
Abstract
Cell and tissue engineering therapies provide promise for regenerating damaged intervertebral disc (IVD) tissue and resolving the low back pain that often accompanies it. However, these treatments remain experimental and unavailable for patients. Furthermore, the large body of work characterizing and utilizing mesenchymal stromal cells (MSCs) for these applications has, unfortunately, not resulted in any FDA-approved spinal therapies. Herein, we characterized DiscGenics’s human cadaver-derived discogenic nucleus pulposus (NP) progenitor cells and, for the first time, their discogenic annulus fibrosus (AF) progenitor cells. We then used these discogenic NP and AF cells to create biomimetic human-sized total tissue-engineered IVD replacements, also known as endplate-modified angle ply structures (eDAPS), and compared these with eDAPS formulated with goat or human MSCs. Prior to eDAPS fabrication, discogenic cells were expanded using either two-dimensional attachment culture or three-dimensional suspension culture. Currently, no data exist as to how these discogenic progenitor cells deposit extracellular matrix in a 3D culture environment, nor do data exist characterizing whether the unique expansion environment influences subsequent discogenic cell behavior. Our data support that NP and AF discogenic cells occupy unique niches and serve distinct functions, both in the IVD and in an in vitro 3D culture environment. As a result, discogenic cells deposited more matrix overall than did MSCs. That matrix was distinct between the NP and AF analogs of the tissue-engineered IVDs while also being more homogeneous within each region. Most importantly, unlike both MSC groups, discogenic cells deposited little to no collagen X, suggesting that discogenic eDAPS possess a more stable regional phenotype that will be less susceptible to hypertrophy and downstream calcification. Overall, DiscGenics’s discogenic NP and AF cells made compositionally and mechanically superior eDAPS when compared with both human and goat MSCs, with only minor differences between attachment- and suspension-derived discogenic cell eDAPS, supporting their use as a cell source for the creation of human-scale living whole disc replacements.
Impact Statement
This study demonstrates the feasibility of utilizing discogenic cells—a cell product currently undergoing clinical testing—to generate human-scale living whole disc replacements that are compositionally and mechanically superior to mesenchymal stromal cell-seeded constructs. Such a composite engineered disc replacement may offer a motion-preserving alternative to spinal fusion, advancing the standard of care for patients suffering from end-stage disc degeneration and back pain.
Introduction
The societal and economic burdens that arise from chronic back pain are significant, diminishing the quality of life for 600 million people worldwide1,2 and costing high-income countries between 2.5 and 8 billion dollars to diagnose and treat annually. 3 The combined effect of these factors underscores the pressing need for effective strategies to prevent, manage, and mitigate chronic back pain’s impact on patients and society.
The intervertebral discs (IVDs), positioned between the spinal vertebrae, consist of an outer annulus fibrosus (AF) and an inner nucleus pulposus (NP). 4 With aging and physical stress, the discs undergo structural degenerative changes, culminating in reduced disc height, altered spinal biomechanics, and the development of painful conditions, including disc herniation and spinal stenosis, that require treatment.5,6 Patients often undergo spinal fusion surgery when conservative treatments fail to provide relief from back pain, in which the adjacent vertebrae are fused, stabilizing the spine and eliminating motion at the affected disc. 7 In many cases spinal fusion can be effective in providing long-term stability and pain relief for patients. However, it may also lead to the development of adjacent segment disease, as the normal kinematics of the spine are disrupted. 8 Whole disc replacement, also known as artificial disc replacement (ADR), is designed to address the limitations of spinal fusion by replacing the degenerative IVD with an artificial disc implant made from metallic and polymeric components; these implants are designed to articulate or compress, providing pain relief while maintaining spinal mobility.9,10 Concerns from surgeons on the challenge of potential revision cases and the general apprehension regarding the breakdown of the implant and the creation of wear particles have limited the widespread use of these devices compared with spinal fusion surgery.10,11 Critically, ADRs do not fully mimic the native disc or its mechanics and lack the ability to fully integrate with the biological environment of the lumbar spine. 12
Total tissue-engineered IVD replacements are able to support living cells and thus endogenous extracellular matrix (ECM) deposition, offering a potential biologically active alternative to ADR. Our group has developed endplate-modified disc-like angle ply structures (eDAPS) that aim to replicate the biomechanical and biological properties of the native disc by combining a composite biodegradable polymeric biomaterial with a living resident cell population.13,14 By integrating cell therapy with ADR, these disc-like structures aim to restore disc mechanics while also promoting biological maturation of the entire implant and integration with the surrounding vertebrae. Rather than wearing from the time of implantation, such biological ADRs would possess the capacity to respond to the physiological loading environment to maintain tissue structure and function in a more prolonged fashion. Moreover, given that the tissue formed is biological in nature, concerns over wear particles and complications in revision surgery would be minimized.
Previous studies used cells from bovine IVD tissues to seed eDAPS; however, these constructs failed to produce the homogeneous collagen and proteoglycan matrix required for a fully functioning human-scale implant. 15 Mesenchymal stromal cells (MSCs) are a more translationally relevant cell source, but the chondrogenic potential of MSCs, which allows them to act as disc-like cells, varies greatly, depending both on the tissue and donor source. 16 Certain types of MSCs, such as those derived from bone marrow, also show poor phenotypic stability when placed into long-term culture in vitro or in vivo. 17 Indeed, prior in vivo studies of goat bone-marrow-derived MSC eDAPS demonstrate progressive loss of proteoglycan-rich matrix over time postimplantation. 13 An outstanding need remains for a translationally relevant cell source that can be used in human-scale disc tissue engineering.
