
Abstract
Multidrug resistance (MDR) caused by human ABCB1 (P-glycoprotein/MDR1) is one of the major obstacles in chemotherapy. To understand the mechanism of MDR by ABCB1 and circumvent the MDR, in the present study, we established human ABCB1-expressing cells (Flp-In-293/ABCB1 cells) and examined the cytotoxic effects of four guanidine alkaloids from Pterogyne nitens (galegine, nitensidine A, pterogynidine and pterogynine) using Flp-In-293/Mock and Flp-In-293/ABCB1 cells. The activity of ABCB1 in Flp-In-293/ABCB1 cells were confirmed by typical substrates for ABCB1 (taxol and vinblastine) in MTT assay. Flp-In-293/ABCB1 cells were also resistant to the four guanidine alkaloids as well as taxol and vinblastine compared to Flp-In-293/Mock cells although the four guanidine alkaloids exhibited cytotoxicity against the two Flp-In-293 cells. Furthermore, the four guanidine alkaloids were also found to stimulate the ATPase activity of ABCB1 in ATPase assays. These results suggest that ABCB1 can confer the resistance to the cytotoxic guanidine alkaloids by transporting them.
Background
Human ABCB1 (P-glycoprotein/MDR1) is a transmembrane efflux pump that belongs to the ATP-binding cassette (ABC) transporter superfamily. It has been cloned as the multidrug resistance 1 (MDR1) gene from cancer cells exerting cross-resistance to several structurally diverse and mechanistically unrelated anti-cancer drugs [1–4]. As a drug transporter, ABCB1 regulates the intracellular concentration of various substrates by extruding them from cells. Overexpression of ABCB1 reportedly confers cancer cells with resistance to anti-cancer drugs such as doxorubicin, vincristine and paclitaxel [5–8]. Thus, the enzymatic activity of ABCB1 can contribute to multidrug resistance (MDR) in cancer cells, and can be an obstacle to successful cancer chemotherapy. Inhibitors, modulators and chemosensitizers of ABCB1 have been screened extensively to circumvent MDR. However, the structural features important for the inhibition, modulation and chemosensitization of ABCB1 are incompletely understood. Thus, screening for inhibitors, modulators and chemosensitizers of ABCB1 with novel structures, and subsequent structure–activity relationship analyses of these structures, would be helpful for understanding the detailed mechanisms by which ABCB1 recognizes and transports its substrates.
Objective
The ultimate goal of this work is to discover ways to circumvent ABCB1-mediated MDR. In the present study, we provide evidence that ABCB1-expressing cells can exert resistance to cytotoxic guanidine alkaloids isolated from a plant species found in Brazil: Pterogyne nitens.
Materials and methods
Guanidine alkaloids
The guanidine alkaloids tested in the present study were obtained from Pterogyne nitens leaves or branches; their isolation and identification have been previously described [9,10].
Cell culture
CCRF-CEM and CEM/ADR5000 cells were kindly provided by Dr. Axel Sauerbrey (Department of Pediatrics, University of Jena, Jena, Germany). CEM/ADR5000 cells are an ABCB1 overexpressing, multidrug-resistant subline of CCRF-CEM cells. All cells were maintained in RPMI 1640 medium (Biochrom AG, Berlin, Germany) supplemented with 10% (v/v) fetal bovine serum (FBS) (ICN Biomedicals, Aurora, OH, USA), 0.8 mM L-glutamine, 50 µg/ml kanamycin, 100 U/ml penicillin and 100 µg/ml streptomycin at 37°C in a humidified atmosphere of 5% (v/v) CO2 in air. CEM/ADR5000 cells were cultured with 2.5 µg/ml doxorubicin for 24 h once a week to maintain ABCB1 expression in the cells.
