
Abstract
Background:
Curcumin possesses significant anti-atherosclerosis properties. Lipocalin-2 (LCN2) is already known as one of the most promising biomarkers of atherosclerosis. However, research on the effect of curcumin on regulating LCN2 expression in atherogenesis is very limited. The aim of the study was to investigate whether curcumin could alleviate atherosclerosis in ApoE−/− mice by down-regulating LCN2 expression.
Methods:
Fifty apolipoprotein E knockout (ApoE−/−) mice were fed with a western diet for 12 weeks and randomly divided into five groups: model group (Mod), positive control group (Lov, with 30 mg/kg/d lovastatin), three curcumin groups (CurL, CurM and CurH, with 40, 60 and 80 mg/kg/d curcumin), while 10 C57BL/6J mice were fed with a standard mouse chow diet as a control group (Con). LCN2 in serum and aorta, biomarkers of serum lipid and inflammation were determined and the plaque area of the atherosclerosis lesions was quantified.
Results:
CurM and CurH group were significantly lower than Mod group in terms of serum LCN2, lipid and inflammatory cytokine levels, plaque area and LCN2 expression in atherosclerotic lesions, similar to lovastatin treatment.
Conclusions:
Our findings indicate that dietary curcumin ameliorates western diet-induced atherosclerosis in ApoE−/− mice, which is related to LCN2 down-regulation, anti-hyperlipidemia effect as well as the inhibition of inflammation.
Introduction
Atherosclerosis is a complex chronic inflammatory and metabolic disease, which is a major contributor of morbidity and mortality in the world. In addition to arterial lipid accumulation and lipid dysfunction, adipose tissue inflammation and perturbation of adipokine secretion have been increasingly recognized as essential reasons in the pathogenesis of atherosclerosis. Lipocalin-2 (LCN2), also termed neutrophil gelatinase-associated lipocalin (NGAL) or siderocalin, mainly released from adipocytes, is a small 25 kDa protein which was first discovered in human neutrophils [1,2]. LCN2 originally identified as an inflammatory factor released by activated neutrophils, is also expressed in hepatocytes and various human epithelial tissue, such as trachea, colon, stomach and kidney in response to various inflammatory signals and different types of injury [3]. Recently LCN2 has emerged as one of the most reliable and attractive biomarkers for the diagnosis and risks tratification of patients suffering from cardiovascular diseases (CVD) [4,5]. Elevated serum LCN2 level is an independent predictor of CVD events in men in a population-based cohort [6]. Recent data also indicated that LCN2 is closely related to atherogenesis, the underlying pathology for CVD. Patients with subclinical atherosclerosis which is assessed by intima-media thickness (IMT) at carotid and femoral arteries with ultrasound showed significantly higher circulating concentrations of LCN2 compared with those without, besides, serum LCN2 level was positively correlated with carotid IMT [7]. In addition to its circulating level, enhanced LCN2 content has been detected in unstable human atherosclerotic plaque when intra plaque hemorrhage or luminal thrombus is present [8]. Based on these pieces of evidence, we hypothesized that agents suppressing LCN2 expression would be potential therapeutic agents that prevent against atherosclerosis.
Curcumin, the active constituent in the dietary spice turmeric (Curcuma longa Linn), is a pharmacologically effective and safe agent that plays a key role in anti-atherosclerosis processes. Curcumin may prevent the formation of atherosclerotic lesions in rabbits by attenuating adhesion molecules and matrix metalloproteinase (MMPs) expression [9]. Curcumin modulated macrophage polarization and inhibited inflammatory cytokines secretion through the inhibition of the toll-like receptor 4 expression and its signaling pathway [10]. The anti-atherogenic property of curcumin also could be linked to its effect on gene networks and cell functions related to leukocyte adhesion and transendothelial migration via nuclear factor-κB pathways [11]. Although beneficial effects of curcumin on atherosclerosis have been suggested, the underlying mechanisms responsible for the amelioration of atherosclerosis have not been fully elucidated. Whether curcumin could ameliorate the development of atherosclerosis by down-regulating LCN2 expression is currently unknown.
