
Abstract
BACKGROUND:
The effectiveness of bone repair is determined by the balance of proliferative and destructive factors in the fracture union site. It can be enhanced by using various nanostructured materials possessing osteoinductive properties, in particular titanium implants with biodegradable calcium phosphate coatings. The effects of these coatings on the state of stem cells, their differentiation and distribution in the repair zone is unknown.
OBJECTIVE:
To study the dynamics of proliferation, differentiation, and apoptosis of stem cells after experimental fracture followed by implantation of titanium implants with calcium phosphate coatings.
METHODS:
The localization of proliferation (PCNA) and differentiation (CD44 and osteocalcin) factors and apoptotic molecules (MDM2, p53, caspase-3) was studied in a rat femoral fracture model with implant placement. Titanium implant screws with bioactive calcium phosphate and hydroxyapatite coatings formed by plasma electrolytic oxidation were used in the study. Experimental rats were arranged into three groups (15 animals per group): control group; rats implanted with uncoated implants; and rats implanted with coated implants. Control rats were subject to a similar fracture as experimental ones and were allowed to heal conservatively. Rats from all groups were sampled on days 7, 14, and 30 after injury.
RESULTS:
Low-differentiated PCNA-, osteocalcin-, and CD44-immunopositive cells were localized around the implant in the inner layer of the periosteum, layer of outer circumferential lamellae, and connective tissue lining of haversian canals. The spatial density of cells expressing the above proliferation and differentiation factors, as well as that of MDM2-immunoreactive cells, increased on day 7 and decreased by day 30 after injury. The spatial density of apoptotic cells reached the maximum on day 14 after injury. They were mainly found in the inner layer of the periosteum and outer circumferential lamellae. p53- and caspase-3-positive cells occurred on the surface of the concentric lamellae surrounding haversian canals and under the periosteum. Their spatial density decreased by day 30 after injury.
CONCLUSIONS:
Calcium phosphate coatings stimulate cell proliferation at early stages of fracture restoration and apoptotic cell death at later stages. Coating components may provide positional information guiding the differentiation of mesenchymal stromal cells. A change in the activity of apoptotic factors, osteocalcin, and CD44 is caused by gene induction in response to the diffusion of calcium phosphate compounds from coating to surrounding tissue.
Introduction
The effectiveness of bone repair is determined by a variety of growth maintaining the balance between cell death, proliferation, and differentiation. Bone tissue is characterized by an exceptionally high concentration of stem cells involved in impaired bone repair after injury [1]. Both in normal bone renewal and injured bone repair, new cells are derived from mesenchymal stem cells (MSCs) of the periosteum and endosteum [2]. MSCs also participate in vasculogenesis [2].
In earlier studies we showed that bioactive calcium phosphate and hydroxyapatite coatings of titanium implants stimulate the expression of vascular endothelial growth factor (VEGF) and transforming growth factor-beta 2 (TGF-𝛽2) in osteoblasts [3,4]. These factors enhance osteogenic differentiation. They are also involved in callus remodeling at late stages of osteogenesis. The TGF-𝛽 family includes bone morphogenetic proteins (BMPs) that participate in osteogenesis induction [4]. BMPs are produced by MSCs, osteoblasts, and chondrocytes. They promote osteogenic proliferation and differentiation, stimulate the synthesis of collagen and osteocalcin, enhance extracellular matrix mineralization, and modulate osteoclasts activation [2].
MSCs are selectively marked by CD44, and MSC-derived osteoblasts selectively express osteocalcin [2]. Bone tissue regeneration is associated with excessive proliferation of MSCs and is regulated by apoptotic signals [5]. The apoptosis is initiated by sequential expression of signal molecules including p53 and caspase-3 [6]. Unlike them, MDM2 is shown to inhibit apoptosis [6]. Fracture repair requires close interaction of gene induction, apoptotic molecules, and differentiation factors. The mechanisms coordinating these factors and processes remain largely obscure.
The present study deals with the dynamics of cell proliferation, differentiation, and apoptotic death after experimental fracture followed by implantation of titanium implants with calcium phosphate coatings.