To address the lack of adequate cell types for ADRs, here we expand upon the eDAPS by incorporating human discogenic cells into the device. Discogenic cells are a promising cell-based treatment for IVD degeneration for regenerating and restoring disc function.18,19 Derived from progenitor stem-like cells, discogenic cells are engineered to differentiate into disc-like cells that can produce ECM components and enhance disc repair. 20 In this study, discogenic cells were produced from both AF and NP human donor-derived disc tissues to create two unique cell populations used to seed the different anatomical regions of the eDAPS. Prior to eDAPS fabrication, discogenic cells were either expanded in two-dimensional attachment culture on tissue culture plastic or in three-dimensional suspension culture in methylcellulose media. These expansion methods may be differentially driving discogenic NP and AF cells toward unique phenotypes, as substrate stiffness and morphology influence the differentiation of MSCs and other stem-like cells.21,22
It is not known how NP and AF discogenic cells will behave in three-dimensional culture, but previous work shows that human NP cells expanded in three-dimensional culture maintain upregulated expression of progenitor surface markers and go on to produce more glycosaminoglycans when differentiated chondrogenically. 23 We hypothesize that discogenic cells expanded in a suspension culture system will be primed to deposit more fibrocartilaginous ECM components in the 3D culture environment of the eDAPS, particularly in the NP analog, as suspension expansion more closely mimics the NP analog’s agarose gel, a material known to support cartilaginous matrix formation in chondrocytes and MSCs. 24
We ultimately hypothesize that by combining both types of discogenic cells within the eDAPS construct, the functional maturation of the eDAPS will be improved through distinct NP/AF cell specialization compared with MSC-derived eDAPS. This approach not only addresses the mechanical needs of the spine but also seeks to promote biological integration and long-term durability of the implant.
Materials and Methods
Harvesting MSCs
Adipose-derived human MSCs (hMSCs) were obtained through ATCC (ATCC PCS-500-011; lot# 80422222) from a single donor (donor characteristics in Supplementary Table S1). Goat MSCs (gMSCs) were harvested from iliac crest bone marrow aspirates of six large-frame, skeletally mature goats. These cells were utilized as a key comparison group for the eDAPS study since our established model for eDAPS in vivo evaluation is implantation in the goat cervical spine, and our historic eDAPS in vitro dataset is built around eDAPS seeded with goat MSCs. Fabrication of eDAPS using goat MSCs allowed us to previously test eDAPS function in a large animal model,13,25 and insights from their in vitro maturation provide critical data to contextualize eDAPS made from a novel human cell source. Bone marrow aspirates were seeded on polystyrene and cultured in basal media (high glucose Dulbecco’s modified Eagle Medium [DMEM] [Invitrogen] supplemented with 10% fetal bovine serum [FBS] [R&D Systems/Bio-Techne S11150] and 1% penicillin/streptomycin/fungizone [PSF] [Antibiotic-Antimycotic; Gibco]) for 2 to 3 weeks to allow the MSCs to attach and wash away nonadherent cells. MSCs were frozen at P1 in FBS with 10% Dimethyl Sulfoxide (DMSO) (Sigma) until needed.
Harvesting discogenic cells
Adult human IVD tissue was procured from three consented organ donors (PD72, PD101, and PD104; donor characteristics in Supplementary Table S1). The tissue was washed and dissected into separate NP and AF regions with transition zones excluded. The tissue was digested using NB5 Collagenase (Nordmark), and cells were seeded at 10,000 cells/cm2 on gelatin-coated T150 flasks (Corning). Expansion of the cells was performed in the presence of DMEM/F12 (Corning) and a proprietary cocktail of other media supplements to promote growth and drive the cells toward a progenitor phenotype, confirmed through screening of the discogenic cells using flow cytometry. Once the confluence of the flasks reached 90%, the cells were detached with TrypLE Express (Gibco) and frozen in FBS (Cytiva) with 10% DMSO (Biolife Solutions). The cells were passaged a second time in attachment culture and subsequently grown out in a 3D suspension culture model using CellStacks with ultralow attachment coating (Corning). A viscous methylcellulose media was used in the suspension culture to generate clusters of cells, 100–200 µm in diameter. The clustered spheres of cells were dissociated into a single-cell suspension using collagenase and were frozen in ProFreeze (Lonza) with 7.5% DMSO. Cells from both the attachment phase (Att.) and the suspension phase (Susp.) were stored in the vapor phase of liquid nitrogen.
Screening hMSCs and discogenic cells
Cell screening using an assortment of analytical assays was performed on hMSCs and human discogenic cells. The hMSCs were expanded for one passage prior to testing. Discogenic cells from both NP and AF tissues were tested from both attachment (Att.) and suspension (Susp.) cultures, for a total of four discogenic subtypes.
