
Abstract
Background:
KCNQ2 plays a key role in the regulation of neuronal excitability. The R214W and Y284C mutants of KCNQ2 channels, which are associated with BFNC, can decrease channel function to cause neuronal hyperexcitability and promote seizures. Previous studies revealed that elevated temperature caused up-regulation of KCNQ2 expression.
Objective:
The present study sought to investigate the impact of temperature elevation on neuronal KCNQ2 ion channel mutants.
Methods:
Protein expression of wt KCNQ2 and the R214W, Y284C and truncated selective filter mutants at different temperatures was detected by live-cell confocal fluorescence microscopy and by Western blotting. Whole-cell patch clamp was performed to record the effect of temperature on the electrophysiological activity of KCNQ2 channels.
Results:
Temperature elevation caused an unexpected increase in voltage-dependent KCNQ2 channel activation but also increased the endoplasmic reticulum (ER) retention of KCNQ2 protein, and the ER retention was greater for mutants associated with BFNC than for wt KCNQ2. Temperature elevation did not increase the fluorescence intensity of cells transfected with a truncated selective filter mutant.
Conclusions:
The direct effect of heat on KCNQ2 channels may be involved in excitability regulation of neurons, and the P-loop region is critical for temperature-dependent modulation of the expression and trafficking of KCNQ2 channels.
Introduction
The KCNQ2 subunit of voltage-gated channels has been proposed to mediate the neuronal M-current [1–4], which is a slowly activating, non-inactivating potassium current that partly determines neurons’ firing properties and responsiveness to synaptic input [5,6]. KCNQ2 channels are highly expressed in forebrain structures involved in several types of seizure, including the hippocampus, neocortex, and thalamus. KCNQ2 channels have also been reported to be densely localized at the axon initial segment (AIS) [7,8], where they are expected under physiological conditions to reduce neuronal excitability; augmentation of their function has been proposed as a potential anticonvulsant therapeutic approach [9]. For example, KCNQ channel activator flupirtine were found to be effective in repetitive febrile seizures prophylaxis and may be a candidate for the treatment of repetitive febrile seizures [10].
Studies have revealed that certain mutations in KCNQ2 channels can decrease channel function to cause neuronal hyperexcitability and promote seizures, as is believed to occur in benign familial neonatal convulsions (BFNC), a dominantly inherited idiopathic epilepsy [11], and myokymia [12]. R214W [13], a KCNQ2 mutation in the voltage-sensing S4 domain of KCNQ2 subunits has been shown to produce slower opening and faster closing kinetics. Y284C, a mutation in the pore region of the KCNQ2 subunit, exhibited decreased macroscopic currents without altering the membrane surface expression, further supporting a role for decreased KCNQ2 channel function in the promotion of seizures [14].
Temperature change is an important component of febrile seizures (FS), which are thought to be directly attributable to temperature-dependent changes in ion channel function. Studies have revealed that elevation of brain temperature alters many neuronal functions, including the properties of several temperature-sensitive ion channels [15] that contribute to shaping neuronal firing and synchronized neuronal activity. However, the mechanisms by which these translate to hyperthermia-induced seizures remain elusive [16].
As mentioned above, KCNQ2 was believed to be a promising target for FS treatment. However, few studies have focused on the effects of temperature elevation on voltage-gated channels. Recently, the effect of temperature changes on a voltage-gated channel was revealed. The sodium channel Nav1.2 was reported to be densely localized to AIS in hippocampal and neocortical neurons, and recombinant Nav1.2 channels exhibited a negative shift in the voltage-dependence of activation (in addition to faster kinetics) in response to elevated temperature [17]. Our previous studies showed that temperature can also increase the activation rates of KCNQ2 channels [18], but endoplasmic reticulum (ER) retention of KCNQ2 was revealed after temperature elevation. In this study, we investigated the impact on KCNQ2 channel trafficking and function of temperature elevation combined with mutations by comparing KCNQ2 channel properties at physiological (37°C) and febrile temperatures (40°C) and discuss the possible role of KCNQ2 mutants in FS.