Materials and methods
Implant samples
Standard titanium implant screws (titanium grade BT-6) and similar implants with bioactive calcium phosphate and hydroxyapatite coatings were used in the study. The implants were manufactured by scientific production associations Deost and Osteomed, Russia. Titanium grade BT-6 is identical to medical ASTM F136 titanium Ti-6Al-4V alloy. Before coating, the implants were machined to 0.12 μm surface roughness. They were then washed in distilled water and degreased with alcohol. Plasma electrolytic oxidation (PEO) coatings were formed in electrolyte containing calcium citrate (Ca3(C6H5O7)2
Some parameters of the superficial layer of PEO coatings examined in vivo
Some parameters of the superficial layer of PEO coatings examined in vivo
200–250 g male rats were used in the study. All experiments were performed in strict accordance with the local experimental animal treatment regulations and approved by a local ethical committee. They were also compliant with the EU Directive 2010/63/EU for animal experiments. The animals were kept in a vivarium meeting standard requirements to animal keeping and treatment under experimental clinical conditions.
All surgeries were performed under aseptic conditions and sevoflurane anaesthesia. Low-invasive closed osteosynthesis resulting in indirect bone healing was modeled. The surgical site was disinfected. To access the intercondylar fossa of the femur bone, a 0.5 cm skin incision was made over the left knee flexed to over 90°. In this position, the patella is above the intercondylar fossa. The patella ligament was split longitudinally to keep the extensor muscles of the crus intact. To reach the femur cavity, an inlet hole was formed in the intercondylar notch using a thin trocar. The intramedullary cavity was widened with sequential use of 1.5 and 1.8 mm Kirschner’s wires. In this case, a 2 mm diameter screw was implanted. If cavity widening using a 1.5 mm Kirschner’s wire was achieved without noticeable effort, the cavity was further widened using a 2 mm Kirschner’s wire. In this case, a 2.3 mm diameter screw was implanted. The inlet hole was widened using an end reamer so that the screw head could be buried in the subchondral layer. A screw was placed into the hole so that its tip protruded from the middle third of the femur shaft. To model a closed fracture, the femur shaft was broken manually at its middle (near the screw tip). Bone fragments were manually set and fixed. To restore knee functionality, the screw was tightened into the central bone fragment until the screw head was completely buried in the subchondral layer. The wound was closed and antiseptically treated. This technique of osteosynthesis and bone repair is preferred in the clinical practice for femoral shaft fractures. The fracture and correct implant position were confirmed by radiovisiography using an X-ray apparatus Evolution X3000 2C Asepti. X-ray graphs were taken in the side projection at 0.08 s exposure, 40 kV tube voltage, and 25 mAs tube current-time product.
Experimental rats were arranged into three groups (15 animals per group): control group; rats implanted with uncoated implants; and rats implanted with coated implants. Control rats were subject to a similar fracture as experimental ones and were allowed to heal conservatively. Rats from all groups were sampled on days 7, 14, and 30 after injury. The animals were euthanatized under general anaesthesia by intraperitoneal injection of 3% sodium thiopental followed by decapitation.
Immunochemical examination
Femoral bone samples were fixed in 4% neutral formaldehyde on 0.1 M phosphate buffer for 24 h, washed in six-seven changes of 0.1 M sodium phosphate buffer (pH 7.2), decalcified in 25% Trilon B solution (pH 7.0) using Orbital Shaker OS-20 Biosan at room temperature. An implant was extracted through the longitudinal slit in the femur without injuring surrounding tissue. The samples were then washed and paraffin embedded following standard protocol. Fifteen μm thick sections were mounted onto slides, deparaffinized, and incubated in 3% hydrogen peroxide for 15 min to block endogenous peroxidases. They were then washed in three changes of 0.1 M sodium phosphate buffer (pH 7.2), incubated in 2% BSA and 0.25% Triton X-100 for 1 h, and incubated with primary antibodies overnight at 4 °C. The following primary antibodies were used: rabbit polyclonal antibodies against proliferating cell nuclear antigen (PCNA) and CD44 (Abcam, UK) diluted to 2 μl/ml and 1:500, respectively; mouse monoclonal antibodies against osteocalcin [OCG3] (Abcam, UK) diluted to 10 μl/ml; rabbit polyclonal antibodies against MDM2 and p53 (Invitrogen, USA) diluted 1:100; and rabbit monoclonal antibodies against caspase-3 (Thermo Fisher Scientific, USA) diluted 1:50. The primary antibodies were diluted in phosphate buffer containing Triton X-100 and BSA. The sections were then washed and incubated with biotinylated secondary antibodies against rabbit or mouse immunoglobulin (depending on primary antibodies) diluted as recommended by the manufacturer (Abcam, UK). The sections were then washed and incubated in avidin-peroxidase complex solution (Abcam, UK), developed in 3,3 ′ -diaminobenzidine solution (DAB Substrate Kit, Abcam, UK) for 4 min, washed, dehydrated, and mounted in Canada balsam following standard protocols. Control sections were treated as described above, but were not incubated with primary antibodies. No specific labeling was found in these sections. Some sections were stained with hematoxylin-eosin.