The identity and purity of the discogenic cells and hMSCs were measured by flow cytometry using fluorochrome-conjugated mouse antihuman monoclonal antibodies, including appropriate isotype controls. The cells were incubated with antibodies at 4°C for 45 min in Phosphate-Buffered Saline (PBS) with 0.5% human serum albumin, human Fc block, and the following antibody markers previously established in discogenic cells 20 : HLA-DR/DP/DQ, CD24, CD44, CD73, CD90, HLA-ABC, CD34, CD45, CD40, CD271, CD80, and CD86 (BD Biosciences). For each donor, a single sample was processed during staining, with no less than 10,000 events (cells) being collected for analysis from this sample. Positive expression was assessed in the live cell populations by using 7-AAD (BD Biosciences) to exclude dead cells. The flow cytometry measurements were performed on a CytoFLEX Flow Cytometer (Beckman Coulter Life Sciences) and analyzed with FlowJo Software (BD Bioscience).
ECM gene expression of hMSCs and discogenic cells was assessed using RT-PCR. After the secondary 3D suspension culture in methylcellulose media as part of the original harvest process, cells were washed three times and then resuspended in 50 mL of PBS and 1% HSA. A suspension aliquot of 1 mL containing approximately 5 × 105 to 5 × 106 cells was spun down in a microcentrifuge tube. The supernatant was removed, and the cells were lysed with 500 μL of TRIzol before being stored at −80°C. Samples were thawed, and a PureLink RNA kit (Invitrogen) was used to isolate RNA. Reverse transcription and cDNA amplifications were performed in a one-step reaction using TaqMan probes to determine Ct values for ACAN, COL1A2, and COL2A1 with normalized relative expression to the housekeeping gene HPRT1 (Thermo Fisher Scientific). For each donor, a single well was tested for each of the three genes of interest.
ECM protein secretion was measured using ELISAs for Aggrecan (DIAsource), Collagen I (Abcam), and Collagen II (R&D Systems). To prepare for the assays, the cells were seeded into a polypropylene 96-well V-bottom plate at 2.5 × 105 cells/well in DMEM high glucose media with pyruvate, Insulin/Transferrin/Selenious (ITS)+ Premix,
Tissue-engineered intervertebral disc fabrication, culture, and characterization
Endplate-modified Disc-like Angle Ply Structures (eDAPS) were fabricated as previously described (W:16 mm × H:9 mm).13,15,26 The eDAPS were seeded with a combination of different cell types to compare their functional maturation:
The eDAPS were cultured at standard culture conditions for 10 weeks with constant mechanical agitation in chemically defined chondrogenic media (high glucose DMEM supplemented with 1% PSF, 0.1 mM dexamethasone, 50 μg/mL Ascorbate-2-phosphate, 40 μg/mL
NP and AF components of these eDAPS were then separated and submerged in 2 mL of 0.5% proteinase K solution for enzymatic digestion at 60°C overnight (∼16 h). Digestion solutions were used to run biochemical assays for DNA content, glycosaminoglycan (GAG) content, and collagen content using the Quant-iTTM PicoGreenTM dsDNA Assay Kit (Invitrogen P7589), dimethylmethylene blue assay, and hydroxyproline assay, respectively (n = 3). 29
The remaining eDAPS from each group were fixed in 10% buffered formalin and processed for paraffin histology and immunofluorescence (n = 2 to 3/group). Paraffin sections were stained with Alcian blue, picrosirius red, and hematoxylin/eosin. Briefly, immunofluorescence staining for collagen I (DSHB 8-3A5; 4 μg/mL; developed by A. L. De Blas), collagen II (DSHB II-II6b3, 4 μg/mL; developed by T. F. Linsenmayer), collagen X (DSHB X-AC9, 4 μg/mL; developed by T. F. Linsenmayer), and chondroitin sulfate (DSHB 9BA12, 4 μg/mL; developed by W. M. Halfter) was performed by serially rehydrating paraffin sections, digesting samples in Proteinase K (Dako S3020) for 5 min at room temperature, and treating them with Background Buster (Innovex NB306) for 30 min at room temperature, after which samples were incubated in primary antibodies overnight at 4°C and then with fluorescent secondaries for 1 h the next day (Invitrogen A11032, Invitrogen A11008). Before proteinase K digestion, sections used for chondroitin sulfate staining were incubated in hyaluronidase (Sigma H3884; 18.675 units per sample) for 15 min at room temperature.
Statistical analysis
Data in Table 1, as well as in Figures 1, 2, and 3, were analyzed using a two-way ANOVA with Tukey’s post hoc test, where cell type (NP, AF) and culture condition (attachment, suspension) were the two factors compared. In these analyses, hMSCs were excluded due to low n number. Mechanics data in Figure 4I–M were analyzed using a nonparametric one-way ANOVA with a Kruskal–Wallis post hoc test, where eDAPS experimental group (gMSC, hMSC, discogenic NP Att., discogenic NP Susp.) was compared. All remaining data in Figures 4 and 5 were analyzed using a two-way ANOVA with Tukey’s post hoc test, where eDAPS experimental group (gMSC, hMSC, discogenic NP Att., discogenic NP Susp.) and eDAPS tissue compartment (NP, AF) were the two factors compared. Significance was defined as p < 0.05.