On the basis of these findings, we hypothesized that curcumin could prevent against atherogenesis by down-regulating LCN2 expression. Apolipoprotein E knockout (ApoE−/−) mice is a genetically modified strain of mice that is commonly used as the animal model for spontaneous atherosclerosis. Therefore, we evaluated the effect of curcumin on high fat diet-induced atherogenesis in ApoE−/− mice and investigated the mechanisms underlying curcumin-mediated modulation of atherosclerosis.
Materials and methods
Animals and diets
Fifty male 6-week-old ApoE−/− mice with a genetic C57BL/6J background and 10 male 6-week-old C57BL/6J mice were purchased from Vital River Experimental Animal Technology Co. Ltd (Beijing, China) and housed in SPF grade Experimental Animal House at Jiangxi University of Traditional Chinese Medicine (Jiangxi, China) in environmentally controlled conditions (
Biochemical tests of serum
All mice were sacrificed by collecting whole blood via the abdominal aorta under ether euthanasia at the last day of experiment after 12-h fasting. Serum were isolated from blood by centrifuge and were stored at −80°C until required for analysis. Serum levels of total cholesterol (TC), triglycerids (TG), high density lipoprotein-cholesterol (HDL-C) and low density lipoprotein-cholesterol (LDL-C) were assayed using commercially available kits (Invitrogen, USA). The circulating levels of serum LCN2, interleukin-6 (IL-6), tumor necrosis factor-alpha (TNF-α) and high-sensitivity C-reactive protein (hs-CRP) were measured by enzyme-linked immunosorbent assay (ELISA) according to the manufacturer’s protocols of ELISA kits (eBioscience, USA).
Histologic analysis
Right atrium was incised and the heart was perfused by PBS (10 mM,
Immunohistochemistry
After antigen retrieval by boiling in 0.01 M sodium citrate for 10 min, deparaffinized sections were quenched in 0.3% hydrogen peroxide for 30 min, followed by incubated in 1% BSA in PBS for 30 min. Sections were labeled with goat anti-mouse LCN2 antibody (Santa Cruz, USA, 1:200 dilution) at 37°C for 45 min and then overnight at 4°C. After washing, the bound antibodies were conjugated with secondary antibodies at 37°C for 1 h, and then the DAB substrate was administrated and incubated for 1 min. The sections were counterstained with hematoxylin and the result was acquired with Image-Pro Plus 5.0 Analysis Software (Media Cybernetics, USA).
Western blotting
The mice aortas were homogenized in lysis buffer containing 1% NP-40, 50 mM Tris (
Quantitative real-time PCR (qRT-PCR)
Total RNA was extracted from the treated mice descending aortas using RNeasy Plus Micro Kit (Qiagen, Germany) according to the operating instruction. The first-strand cDNA was synthesized from the total RNA to establish a reverse transcription system by PrimeScript™ RT Master Mix Sample kit (Takara, Japan). Amplified DNA was separated on 1.5% agarose gels and visualized under UV light. qRT-PCR was performed using an Applied Biosystems Prism sequence detection system (Applied Biosystems, USA) according to the manufacturer’s protocols. Primer sequences for qRT-PCR were: LCN2: forward 5′-TGACAGCCCACCTCCCT-3′ and reverse 5′-CCACTTCCATCCTGGATGGT-3′; β-actin, forward 5′-CAAGAGATACGGCCGGCTGCT-3′ and reverse 5′-TCCTTCTGCTGCATCTCGGCA-3′.