We prepared Flp-In-293/ABCB1 cells from Flp-In-293 cells (Invitrogen, Carlsbad, CA, USA) having the Flp recombination target (FRT) site at the telomeric region of only one of the pair of chromosomes 12 [11,12] to examine the effect of ABCB1 on the cytotoxicity of guanidine alkaloids. For this purpose, Flp-In-293 cells were transfected with the pcDNA5/FRT/ABCB1 vector, the Flp recombinase expression plasmid pOG44 [13–15], and Lipofectamine™-2000 (Invitrogen) according to manufacturer instructions. Single colonies resistant to hygromycin B (Invitrogen) were selected and subcultured. Selection of positive colonies was undertaken by immunoblotting with ABCB1-specific monoclonal antibody C219 (EMD Biosciences, Darmstadt, Germany). Mock (Flp-In-293/Mock) cells used in the present study had been established in our previous studies [11]. Parental Flp-In-293 cells were maintained in Dulbecco’s Modified Eagle’s Medium (DMEM) supplemented with 10% (v/v) heat-inactivated FBS, 2 mM L-glutamine, 100 U/ml penicillin, 100 µg/ml streptomycin, 250 ng/ml amphotericin B and 100 µg/ml Zeocin™ at 37°C in a humidified atmosphere of 5% (v/v) CO2 in air. Flp-In-293/ABCB1 and Flp-In-293/Mock cells were maintained in DMEM supplemented with the agents shown above except that 100 µg/ml hygromycin B was used instead of Zeocin.
2,3-bis-(2-methoxy-4-nitro-5-sulfophenyl)-2H-tetrazolium-5-carboxanilide (XTT) assay
CCRF-CEM and CEM/ADR5000 cells were seeded (2 × 104 cells/well) into 96-well culture plates and cultured with compounds for 72 h. After incubation, cells were treated with XTT for 5 h using Cell Proliferation Kit II (XTT assay) (Roche, Mannheim, Germany) according to manufacturer protocols. Absorbance at 490 nm and 655 nm was measured as test and reference wavelengths, respectively. Data are the mean ± SEM of two experiments.
3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide (MTT) assay
Flp-In-293/Mock and Flp-In-293/ABCB1 cells were seeded (5 × 103 cells/well) into 96-well culture plates and pre-cultured for 24 h. After pre-culture, compounds were added into the cell cultures and cells were incubated for 72 h. After incubation, cells were treated with 500 µg/ml MTT (Sigma-Aldrich, St. Louis, MO, USA) for 3 h. And then 20% (w/v) sodium dodecyl sulfate in phosphate-buffered saline (100 µl/well) was added to solubilize MTT-formazan, followed by overnight incubation. After the incubation, absorbance at 570 nm and 630 nm was measured as a test and reference wavelengths, respectively. Data are the mean ± SEM of two experiments.
ATPase assay
The ATPase activities of ABCB1 in the presence of various test compounds were measured in 96-well plates by using plasma membrane fractions prepared from ABCB1-expressing Sf9 cells [16]. In brief, cell membranes (1 µg of protein per well) were suspended in 10 µl of the incubation medium containing 50 mM Tris-Mes (pH 6.8), 2 mM EGTA, 2 mM dithiothreitol, 50 mM potassium chloride, 5 mM sodium azide and 2 mM ouabain. This medium was mixed with 10 µl of a test compound solution in 96-well plates. The ATPase reaction was started by adding 20 µl of 4 mM ATP/Mg solution to the reaction mixture in the plates, and the incubation was maintained at 37°C for 30 min. The reaction was stopped by the addition of 20 µl of 5% trichloroacetic acid followed by 42 µl of solution A (2 N hydrochloric acid: 0.1 M sodium molybdate = 4 : 3) and 18 µl of solution B (0.084% (w/v) malachite green in 1% (w/v) polyvinyl alcohol). Thereafter, 120 µl of solution C (7.8% (v/v) sulfuric acid) was added to the mixture in the well plates. After one hour of incubation at room temperature, the absorbance of the reaction mixture in each well was measured at a wavelength of 655 nm by using Bio-Rad Model 680 Absorbance Microplate Reader (Bio-Rad Laboratories, Inc.). The amount of liberated phosphate was quantified on the basis of the calibration line established with inorganic phosphate standards.