Methods
DNA constructs and mutagenesis
Full-length cDNAs for the human KCNQ2 gene were polymerase chain reaction (PCR)-cloned from the plasmid pCDNA3.0-KCNQ2, gifts of Thomas Jentsch, Hamburg University, Hamburg, Germany. Then, KCNQ2 cDNAs were subcloned into the vector pEGFP-C1 (Clontech, Mountain View, CA, USA). Two nucleotide substitutions of KCNQ2 cDNA, R214W and Y284C, were introduced into the wild-type KCNQ2 construct in the pEGFP-C1 vector using the Quick-Change Site-Directed Mutagenesis Kit (Stratagene, La Jolla, CA). Truncated selective filter mutants were engineered in human KCNQ2 cDNA by sequence overlap extension PCR using KOD DNA polymerase. All of the inserted sequences, as well as the mutated sites, were confirmed by DNA sequencing.
Cell culture and cDNA transfection
HEK293T cells were cultured in Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 10% foetal bovine serum and incubated at 37°C in 5% CO2. Mouse neuroblastoma (N2a) cells were grown in 50% DMEM/50% OPTI-MEM with 5% foetal bovine serum. Cells were transfected with the recombinant plasmids using Lipofectamine 2000 (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s instructions. Cells were used for subsequent experiments 24 hours after transfections.
Live-cell confocal microscopy and fluorescence quantification
Temperature-controlled confocal microscopy was performed using the CTI Controller 3700 digital with the Tempcontrol 37-2 digital system (Leica Microsystems, Hong Kong, China). To identify the distribution of KCNQ2, it was co-expressed with a red fluorescent ER marker, pDSRed2-ER [19,20]. Over a given time series, the image acquisition settings used for the same cells were kept constant. The fluorescence intensity values of cells were determined using Leica Confocal Software. After incubation at 40°C, fluorescence intensities of channels were counted in the same regions using the same image acquisition settings. Co-localization images were analysed using ImageJ software (version WCIF, National Institutes of Health), using the plug-in “Manders’ Coefficient” to calculate the Manders’ coefficient [19,20].
Western blotting
Forty-eight hours after transfection, HEK293T cells were transferred from a 37°C incubator to a 40°C incubator for 2 hours. The cells were then washed with cold PBS buffer and lysed with RIPA lysis buffer (150 mM NaCl, 20 mM Tris, 1% sodium deoxycholate, 0.1% SDS, pH 8.0) containing protease inhibitor cocktail (Novagen, Merck KGaA, Darmstadt, Germany). The cell lysates were clarified by centrifugation, and the supernatants were collected to obtain total protein samples.
Cell membrane protein was collected using a plasma membrane protein extraction kit (TM-PEK; 71772-3; Novagen, Merck KGaA, Darmstadt, Germany) according to the manufacturer’s instructions. In brief, cells were washed with cold PBS and then permeabilized using Extraction Buffer 1. Then, the mixture was centrifuged at 1000 g for 5 min at 4°C, and the supernatant was collected to obtain cytoplasmic protein samples. Pellets were resuspended in transmembrane solubilization agent (Extraction Buffer 2B) and incubated for 45 min at room temperature with gentle agitation. Then, transmembrane proteins were extracted from the lipid bilayer with centrifugation at 16,000 g for 15 min at 4°C.