Statistical analysis
Data were expressed as means ± SD. Statistical analyses were performed using one-way ANOVA and a value of
Results
Curcumin decreased serum lipid profiles
To determine whether curcumin altered the serum lipid profiles, we measured serum TC, TG, LDL-C and HDL-C levels in each group at the conclusion of the study (Table 1). Comparing with Con group, serum levels of TC, TG and LDL-C in Mod group were significantly increased, indicating occurrence of hyperlipidemia. After treatments of lovastatin or curcumin, serum lipid profiles were remarkably improved. TC, TG and LDL-C levels in Lov, CurM and CurH groups were significant reduced compared to Mod group, while curcumin supplementation did not affect HDL-C levels compared to Mod group.
Effects of curcumin on serum lipid profiles in mice (
, mmol/L)
Effects of curcumin on serum lipid profiles in mice (
Values are
Con, C57BL/6J mice were fed with a standard mouse chow diet; Mod, ApoE−/− mice were fed with a western diet; Lov, ApoE−/− mice gavaged with lovastatin (30 mg/kg) were fed with a western diet; CurL, ApoE−/− mice gavaged with curcumin (40 mg/kg) were fed with a western diet; CurM, ApoE−/− mice gavaged with curcumin (60 mg/kg) were fed with a western diet; CurH, ApoE−/− mice gavaged with curcumin (80 mg/kg) were fed with a western diet.
In order to evaluate whether curcumin altered the serum levels of inflammatory cytokines, serum IL-6, TNF-α and hs-CRP levels were detected by ELISA (Table 2). Western diet fed in Mod group had remarkably higher serum IL-6, TNF-α and hs-CRP levels than those of Con group, whereas administration with lovastatin significantly decreased the alterations compared with the Mod group. Curcumin in CurM and CurH group showed a similar trend of lovastatin effect on serum levels of IL-6, TNF-α and hs-CRP.