Statistical analyses
The results were statistically analyzed using Microsoft Excel 2010 software (Microsoft, Redmond, WA, USA). The statistical significance of differences was determined using the Student’s t-test. Structures of guanidine alkaloids.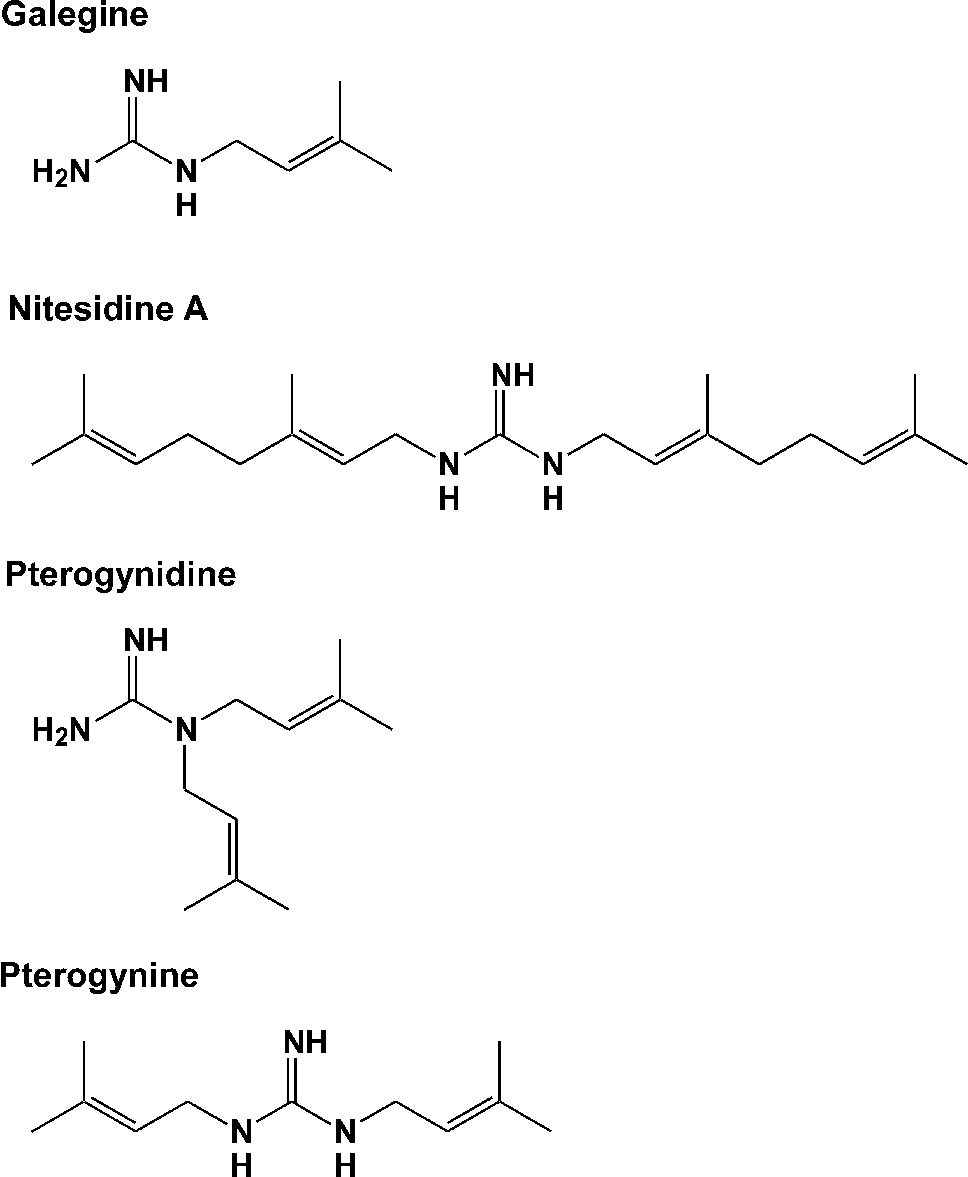
Cytotoxicity of guanidine alkaloids against CCRF-CEM, CEM/ADR5000, Flp-In-293/Mock, Flp-In-293/ABCB1 cells
Note: Data are expressed as the means ± SEM of triplicate determinations.