The protein concentration was determined using the bicinchoninic acid (BCA) protein assay (Pierce). Samples of 40
Electrophysiology
HEK293T cells were transfected with 1
Cells were bathed with mammalian Ringer’s solution: 2 mm KCl, 150 mm NaCl, 10 mm HEPES, 2 mm MgCl2, 10 mm D-glucose, pH adjusted to 7.2 using NaOH. The pipette solution contained 140 mm KCl, 5 mm EGTA, 5 mm NaCl, 1 mm Na2ATP, 10 mm HEPES (pH 7.2 with KOH). Electrodes had resistances of 1–3
Data analysis
Numerical data are expressed as the mean ± SEM. Statistical significance was evaluated using Student’s unpaired t test and was defined as
Results
Increased expression of mutant KCNQ2 after temperature elevation
To test the effect of temperature elevation on KCNQ2 mutations associated with BFNC, N2a cells were transfected with wt KCNQ2, the R214W mutant, or the Y284C mutant, and the fluorescence intensity of each group was compared between 37°C and 40°C. Total fluorescence intensities of both the R214W- and the Y284C-transfected cells were significantly increased after 30 minutes of incubation at 40°C (Fig. 1(A)), similarly to wt KCNQ2. Fluorescence intensities of the cells transfected with a blank pEGFP-C1 vector were unchanged or decreased with the temperature elevation (Fig. 1(A)), which proved that the increase of fluorescence intensity was not caused by the effect of elevated temperature on the GFP fusion protein (Fig. 1(A)). However, the difference in wt KCNQ2 fluorescence intensity between 37°C and 40°C was significantly smaller than that for R214W (
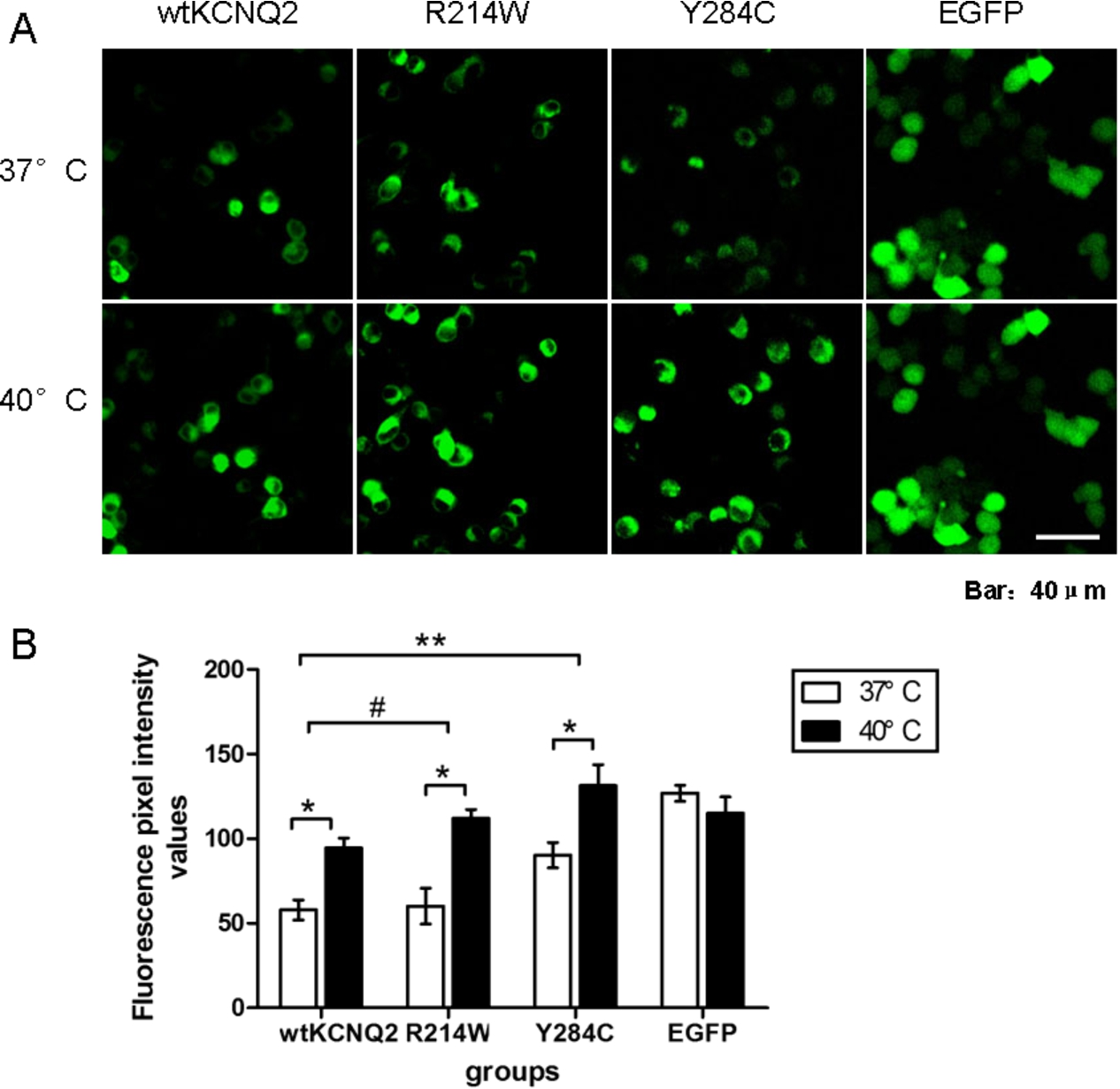
Elevated temperature increased the expression of wt and mutant KCNQ2 subunits. (A) Confocal images of N2a cells transfected with EGFP-wtKCNQ2, EGFP-R214W, EGFP-Y284C and EGFP at 37°C and 40°C. (B) Fluorescence intensity values of N2a cells transfected with EGFP-wtKCNQ2, EGFP-R214W, EGFP-Y284C and EGFP after 30 min of incubation at 37°C or 40°C (