Effects of curcumin on serum levels of LCN2, IL-6, TNF-α and hs-CRP in mice (
)
Effects of curcumin on serum levels of LCN2, IL-6, TNF-α and hs-CRP in mice (
Values are
Con, C57BL/6J mice were fed with a standard mouse chow diet; Mod, ApoE−/− mice were fed with a western diet; Lov, ApoE−/− mice gavaged with lovastatin (30 mg/kg) were fed with a western diet; CurL, ApoE−/− mice gavaged with curcumin (40 mg/kg) were fed with a western diet; CurM, ApoE−/− mice gavaged with curcumin (60 mg/kg) were fed with a western diet; CurH, ApoE−/− mice gavaged with curcumin (80 mg/kg) were fed with a western diet.
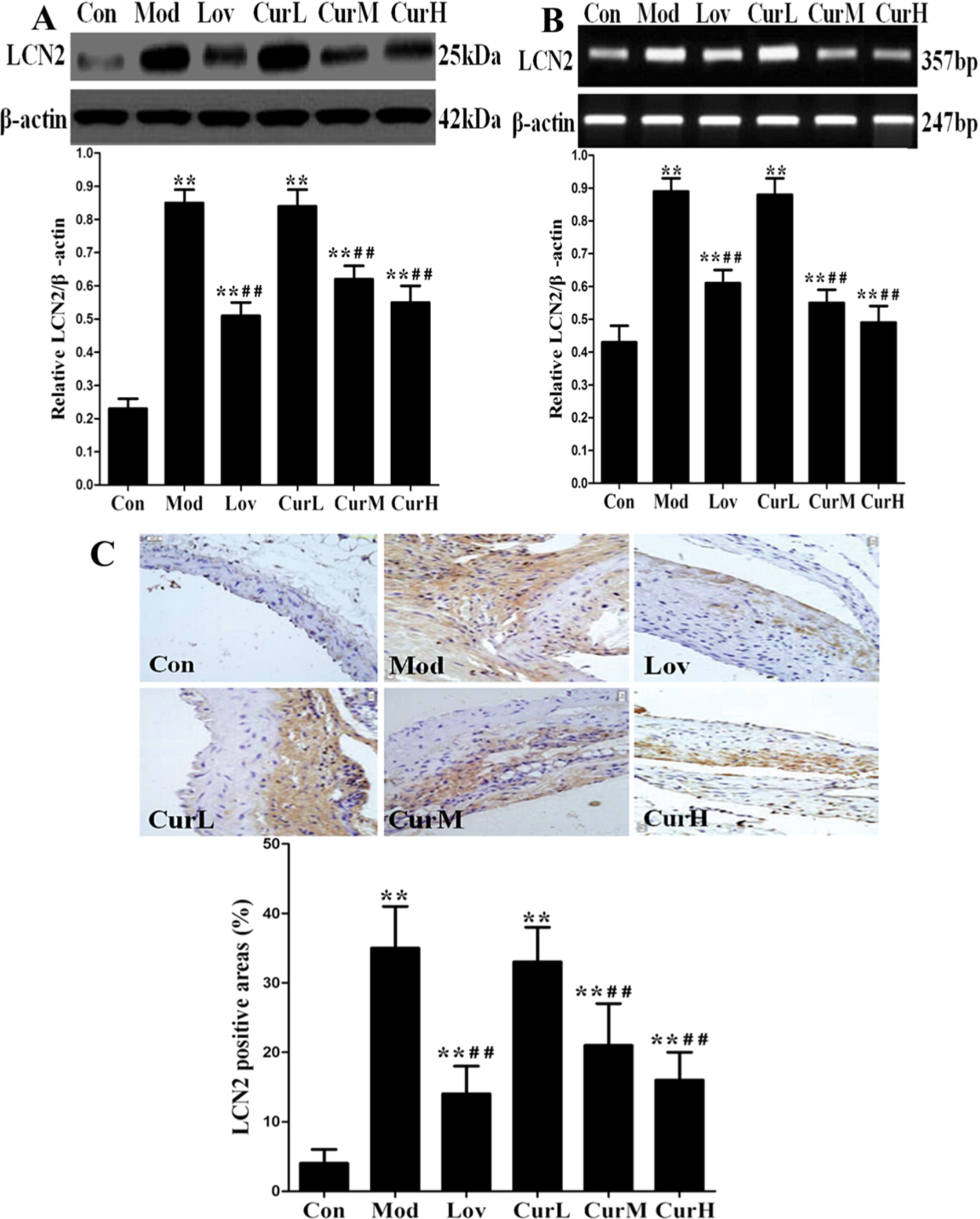
Effect of curcumin on LCN2 expression in ApoE−/− mice fed a western diet for 12 weeks. (A) The protein expression of LCN2 in mice aorta. (B) The mRNA expression of LCN2 in mice aorta. (C) Representative images of immunohistochemistry for LCN2 in mice aorta (400×). Values are
We measured serum LCN2 level, LCN2 protein and mRNA expression in the aorta in each group by ELISA, Western blotting and qRT-PCR assay, respectively. Serum LCN2 level (Table 2), LCN2 protein (Fig. 1(A)) and mRNA expression (Fig. 1(B)) in the aorta in Mod group were much higher than in Con group. Serum LCN2 level, LCN2 protein and mRNA expression in Lov, CurM and CurH groups were significant reduced compared to Mod group. To identify the distribution of LCN2 in mice aorta, we determined its level by immunohistochemistry assay. Immunohistochemical staining result (Fig. 1(C)) showed that significantly more LCN2 was detected from Mod group compared with Con group, and less LCN2 were detected from Lov, CurM and CurH groups compared with Mod group. These results indicated that curcumin supplementation in a dose-dependent manner suppressed LCN2 expression in the progression of atherosclerosis.