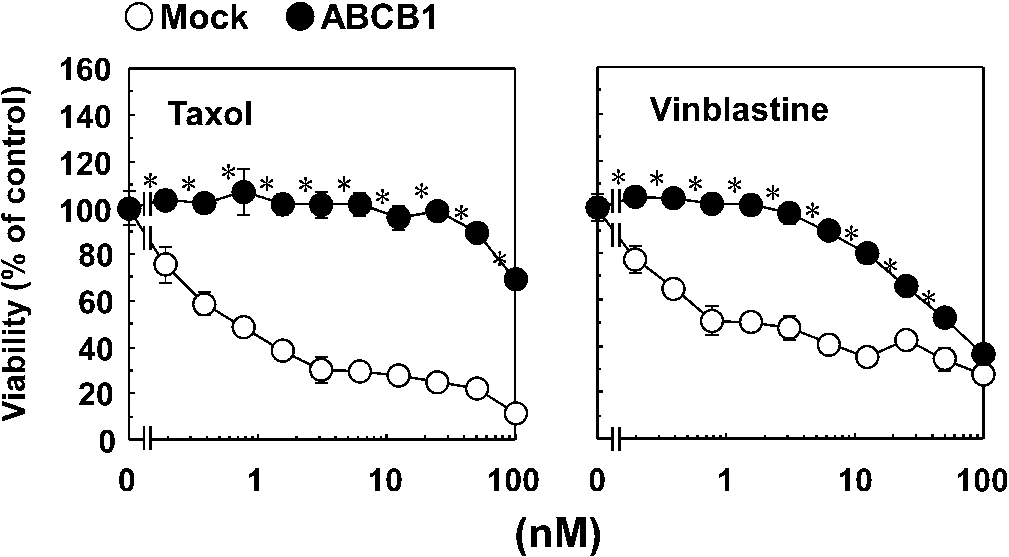
Drug resistance ability of Flp-In-293/ABCB1 cells. Flp-In-293/Mock (open circles) and Flp-In-293/ABCB1 cells (closed circles) were cultured with or without the taxol and vinblastine for 72 h. After the culture, viability of the cells was determined by MTT assay as described in Section 3. Similar results were obtained in more than two other experiments. Data are expressed as the mean ± SEM of triplicate determinations. *
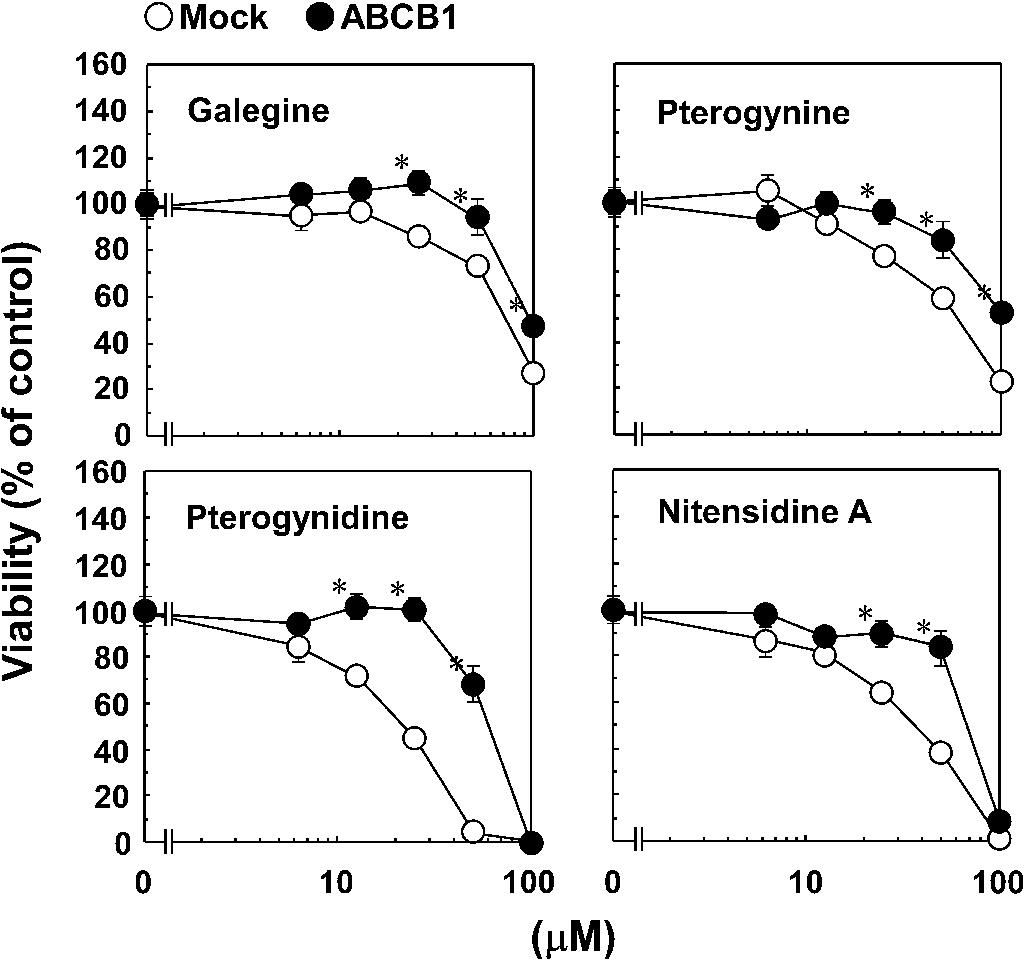
Effect of the guanidine alkaloids on the viability of Flp-In-293/Mock and Flp-In-293/ABCB1 cells. Flp-In-293/Mock (open circles) and Flp-In-293/ABCB1 cells (closed circles) were cultured with or without the guanidine alkaloids for 72 h. After the culture, viability of the cells was determined by MTT assay as described in Section 3. Similar results were obtained in more than two other experiments. Data are expressed as the mean ± SEM of triplicate determinations. *
We evaluated the cytotoxic effects of four guanidine alkaloids isolated from Pterogyne nitens on CCRF-CEM and its MDR counterpart (CEM/ADR5000 cells) (Fig. 1 and Table 1). CCRF-CEM and CEM/ADR5000 cells were cultured with or without guanidine alkaloids for 72 h. After the culture, cell viability was determined using the XTT assay. All four guanidine alkaloids exhibited toxicity against CCRF-CEM cells as shown in Table 1. The order of cytotoxic activity was