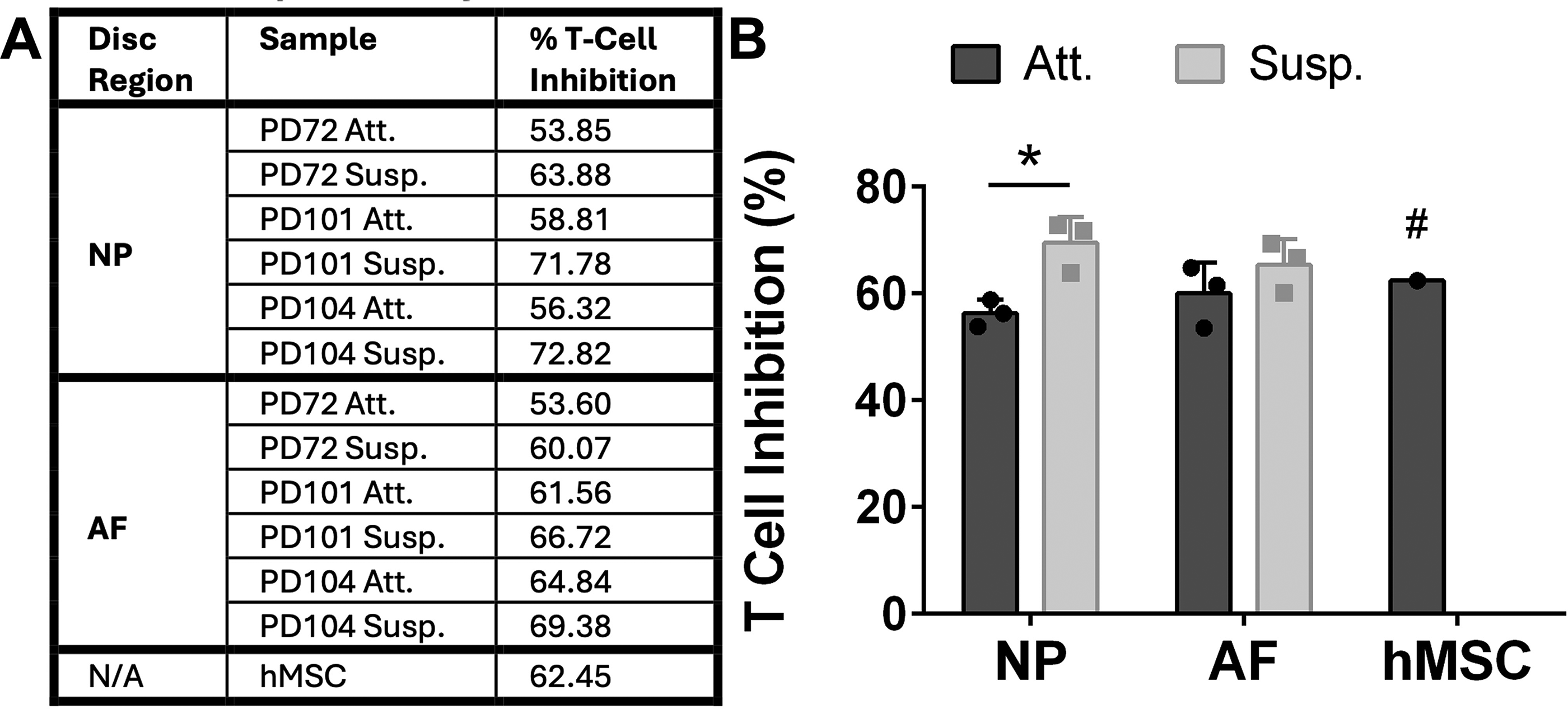
Cellular immunopotency.
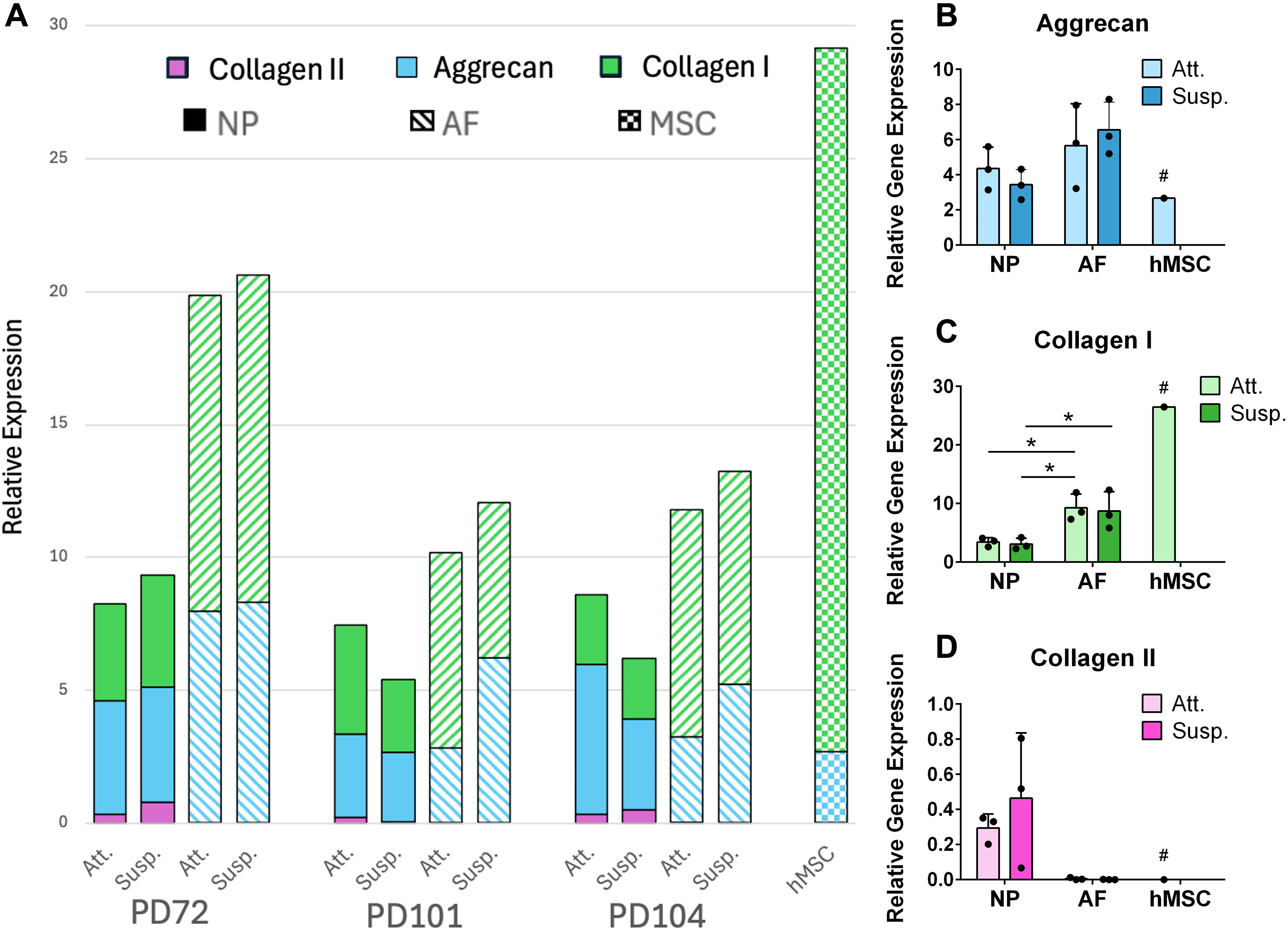
Relative gene expression. The genes for aggrecan (turquoise), collagen I (green), and