The sections were examined using an AxioScope A1 microscope (Carl Zeiss, Germany). Photomicrographs were taken by means of an AxioCam ICc3 digital camera. Numerical parameters were estimated using the AxioVision 4.8.1 package. The spatial density of cells was measured as cells per 0.01 mm2 of bone section, corrected for section thickness and nucleus diameter. The significance of differences was assessed using the Student t-test at P < 0.05.
Results
Using a number of calcium-containing electrolytic systems and oxygenation regimes, we demonstrated that by means of PEO, various calcium-phosphate coatings including hydroxyapatite ones may be formed on the surface of implants made of Ti-6Al-4V alloy. Table 1 presents data on the electrolyte composition, PEO parameters, phase and element composition, and Ca/P ratio of titanium implant coatings obtained in the electrolytes containing calcium citrate, calcium acetate, and twice-substituted sodium phosphate. Their examination showed that the coatings obtained in acetate-containing electrolytes contained hydroxyapatite (Table 1). The crystalline phase of the coatings formed in citrate-containing electrolyte consisted of titanium dioxide. However, proceeding from their Ca/P ratios, their amorphous phase contained calcium phosphate compounds.
On day 7 after injury, visible signs of bone regeneration in the fracture site could be seen. In the control animals, in the immediate vicinity of the fracture, the thickness of the periosteum increased to 189.4 ± 11.3 μm. Under the periosteum, woven bone tissue was observed to form parallel layers. It contained separate randomly interlaced collagen fibers resembling amorphous connective tissue. Large basophilic cells up to 20 microns in diameter with morphological signs of osteoblasts were clearly visible between the fibers. On day 14 after injury, distinct bone lamellae were present. By day 30 after injury, lamellar bone tissue predominated. The system of newly formed osteons appeared on day 14 after injury. In the immediate vicinity of the fracture site, young osteons 87.6 ± 5.1 μm in diameter were observed. They were formed by 2–3 lamellar layers surrounding a wide haversian canal.
The distribution of PCNA-expressing cells in the repair site was inhomogeneous (Fig. 1). On day 7 after injury, their density reached maximum (inner layer of the periosteum: 19 ± 1.4/0.01 mm2; outer and inner circumferential lamellae: 7 ± 0.4 and 9.7 ± 0.5/0.01 mm2, respectively; connective tissue lining of haversian canals: 17.6 ± 2.8/0.01 mm2. By day 30 after injury, the density of PCNA-positive cells decreased (periosteum: 14 ± 0.9/0.01 mm2; outer circumferential lamellae: 3.2 ± 0.03/0.01 mm2; lining of haversian canals: 6.2 ± 0.3/0.01 mm2) indicating a decrease in proliferative activity. Osteocalcin- and CD44-immunoreactive cells showed the same trend (Figs 2, 3A).
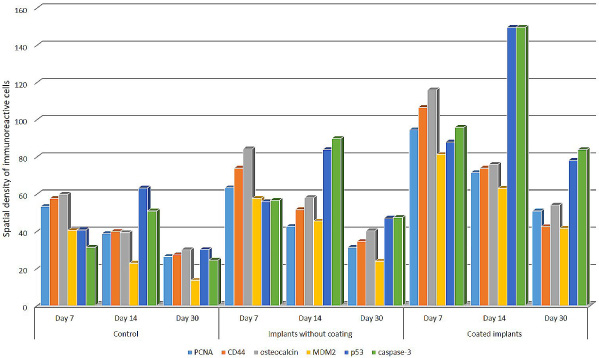
The changes in the spatial density of PCNA-, CD44-, osteocalcin-, MDM2, p53-, and caspase-3-immunopositive cells (cells per 0.01 mm2) in rat bone tissue after experimental fracture.
