaufmann, Q.Yang, J.T.Mettetal and A.van Oudenaarden, Heritable stochastic switching revealed by single-cell genealogy, PLoS Biol.5(9) (2007), e239.
29.
J.Kopfová, Hysteresis in biological models, J. Phys.: Conf. Ser.55 (2006), 130–134.
30.
J.Kopfová and T.Aiki, A mathematical model for bacterial growth, in: Recent Advances in Nonlinear Analysis, Proceedings of the International Conference on Nonlinear Analysis, World Scientific, 2008, pp. 1–10.
31.
A.Krasnosel’skii and D.Rachinskii, On continua of cycles in systems with hysteresis, Doklady Math.63(3) (2001), 339–344.
32.
A.Krasnosel’skii and D.Rachinskii, On a bifurcation governed by hysteresis nonlinearity, NoDEA Nonlinear Differential Equations Appl.9 (2002), 93–115.
33.
P.Krejčí, Hysteresis, Convexity and Dissipation in Hyperbolic Equations, GAKUTO International Series: Mathematical Sciences and Applications, Vol. 8, Gakkotosho, Tokyo, 1996.
34.
P.Krejci, P.O’Kane, A.Pokrovskii and D.Rachinskii, Stability results for a soil model with singular hysteretic hydrology, J. Phys.: Conf. Ser.268(1) (2011), 012016.
35.
P.Krejci, P.O’Kane, A.Pokrovskii and D.Rachinskii, Properties of solutions to a class of differential models incorporating Preisach hysteresis operator, Physica D241 (2012), 2010–2028.
36.
E.Kussell and S.Lieber, Phenotypic diversity, population growth, and information in fluctuating environments, Science309 (2005), 2075–2078.
37.
M.Mimura, H.Sakaguchi and M.Matsushita, Reaction–diffusion modelling of bacterial colony patterns, Physica A282 (2000), 283–303.
38.
H.D.Møller, K.S.Andersen and B.Regenberg, A model for generating several adaptive phenotypes from a single genetic event Saccharomyces cerevisiae GAP1 as a potential bet-hedging switch, Commun. Integr. Biol.6(3) (2013), e23933.
39.
J.Monod, From enzymatic adaptation to allosteric transitions, Science154 (1966), 475–483.
40.
A.Novick and M.Weiner, Enzyme induction as an all-or-none phenomenon, Proc. Natl. Acad. Sci. USA43 (1957), 553–566.
41.
E.M.Ozbudak, M.Thattai, H.N.Lim, B.I.Shraiman and A.van Oudenaarden, Multistability in the lactose utilization network of Escherichia coli, Nature427 (2004), 737–740.
42.
V.R.Pannala, P.J.Bhat, S.Bhartiya and K.V.Venkatesh, Systems biology of GAL regulon in Saccharomyces cerevisiae, WIREs Syst. Biol. Med.2(1) (2010), 98–106.
43.
A.Pimenov, T.Kelly, A.Korobeinikov, M.J.A.O’Callaghan, A.Pokrovskii and D.Rachinskii, Memory effects in population dynamics: Spread of infectious disease as a case study, Mathematical Modelling of Natural Phenomena7(1) (2012), 1–30.
44.
A.Pimenov and D.Rachinskii, Linear stability analysis of systems with Preisach memory, Discrete Cont. Dynam. Systems B11(4) (2009), 997–1018.
45.
J.R.Pomerening, E.D.Sontag and J.E.FerrellJr., Building a cell cycle oscillator: Hysteresis and bistability in the activation of Cdc2, Nature Cell Biol.5 (2003), 346–351.
46.
D.Rachinskii, Asymptotic stability of large-amplitude oscillations in systems with hysteresis, NoDEA Nonlinear Differential Equations Appl.6(3) (1999), 267–288.
47.
S.Spiegelman and W.F.DeLorenzo, Substrate stabilization of enzyme-forming capacity during the segregation of a heterozygote, Proc. Natl. Acad. Sci. USA38(7) (1952), 583–592.
48.
M.Thattai and A.van Oudenaarden, Stochastic gene expression in fluctuating environments, Genetics167 (2004), 523–530.
49.
A.Visintin, Evolution problems with hysteresis in the source term, SIAM J. Math. Anal.17 (1986), 1113–1138.
50.
A.Visintin, Differential Models of Hysteresis, Springer, Berlin, Heidelberg, 1994.
51.
Ö.Winge and C.Roberts, Inheritance of enzymatic characters in yeasts, and the phenomenon of long-term adaptation, Compt. Rend. Lab. Carlsberg, Sér. Physiol.24 (1948), 263–315.