Curcumin suppressed the formation of atherosclerotic lesions
We investigated whether curcumin attenuated western diet-induced atherosclerotic lesions in mouse aortas. Plaque area was measured using cross-sectional slices of the ascending aorta to estimate the progression of atherosclerosis. The analysis of representative images from the sections after staining with H&E (Fig. 2(A)) showed that atherosclerotic lesions in Mod group were markedly larger than those of Con group, plaque was predominantly observed in the medial and intimal areas of the arterial wall but less so in the adventitial layers, whereas small, sparse plaques were observed in the Lov group. Supplementing with curcumin in CurM group and CurH group reduced lesion development significantly in spite of high fat diet intake compared to Mod group. The percentages of aortic surface area occupied by the atherosclerotic lesions were significantly reduced in CurM group and CurH group compared with Mod group. Mice fed the low curcumin concentration in CurL group showed no suppressive effect.
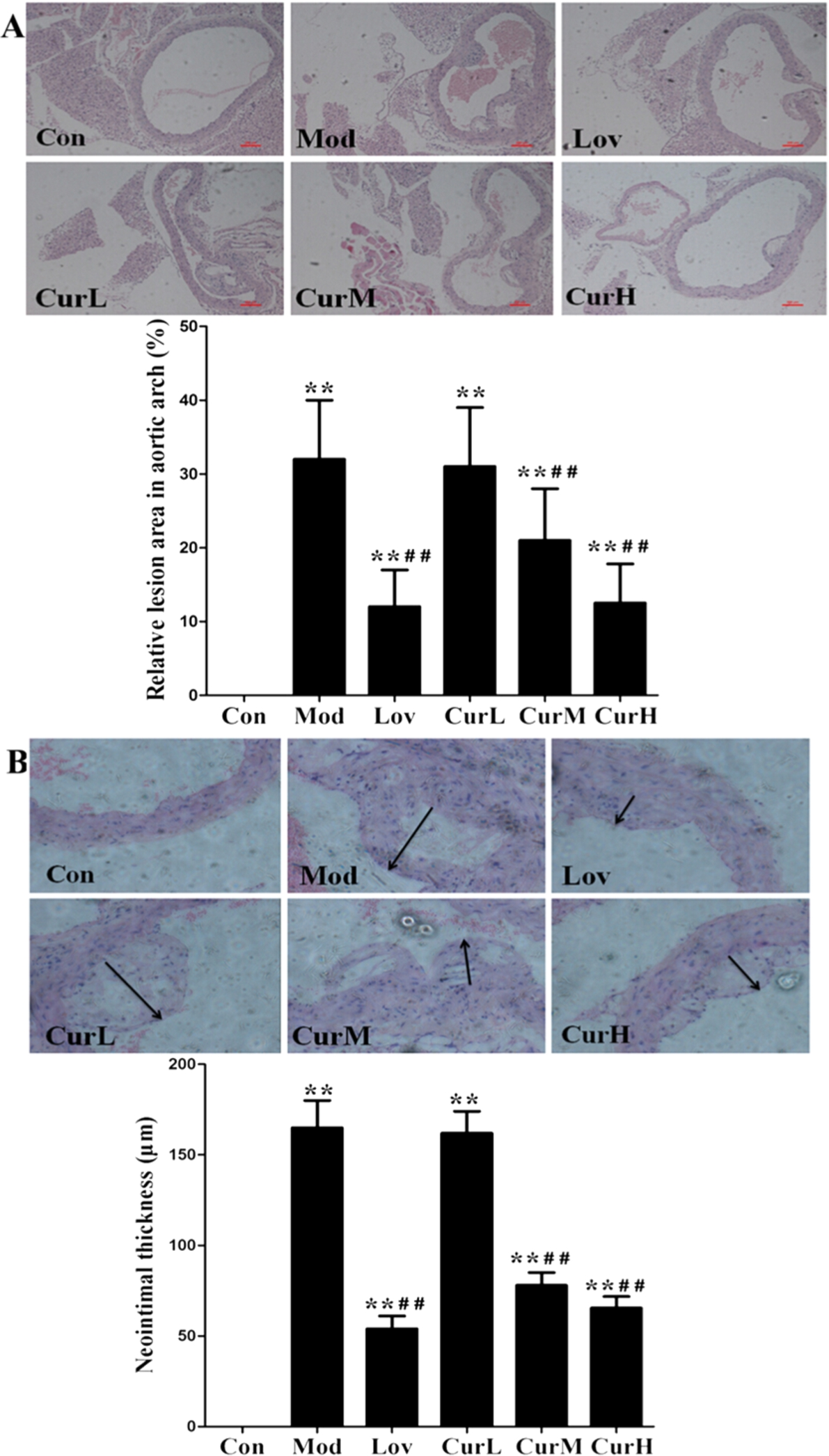
Effect of curcumin on atherosclerotic lesions formation in ApoE−/− mice fed a western diet for 12 weeks. (A) Representative photographs of aortic arch stained with H&E (40×). (B) Representative images of aorta neointimal thickness stained with H&E (200×). Arrows indicate neointimal thickness. Values are
Findings also revealed significantly greater neointimal thickening in Mod group compared with Con group. However, the curcumin supplement in CurM group and CurH group reduced the neointima formation (Fig. 2(B)). Taken together, these results indicated that curcumin suppressed the development of atherosclerotic lesions in aortas of western diet-fed mice.