collagen II (pink) were measured for relative expression against the human housekeeping gene HPRT1.
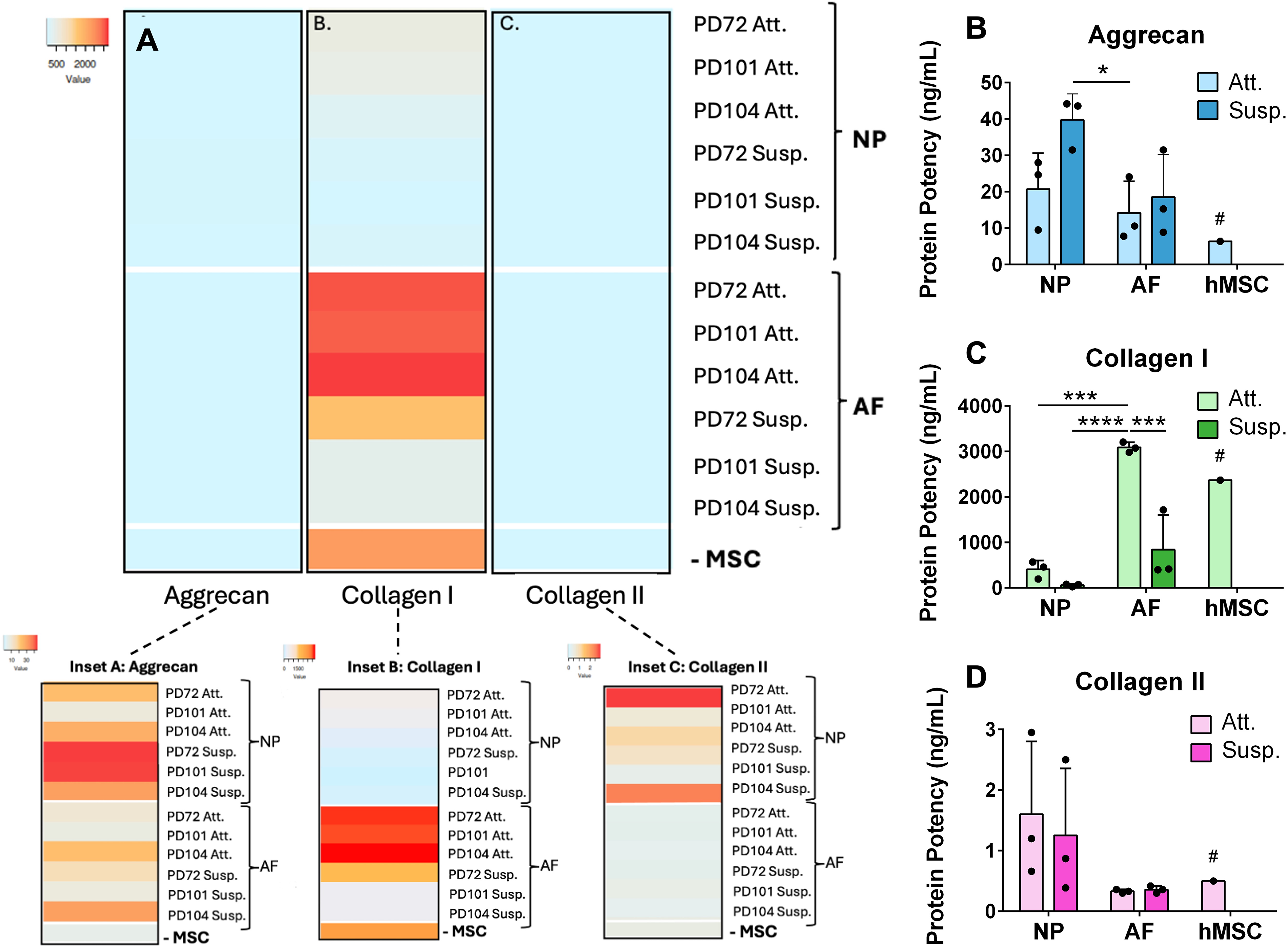
Secretion of extracellular matrix proteins.