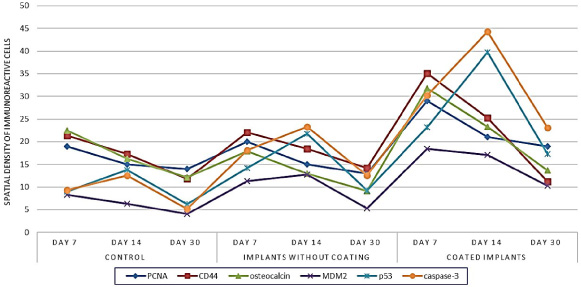
The spatial density of PCNA-, CD44-, osteocalcin-, MDM2-, p53-, and caspase-3-positive cells in the periosteum at different stages of bone regeneration after experimental fracture.
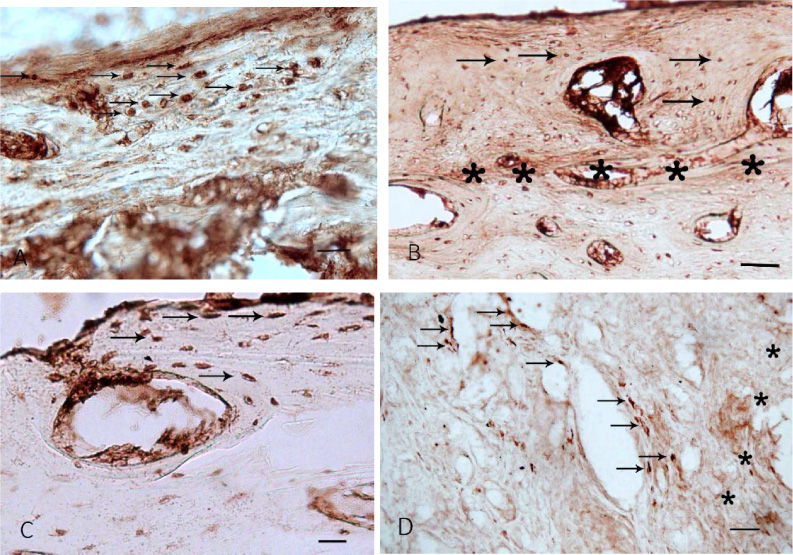
Osteocalcin, CD44, caspase-3, and p53-immunopositive cells in rat bone tissue after experimental fracture. (a) CD44-positive cells (putative osteoblasts) in the inner layer of the periosteum and outer circumferential lamellae (arrows). Control rats, day 7 day after fracture. (b) p53-positive cells on the surface of outer circumferential lamellae under the periosteum and around haversian canals (arrows). The tissue layer adjacent to the implant surface is marked with asterisks. Control rats, day 14 day after fracture. (c) Osteocalcin-positive cells at the outer circumferential lamellae and young osteons (arrows). Rats with uncoated implants, day 7 after fracture. (d) Caspase-3-positive cells in concentric lamellae of young osteons and haversian canals (arrows). The tissue layer adjacent to the implant surface is marked with asterisks. Rats with uncoated implants, day 14 after fracture. Scalebar: (a) 50 μm, (b, c, d) 100 μm.
Throughout the experimental period, the expression rate of all apoptotic markers changed in a similar way. p53-, caspase-3-, and MDM2-positive cells were localized in the periosteum and at the surface of the lamellae (Fig. 3B). The maximum expression rate of proapoptotic factors was registered in the fracture site on day 14 after injury. The density of p53- and caspase-3-positive cells in the periosteum was 13.8 ± 0.9 and 12.5 ± 1.5/0.01 mm2, and in the layer of outer circumferential lamellae 18.7 ± 0.7 and 11.3 ± 0.7/0.01 mm2, respectively. A similar expression rate of proapoptotic factors was observed in low-differentiated cells in connective tissue lining of haversian canals and in osteocytes situated between concentric lamellae of newly formed osteons. Similarly to the expression dynamics of the proliferation and differentiation markers, the expression rate of MDM2 steadily decreased from the day 7 to day 30 after injury (Fig. 1).