ApoE−/− mice are unable to produce the key glycoprotein ApoE essential for transport and metabolism of lipids. Fed with normal chow, ApoE−/− mice start to develop atherosclerosis lesions at the age of 1 month to 2 months [12]. Therefore, we chose 6-week-old ApoE−/− mice on a high-fat diet for 12 weeks in this study to examine the development of atherosclerosis at initial stages. As our expectation, the sizes of atherosclerotic lesions within aorta in Mod group were remarkably increased as compared with Con group.
Adipose tissue is no longer considered as an inert tissue mainly devoted to energy storage but is emerging as a huge gland which participates actively in regulating physiologic and pathologic processes, including inflammation and immunity. Recently it has become clear that many adipokines which are secreted by adipose tissue, are mediators in the “adipo-cardiovascular axis”, the cross talk between adipose tissue and cardiovascular disease.
LCN2, mainly released from adipocytes, is an adipokine belonging to the lipocalin superfamily whose members generally bind a range of small hydrophobic molecules, specific cell-surface receptors and forms complexes with soluble macromolecules [13]. Recent studies seem to suggest a potential involvement of this adipokine in the genesis and progression of atherosclerosis. LCN2 level was positively correlated with lesion severity and complexity of coronary artery disease in patients with non-ST elevation acute coronary syndrome, and serum LCN2 levels on admission are associated with increased burden of atherosclerosis in these patients compared to control subjects. LCN2 also plays a critical role in the development and disruption of atherosclerotic plaques. Increased LCN2 is able to inhibit MMP-9 inactivation by forming a complex with MMP-9, thereby prolonging the enzymatic activity leading to degraded collagen content, weakening the atherosclerotic plaque structure, and making it more prone to rupture, which is the majority of acute clinical manifestations of atherosclerosis [14]. Thus, increased LCN2 results in an increase in plaque instability. From our results, serum LCN2 level, LCN2 protein in the aorta and the distribution of LCN2 in the atherosclerotic plaque in Mod group were much higher than in Con group [15]. These results indicated that LCN2 may play a significant role in the pathogenesis of atherosclerosis.
Hyperlipidemia, as a result of accumulation of lipids in blood has come to issue on the therapy for the atherosclerosis. Oxidized phospholipids contribute to inflammation within the artery wall, initiating atherogenic chemokine expression that leads to monocyte adhesion [16]. Therefore, lipids can be regarded as triggers of the inflammatory process in atherosclerosis. Clinical and epidemiologic observations have consistently documented that LDL-C concentration is positively correlated with atherosclerosis [17]. In this study, we found that lovastatin or curcumin treatment partly recovered high serum lipid profile induced by western diet and their anti-hyperlipidemia effects were comparable [18].