nitensidine A > pterogynidine > pterogynine > galegine. Compared with CCRF-CEM cells, CEM/ADR5000 cells were resistant to the toxic effects of all four guanidine alkaloids. To elucidate the involvement of ABCB1 in the resistance of CEM/ADR5000 cells against the four guanidine alkaloids, the cytotoxic effects of the four compounds were examined using Flp-In-293/ABCB1 cells stably expressing wild-type human ABCB1 and its counterpart Flp-In-293/Mock cells. For this purpose, Flp-In-293/ABCB1 cells were established and their drug-resistance activities were examined using taxol and vinblastine (typical substrates for ABCB1). Flp-In-293/Mock (open circles in Fig. 2) and Flp-In-293/ABCB1 cells (closed circles in Fig. 2) were cultured with or without the typical ABCB1 substrates for 72 h. After the culture, cell viability was determined by the MTT assay. Compared with Flp-In-293/Mock cells, Flp-In-293/ABCB1 cells showed resistance to both ABCB1 substrates (Fig. 2). Flp-In-293/Mock (open circles in Fig. 2) and Flp-In-293/ABCB1 cells (closed circles in Fig. 3) were also cultured with or without the guanidine alkaloids for 72 h. All four guanidine alkaloids showed toxicity against Flp-In-293/Mock cells (Fig. 3). The order of cytotoxic activity was pterogynidine > nitensidine A > pterogynine > galegine. As well as CEM/ADR5000 cells, Flp-In-293/ABCB1 cells were resistant to four guanidine alkaloids compared with Flp-In-293/Mock cells.