To further determine the effect of temperature elevation on the expression of KCNQ2 channels, western blot analyses were performed on HEK293T cells transfected to express EGFP-tagged wt KCNQ2, R214W, or Y284C. Total cell lysates taken 24 hours after transfection showed an expected band for the EGFP-KCNQ2 fusion protein near 130 kDa MW (Fig. 2(A)). Quantification of western blots revealed that after one hour of temperature elevation, the expression levels of wt KCNQ2, R214W and Y284C subunits were significantly increased (
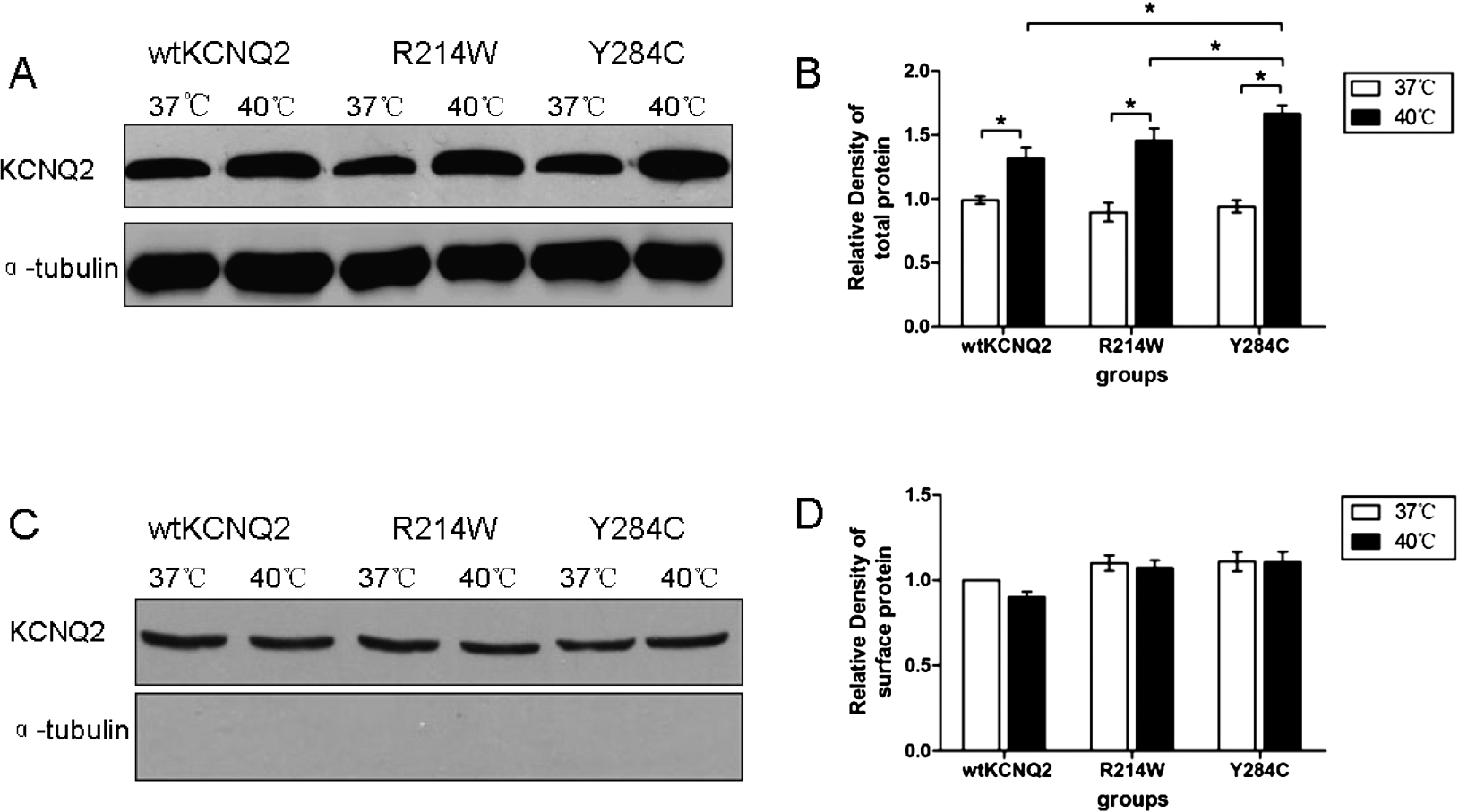
Western blot analysis. (A) Total cell lysates from cells transfected with the plasmids encoding wt KCNQ2, R214W and Y284C subunits. The lower image in this panel shows the same blot stripped and reprobed with anti-α-tubulin antibody as a loading control. (B) Quantification of western blots of the total cell lysates from HEK293T cells expressing wtKCNQ2, R214W or Y284C subunits incubated at different temperatures. (C) Cell surface protein expression of wtKCNQ2 and mutants incubated at 37 or 40°C for 1 h. (D) Quantification of western blots of cell membrane protein. In all groups, data are presented as the mean ± SEM (
The relative contribution of ion channels to neuronal excitability is determined by their functional characteristics, abundance, and distribution at the plasma membrane [21]. To identify whether KCNQ2 channels were expressed intracellularly or at the cell surface, KCNQ2 subunits were coexpressed with an endoplasmic reticulum (ER) marker, pDsRed2-ER [19,22,23]. (Fig. 3(A) to (C)). At 37°C, the fraction of the Y284C mutant in the ER (0.70) was clearly greater than those of wt KCNQ2 (0.53) and the R214W mutant (0.64) (Fig. 3(D)), which could explain the functional decrease in current caused by this mutation. After a 30 min incubation at 40°C, the majority of transiently expressed wt or mutant KCNQ2 was distributed intracellularly and retained in the ER (Fig. 3(A) to (D)). Indeed, co-localization analysis revealed that the fraction of Y284C retained in the ER was clearly greater at 40°C (0.77) than at 37°C (0.70), as well as the fractions of wt KCNQ2 (0.61) and R214W (0.71) retained at 40°C. These results demonstrated that temperature plays a critical role in the trafficking of KCNQ2 to and from the surface membrane, that an epilepsy-associated mutation can alter this process, and that the effect of Y284C was greater than that of R214W.
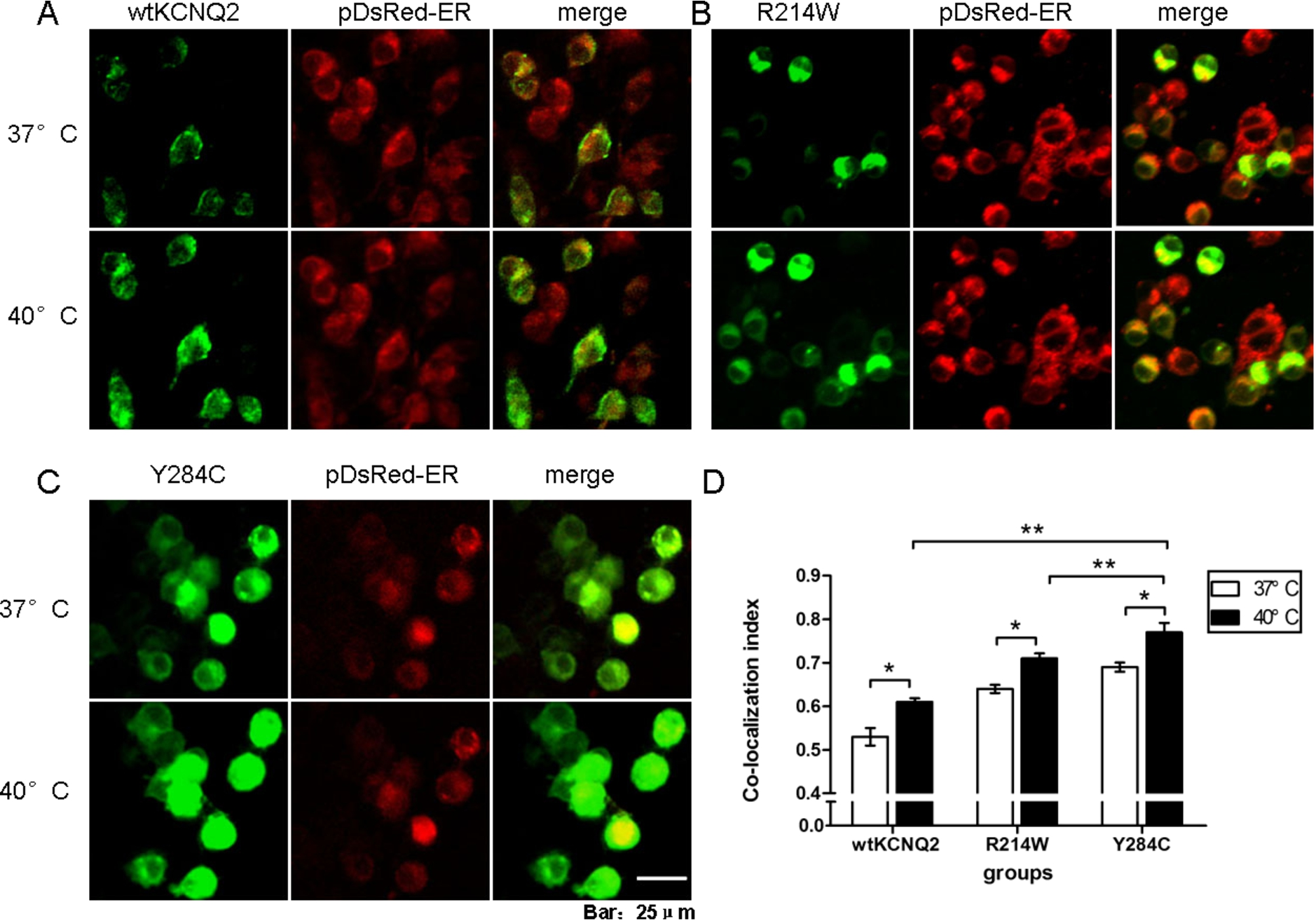
Elevated temperature increased intracellular retention of wtKCNQ2 and mutants subunits. (A)-(C) Confocal microscopy images of N2a cells cotransfected with EGFP-tagged wt or mutant KCNQ2 subunits and with a pDsRed-tagged ER marker after a 30 min incubation at 37°C or 40°C. The cells were processed as described in the Methods. The KCNQ2 subunits and the ER marker were pseudocoloured in green and red, respectively, resulting in a yellow colour when colocalized. (D) Averaged Manders’ coefficient of colocalization in cells expressing the different constructs at different temperatures (
Elevated temperature significantly affected the voltage-dependence of KCNQ2 channel activation (Table 1). At 37°C and 40°C, both the fast and slow time constants of activation (
The pore region is essential for the effects of temperature on KCNQ2 channels
The above results suggested that the effects of temperature elevation were more profound on the Y284C mutant compared to the R214W mutant or the wt channel. At the higher temperature, the expression of KCNQ2 Y284C homomeric channels was more clearly increased, and a higher fraction of these were retained in the ER. We thus speculated that the P-loop region may be essential for the modulation of KCNQ2 channels by temperature elevation. Confocal microscopy indicated that temperature elevation did not increase the fluorescence intensity of cells transfected with a truncated selective