Activation of pro-inflammatory cytokines play a major role in the progression of atherosclerosis by increasing monocyte adhesion, smooth muscle cell proliferation, endothelial dysfunction, oxidative stress, and vascular calcification. In atherosclerosis patients marked elevated circulating levels of IL-6, TNF-α and hs-CRP were observed. IL-6 induces oxidative stress and endothelial dysfunction by over-expression of the angiotensin II type 1 (AT1) receptor in the atherosclerotic process [19]. TNF-α may play an atherogenic role by up-regulating the expressions of monocyte chemotactic protein-1 (MCP-1) in the vascular wall, and by inducing oxidized LDL (Ox-LDL) uptake and scavenger receptor class A expression in macrophages [20]. Hs-CRP is a classical acute-phase reactant, which binds to phosphocholine expressed on the surface of dead or dying cells and activates the complement system in atherosclerotic plaques via the C1q complex [21]. LCN2 was originally identified as an inflammatory cytokine released by activated neutrophils, and it could inhibit the secretion of anti-inflammatory factors such as peroxisome proliferator-activated receptor γ (PPARγ) and adiponectin in both adipocytes and macrophages [3,22]. In the present study, treatment with curcumin remarkably reduced the plaque area, LCN2 expression in atherosclerotic lesions and serum levels of LCN2, IL-6, TNF-α and hs-CRP in ApoE−/− mice compared with Mod group, and such interactions may help to explain the anti-atherosclerosis properties of curcumin.
Consumption of curcumin was shown to be beneficial to prevent atherosclerosis. A 6-month randomized controlled trial showed that curcumin intervention in type 2 diabetic population could lower the atherogenic risks by increasing serum level of adiponectin and decreasing serum level of leptin [23]. The key initiating step of the earliest stage of atherosclerosis is sub-endothelial accumulation of cholesterol and monocyte-derived macrophages, leading to chronic inflammation. Curcumin significantly suppressed MCP-1 production and up-regulated cholesterol efflux via suppressing the c-Jun N-terminal kinase (JNK) pathway, which indicated that curcumin could have the effect of anti-atherosclerosis via inflammation-related pathway [24]. In addition, curcumin increased the stabilization of atherosclerotic plaque by inhibiting MMP-9 expression through Protein Kinase C (PKC) signaling pathway in phorbol myristate acetate (PMA)-induced macrophages [25]. Our results indicated that the western diet significantly accelerated atherosclerosis and up-regulated adipokine LCN2 and several inflammatory cytokines such as IL-6, TNF-α and hs-CRP in ApoE−/− mice, dietary curcumin attenuated atherosclerosis by reducing LCN2 and inflammatory cytokines production.
Although the findings of our study could give new insights to the understanding of atherogenesis, in association with LCN2 expression, other adipokine should not be ignored in contributing to atherosclerosis, such as adipocyte fatty acid-binding protein (A-FABP). Curcumin was reported to alleviate atherosclerosis in LDL receptor deficient mice through suppression of A-FABP [26], the beneficial effects of curcumin in attenuating atherosclerosis may due to not only down-regulating LCN2 expression but also suppression of A-FABP.
Conclusion
In summary, our findings provide a potential biological basis for the association between an atherosclerosis-related biomarker and a Chinese herb extract, this presents the high beneficial effects of curcumin on lipid metabolism and cardiovascular health, particularly when these findings need to be translated for human use. Further studies are needed to add additional insights into the molecular mechanisms that drive down-regulating lipocalin-2 expression of curcumin in treating atherosclerotic lesions.
Footnotes
Acknowledgements
This work was supported by grants from the National Nature Science Foundation of China (No. 81660770), the National Nature Science Foundation of Jiangxi Province (No. 20161BAB215256), the Science and Technology Planning Project of Guangdong Province (No. 2016A020226023) and the China Postdoctoral Science Foundation (No. 2016M592476).
Conflict of interest
The authors have no conflict of interest to report.