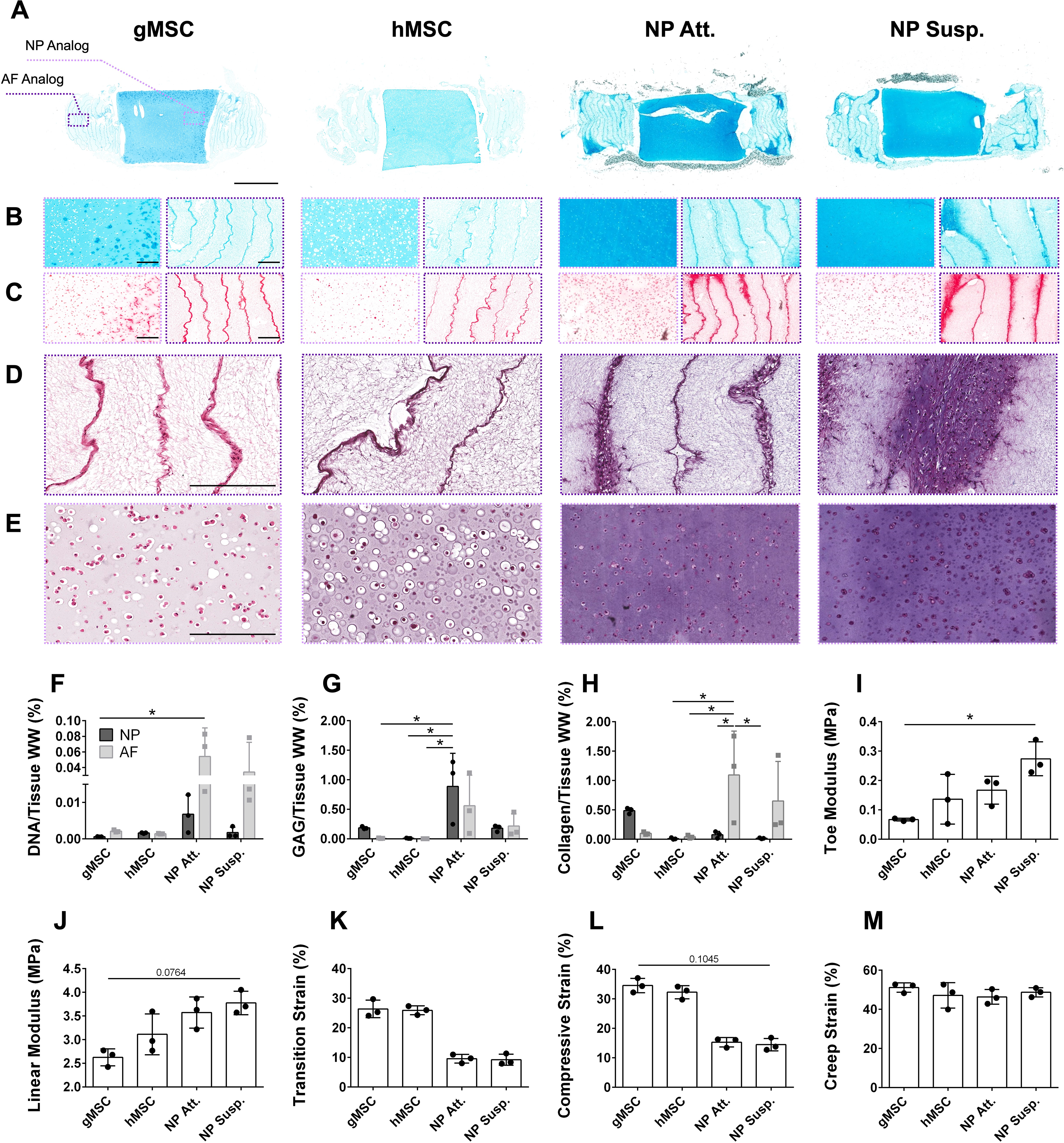
eDAPS histology, biochemical composition, and mechanics.