In the rats implanted with uncoated implants, the distribution of PCNA-, osteocalcin-, and CD44-immunoreactive cells did not differ much from that in the control group (Fig. 3C). The maximum MSC proliferation rate was registered on day 7 after injury in the vicinity of implants, mainly in the periosteum and osteons, where the density of PCNA-positive cells was 20 ± 1.3 and 21.4 ± 1.7/0.01 mm2, respectively. The density of differentiating osteocalcin- and CD44-immunoreactive cells changed in a similar way dropping down substantially by day 30 after injury. The density of MDM2-immunoreactive cells followed the same dynamics. The maximum density of caspase-3- and p53-positive cells was registered in the periosteum, newly formed lamellae, and haversian canals on day 14 after injury (Figs 1, 3D).
In the rats implanted with coated implants, at all times after injury, the density of cells immunoreactive to the markers used in this study differed substantially from that in the rats implanted with uncoated implants. During fracture repair, PCNA-, CD44-, and osteocalcin-positive cells were observed in the osteogenic layer of the periosteum, at the surface of outer circumferential lamellae, and in connective tissue lining of haversian canals (Fig. 4A, B). On day 7 after injury, an increase in proliferative activity was detected in the vicinity of the implant surface. At that moment, in connective tissue of osteons, the density of PCNA-positive cells reached maximum (35.1 ± 1.8/0.01 mm2). It decreased considerably by day 30 after injury (16.3 ± 1.1/0.01 mm2). The density of CD44- and osteocalcin-positive cells changed in a similar way (Fig. 1, 2). The peak density of apoptotic cells was registered in the inner layer of the periosteum and in outer circumferential lamellae on day 14 after injury. At that moment, the density of p53-positive cells in the periosteum, outer circumferential lamellae, and connective tissue lining of haversian canals was 39.7 ± 1.2, 45.2 ± 2.9, and 30.6 ± 1.1/0.01 mm2, respectively. On day 14 after injury, caspase-3-positive cells were mainly localized in the periosteum (44.3 ± 3.5/0.01 mm2). In osteons and outer circumferential lamellae, their density was lower (36.3 ± 2.1 and 33.2 ± 4.7/0.01 mm2, respectively). On day 30 after injury, the density of p53- and caspase-3-immunoreactive cells decreased, they were located mainly on the surface of newly formed concentric lamellae and under the periosteum (Figs 1, 4C, D).
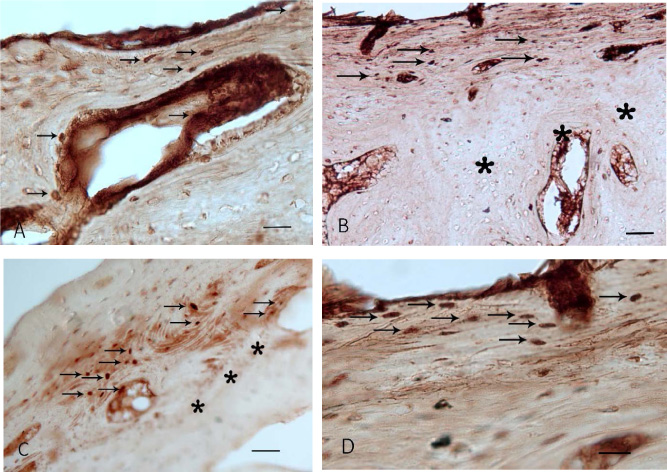
Proliferative and proapoptotic factors in rat bone tissue after experimental fracture followed by implantation of coated implants. (a) Overexpression of CD44 in cells of haversian canals and between outer circumferential lamellae under the periosteum (arrows). Day 7 after injury. (b) Increased levels of osteocalcin in osteoblasts in outer circumferential lamellae and osteons (arrows). The tissue layer adjacent to the implant surface is marked with asterisks. Day 7 after injury. (c) Overexpression of caspase-3 in concentric lamellae of osteons and haversian canals (arrows). The tissue layer adjacent to the implant surface is marked with asterisks. Day 14 after injury. (d) p53-immunoreactive cells (arrows) under the periosteum. Day 14 after injury. Scalebar: (a, d) 50 μm, (b, c) 100 μm.