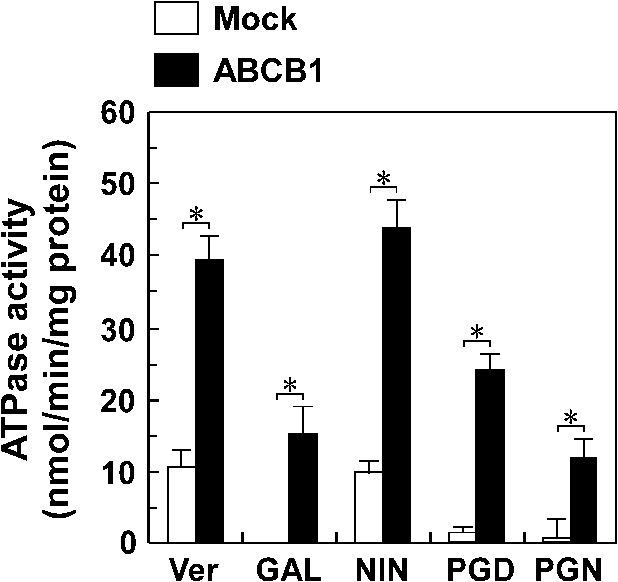
Effect of the guanidine alkaloids on ATPase activity of ABCB1. The ATPase activity in the plasma membrane from Sf9 cells expressing ABCB1 (closed bars) or not expressing ABCB1 (open bars) was measured in the presence of 10 µM guanidine alkaloids as described in Section 3. Similar results were obtained in more than two other experiments. Data is expressed as the mean ± SEM of triplicate determinations. Ver – verapamil, GAL – galegine, NIN – nitensidine A, PGD – pterogynidine, PGN – pterogynine. *
As shown in Fig. 4, four guanidine alkaloids were each found to cause an ABCB1-dependent hydrolysis of ATP when incubated with plasma membranes from Sf9 cells expressing wild-type ABCB1. Furthermore, the hydrolysis levels of ATP by nitensidine A were comparable to those by verapamil. These results suggest that the guanidine alkaloids were recognized as substrates by ABCB1.
We carried out XTT and MTT assays to investigate the cytotoxic effects of guanidine alkaloids on cells expressing human ABCB1 and their counterpart cells. Guanidine alkaloids have been reported to be cytotoxic against the human infiltrating ductal carcinoma ZR-7531 cell line but not against human myeloblastic leukemia HL-60, human colon carcinoma HCT-8, human melanoma MDA-MB-435 and human glioblastoma SF-295 cells [9,10]. In the present study, four guanidine alkaloids were found to be cytotoxic against four cell lines (CCRF-CEM, CEM/ADR5000, Flp-In-293/Mock and Flp-In-293/ABCB1 cells). Although the current number of the compounds is not sufficient for structure–activity relationship analysis, based on the structural differences among the four guanidine alkaloids, the number, length and binding site of aliphatic side chains of the four guanidine alkaloids seem to determine their levels of cytotoxicity.
The sensitivities of CCR-CEM and Flp-In-293 cells to the four guanidine alkaloids were slightly different; CCR-CEM and Flp-In-293 cells were the most sensitive to nitensidine A and pterogynidine, respectively. As well as the chemical features of the four guanidine alkaloids, the present study clearly showed that the cytotoxic activities of the four compounds were dependent upon ABCB1 expression. In the XTT and MTT assays, cells expressing ABCB1 were resistant to all of the guanidine alkaloids tested. These results suggest that ABCB1 confers the cells resistance to the four guanidine alkaloids. A study to ascertain if the four guanidine alkaloids are substrates for ABCB1 is in progress although they were shown to stimulate ATPase activity of ABCB1 in Fig. 4.
In the present study, ABCB1-expressing cells showed resistance to the four cytotoxic guanidine alkaloids, suggesting that several guanidine alkaloids can serve as substrates for ABCB1. Among the guanidine alkaloid test compounds, pterogynidine and pterogynine (which both have a molecular weight of 195) may be the smallest molecules transported by ABCB1 because these guanidine alkaloids are smaller than phenytoin (molecular weight = 252), which was reported to be the smallest substrate for ABCB1 by Schinkel et al. [17]. Further studies are required to circumvent MDR, but the present study has shed light on the molecular mechanisms by which ABCB1 recognizes its substrates.
Footnotes
Acknowledgements
This study was supported by the Grant-in-Aid for Scientific Research (C) (JSPS KAKENHI Grant Number 24592822) from the Ministry of Education, Culture, Sports, Science and Technology (MEXT) and intramural grants from Chubu University (Grant A (23IM04A) and B (23IM02B and 24M01B)) and the German Cancer Research Center (Heidelberg, Germany). The phytochemical studies were supported by a grant from the Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP) as part of the Biota-FAPESP-Biodiversity Virtual Institute Program (![]() ), grant number 03/02176-7, awarded to V.S.B. We thank Karen Duffy (Cornell University, Ithaca, NY, USA) for her critique of the manuscript.
), grant number 03/02176-7, awarded to V.S.B. We thank Karen Duffy (Cornell University, Ithaca, NY, USA) for her critique of the manuscript.