filter mutant (Fig. 4(A) to (B)). The ER fraction of selective filter truncated mutant channels was also unchanged after temperature elevation (Fig. 4(C) to (D)). Thus, the P-loop region is critical for the temperature-dependent modulation of expression and trafficking of KCNQ2 channels.
Discussion
Temperature changes are an important component of many disease processes (e.g., fever, inflammation), and some manifestations such as febrile seizures are thought to be directly attributable to temperature-dependent changes in ion channel function. Our previous studies [18] indicated that KCNQ2 subunits showed increased intracellular localization with temperature elevation, suggesting that KCNQ2 channels could be involved in temperature-responses and indicating that elevated temperature alters the expression, gating, and trafficking of KCNQ2 channels [18]. In this study, our data indicated that KCNQ2 mutant channels that are associated with a form of childhood epilepsy (BFNC) were regulated by temperature similarly to wt channels but that one mutation (Y284C) exhibited more labile temperature sensitivity. Similar to wt, the two mutant subunits showed increased expression and intracellular localization at the elevated temperature. However, the magnitude of increased expression was significantly greater than for the wt, and the channel fraction localized intracellularly was significantly greater for the Y284C mutant compared to both wt and R214W. The Y284C mutation also displayed a greater negative shift (∼20 mV) in steady-state voltage dependence of activation. Taking these data together, the Y284C mutant appeared to exhibit generally more sensitivity to temperature, and our data further suggest that mutations in the pore region alter temperature sensitivity more profoundly than mutations in the S4 domain. Our results support the idea that the pore region plays a very prominent role in the trafficking of KCNQ channels.