By means of PEO, a variety of calcium phosphate coatings including hydroxyapatite ones may be formed on titanium surfaces. In the present study, calcium-containing salts with the Ca/P ratio close to that in human bone tissue (1.67) were added to electrolytes to form PEO coatings on titanium implants [7]. The formation of hydroxyapatite in the surface layer of coating is determined by PEO polarization regime. Hydroxyapatite (Ca10(PO4)6(OH)2) coatings were only formed at the bipolar regime. Probably, in this case, the inversion of electrode polarity results in alternate saturation of the peri-electrode space with calcium and phosphate ions. At cathode polarization, the superficial zone of electrolyte is saturated with Ca2+ ions, and at anode polarization, with (HPO4)2− ions. They interact to form various calcium phosphate compounds: Ca10(PO4)6(OH)2, Ca3(PO4)2, CaHPO4⋅2H2O. In addition, in the bipolar PEO regime, the plasma contained a substantial proportion of electrolyte components. This results in more intense electrochemical synthesis of calcium phosphate compounds compared to the unipolar regime [7].
Here, we examined osteoinductive properties of calcium-phosphate implant coatings affecting cell proliferation and apoptosis after experimental fracture. In early fracture repair, the thickness of the periosteum and connective tissue lining of haversian canals increased. An increase in the number of PCNA-positive cells in these structures marks the beginning of bone repair. The inner layer of the periosteum comprises an extensive system of microvessels, as well as fibroblasts and adventitious cells [8]. In the rats implanted with coated implants, the number of CD44-positive stem cells in this layer was greater than in the rats implanted with uncoated implants. The differentiation of MSCs into osteogenic lineage was confirmed by a high expression rate of osteocalcin in late osteoblasts.
MSCs and osteoprogenitor cells can migrate towards titanium implant surface (possessing osteoinductive and osteoconductive properties), differentiate into osteoblasts, form tight attachments to implant surface, and develop new bone tissue [9,10]. These processes are initiated and controlled by growth factors produced by MSCs that maintain a complex balance of paracrine and autocrine signaling in osteogenic differentiation [11]. For example, BMP-2 is expressed during the first days of fracture restoration. It stimulates the differentiation of MSCs and osteoprogenitor cells into osteoblasts [2,12]. Later on, BMP-6 and BMP-9 are expressed. These factors together with TGF-𝛽 inhibit the activation of osteoclasts and initiate their apoptotic death [13,14]. At the same time, BMP affects the secretion of vascular endothelial growth factor (VEGF), which in turn promotes the migration of endothelial cells and vasculogenesis [15,16].
Calcium-phosphate coatings can modify the osteoinductive properties of morphogenetic molecules thereby changing the balance of pro- and anti-apoptotic factors regulating the delicate mechanics of bone restoration [4]. After the period of active osteogenesis, some osteoblasts die by apoptosis [17,18]. It is induced by BMP-2, which upregulates the expression of Bax protein and downregulates that of Bcl-2, enhances the release of mitochondrial cytochrome c into the cytosol, and activates caspases 9, 3, 6 and 7 [19].
The apoptotic markers used in this study label cells at different stages of apoptosis. p53 initiates mitochondrial apoptotic pathway. It therefore labels early apoptotic cells. The expression of effector caspase-3 promotes activation of Ca2+/Mg2+-dependent endonuclease and DNA and nuclear fragmentation. MDM2 protein binds p53 and blocks its function thereby interrupting the apoptotic cascade [18,20]. In accordance with our data, the expression of MDM2 correlates with the density of PCNA-immunopositive cells. The peak of proliferative activity occurred on day 7 after injury and then gradually decreased by day 24. The proliferative and apoptotic activities showed the opposite dynamics. This suggests that apoptotic cell death is involved in ultimate osteoblast positioning and remodeling of newly formed bone tissue.
To summarize the above, hydroxyapatite coatings have a strong effect on apoptotic cell death enhancing it in late stages of repair. Implant coatings can regulate ion homeostasis in the fracture site at the stage of soft callus by imitating the mineral composition of mature bone. Calcium phosphates may provide positional information and direct the differentiation of MSCs. BMP may have a similar osteoinductive effect on MSCs by activating MAP-kinase and canonical WNT-dependent signaling [4,21]. Together, these molecules contribute much to the regulation of bone repair.
Footnotes
Acknowledgements
The authors are grateful to the two anonymous reviewers for their valuable suggestions and comments on the manuscript.
Conflict of interest
The authors report no conflict of interest. The authors alone are responsible for the content and writing of the article.