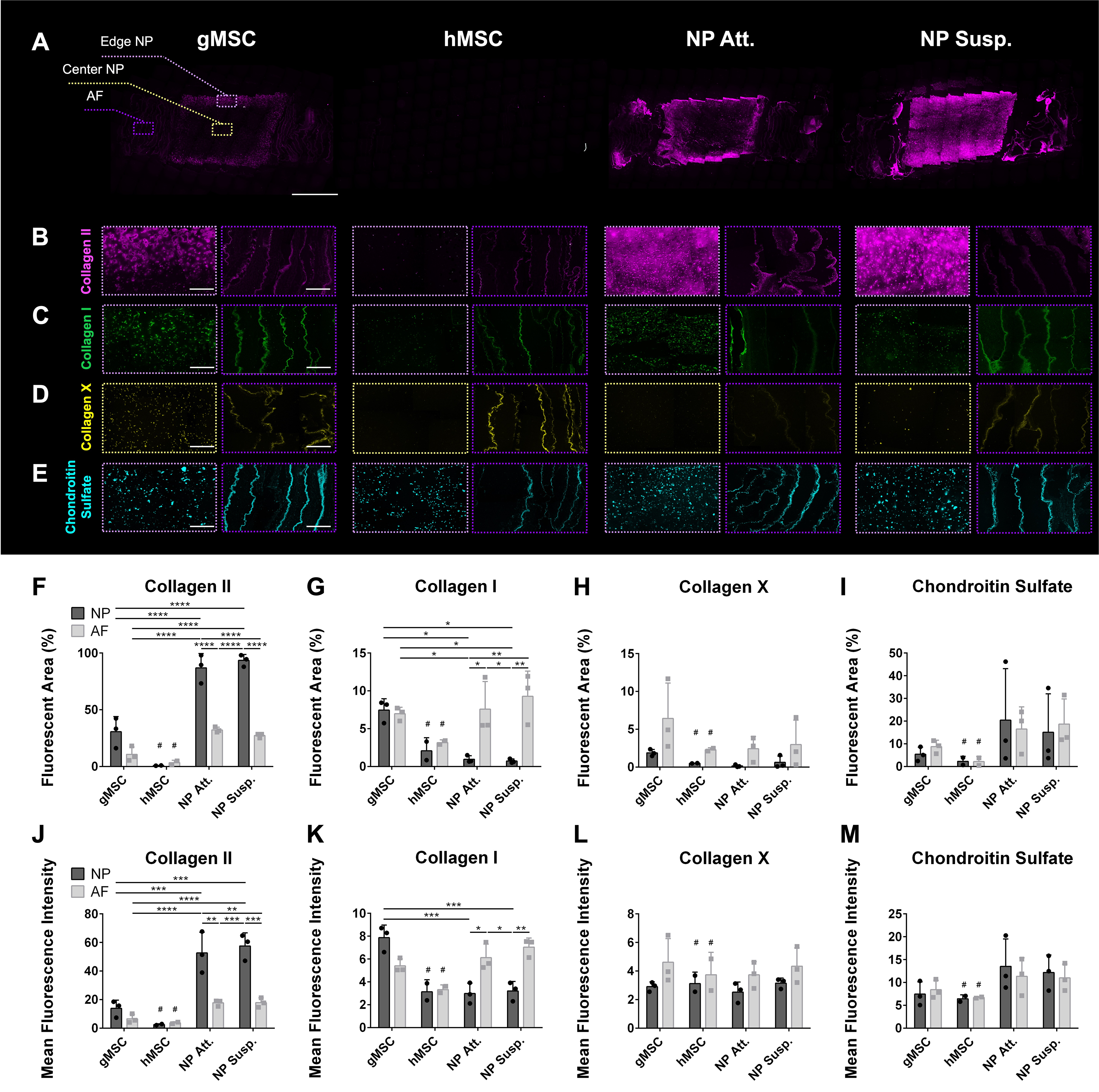
Immunofluorescence of eDAPS for detection of extracellular matrix components. IF staining of collagen II (pink) in
Discogenic Cell and Human MSC Surface Markers
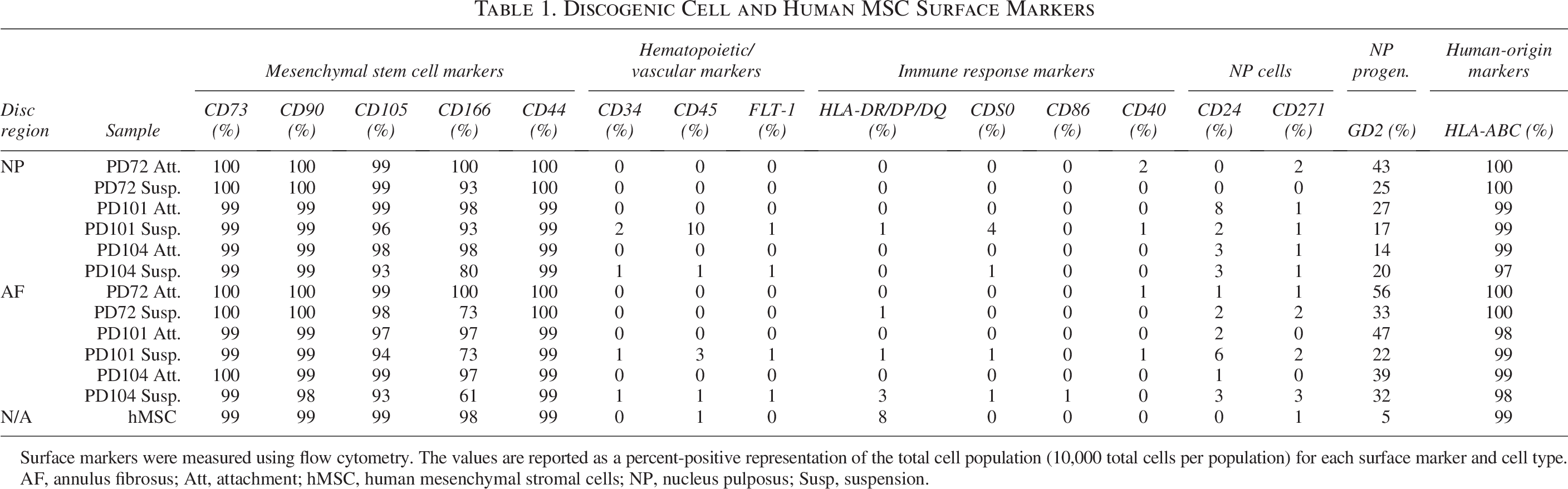
Surface markers were measured using flow cytometry. The values are reported as a percent-positive representation of the total cell population (10,000 total cells per population) for each surface marker and cell type.