Gating properties of channels carrying KCNQ2 mutation

Expression of the truncated selective filter mutant did not change at high temperature. (A) Confocal images of N2a cells transfected with truncated mutant at 37°C and 40°C. (B) Fluorescence intensity values of the truncated mutant did not change after 30 min of incubation at 37°C and 40°C. (C) Confocal images of N2a cells cotransfected with the truncated subunits and a pDsRed-tagged ER marker. (D) Averaged Manders’ coefficient of colocalization showed no significant differences between 37°C and 40°C. (
It has generally been thought that the pore region does not play a significant role in trafficking of KCNQ2/3 channels and that surface expression is controlled by the intracellular C-terminal region [19,24–26]. However, a KCNQ3 mutation in the pore region (A315T) was demonstrated to increase the stability and cell-surface trafficking of homotetramers [20]. Consistent with our results, a mutation in the inhibitory GABAA receptor
The specific mechanisms by which temperature regulates ion channel function are varied, and evidence indicates that heat can directly affect channel gating [17] and trafficking [27]. However, despite increased total KCNQ2 expression, elevated temperature also increased the intracellular fraction of channels, suggesting less availability of membrane surface channels. It is remarkable that although KCNQ2 is only functional in the plasma membrane, the majority is located in the ER and other intracellular compartments [30]. Temperature elevation increased the expression of KCNQ2, which would suggest more channels available for activation. However, greater ER retention of KCNQ2 was revealed after temperature elevation [18]. In this study, ER retention was also revealed for KCNQ2 mutants. Interestingly, the KCNQ2 macroscopic current density did not change upon rapid temperature elevation, indicating that the functional expression of the protein in the cell membrane was likely unaffected under the recording conditions. This may have been partly due to the ER retention of KCNQ2.
KCNQ trafficking is a complicated process that involves protein assembly, ER exit, endocytosis, and degradation [31]. ER retention will affect KCNQ trafficking but the mechanism underline ER retention is unknown. The cytoskeletal scaffolding protein ankyrin G (ankG) plays an important role in KCNQ2 trafficking. In axons, ankG is the only known binding partner of KCNQ2 and KCNQ3, and KCNQ2/3 colocalizes with ankG at the AIS [8]. The ability of KCNQ2/3 to bind AnkG is markedly enhanced by protein kinase CK2 (CK2) phosphorylation [32]. CK2 has been found to regulate KCNQ2 activity through phosphorylation and thereby enhance KCNQ2 binding to AnkG [33–35]. Then, KCNQ2 channels are transported to the axon hillock, admitted to the AIS, and phosphorylated at their anchor domains by local CK2 [32]. Hence the regulation of KCNQ ER retention by temperature elevation should be further investigated.
In this study, KCNQ2 mutants were up-regulated after temperature elevation, but no obvious changes in KCNQ2 macroscopic current density were observed, which is partly due to KCNQ2 ER retention. KCNQ2 channels might contribute to the pathogenesis of febrile seizures, but KCNQ2 channel activation may also be a viable strategy for treating febrile seizures. The direct effect of heat on KCNQ2 channels may be involved in the regulation of neuronal excitability, and the P-loop region is critical for temperature-dependent modulation of expression and trafficking of KCNQ2 channels. The precise mechanism of ER retention after temperature elevation should be further investigated.
Footnotes
Acknowledgements
This work was supported by the National Natural Sciences Foundation of China (No. 81401241, No. 81371422, No. 81401230), the Foundation of Health and Family Planning Commission of Hubei Province (WJ2015MB112) and the Translational Medical Research Fund of Wuhan University school of medicine.
Conflict of interest
The authors have no conflict of interest to report.