AF, annulus fibrosus; Att, attachment; hMSC, human mesenchymal stromal cells; NP, nucleus pulposus; Susp, suspension.
Results
All four of the discogenic subtypes (NP Att., NP Susp., AF Att., and AF Susp.) and the hMSCs were above 97% positive for the stem cell identity markers CD73, CD90, and HLA-ABC. The hMSCs minimally expressed HLA-DR/DP/DQ (8%), whereas the discogenic cells expressed levels no higher than 3%. A single discogenic cell donor (PD101) expressed CD24 at 8% in the NP Att. cells and at 6% in the AF Susp. cells. All other donors, including the hMSCs, expressed CD24 at 3% or lower. GD2 expression ranged from 14% to 56% in the discogenic cells, with expression levels being both significantly higher in the AF than in the NP (p = 0.0310) and significantly higher in the Susp. than in the Att. cultures (p = 0.0417). In comparison, the expression level for GD2 in the hMSCs was 5%. CD166 expression ranged from 61% to 100% in the discogenic cells, with expression significantly higher in the NP versus the AF (p = 0.0097) and higher in Att. versus Susp. cultured cells (p = 0.0002). The hMSCs were 98% positive for CD166. The expression levels for CD105 were 93% or greater in all cell populations, and the expression levels for CD44 were 99% or greater in all cell populations. The markers for CD34, CD271, FLT-1, CD45, CD40, CD80, and CD86 were below 3% expression for all cells. The data table for the identity and purity surface markers is summarized in Table 1; GD2 and CD166 were the only markers for which significant differences were detected. The immunopotency potential of hMSCs was equivalent to that of the discogenic cells, and the immune suppression capability of discogenic cells was increased in cells grown in Susp. (p = 0.0497 for NP Att. vs. NP Susp.; p = ns for AF Att. vs.AF Susp.) (Fig. 1).
Quantification of gene expression for matrix proteins (Fig. 2; Supplementary Fig. S2 for full dataset) revealed low expression levels of collagen II in all cell types. Although not significant, expression of collagen II was on average 99% higher in the NP when compared with the AF. There was no detectable collagen II gene expression in the hMSCs. Collagen I gene expression was 64% higher in the AF compared with the NP (p = 0.0397 for AF Att. vs. NP Att.; p = 0.0297 for AF Att. vs. NP Susp.; p = 0.0457 for AF Susp. vs. NP Susp.). Out of all cell types, collagen I was expressed most highly in the hMSCs. There was no statistical difference found in aggrecan gene expression between the NP and AF discogenic cells. Likewise, no statistically significant differences were observed in gene expression between Att. and Susp. cultured discogenic cells within each cell type for any of the measured matrix proteins.
When ECM protein secretion was measured using ELISAs, similar trends to gene expression were observed, with the exception of aggrecan secretion (Fig. 3A; Supplementary Fig. S3 for full dataset). After 15 days of culture, NP cells synthesized more aggrecan than AF cells did (p = 0.0434 for NP Susp. vs. AF Att.) (Fig. 3B). Collagen I synthesis followed the gene expression trends; production was higher in AF cells when compared with NP cells, with Att. cells secreting more collagen I than Susp. cells (p = 0.0001 for AF Att. vs. NP Att.; p < 0.0001 for AF Att. vs. NP Susp.; p = 0.0005 for AF Att. vs. AF Susp.) (Fig. 3C). Collagen I secreted by hMSCs was comparable with that secreted by AF Att. cells. Similar to the gene expression for collagen II, low levels were observed across all cell types. There was a trend toward higher collagen II secretion in the NP versus the AF discogenic cells; however, this did not reach statistical significance (Fig. 3D). AF cells and hMSCs secreted collagen II at similar levels.
Tissue-engineered discs cultured with discogenic cells were histologically, biochemically, and mechanically similar to each other; discogenic eDAPS proved superior in all three categories to gMSC- and hMSC-seeded eDAPS. Discogenic cells deposited more proteoglycan in both the NP and AF analogs than did the best-performing gMSCs (Supplementary Fig. S4) or hMSCs (Fig. 4A and B). In addition, proteoglycan and collagen deposition was more homogeneous in discogenic eDAPS overall (Supplementary Fig. S5). The goat MSCs, in particular, exhibited strong staining for collagen mainly in the outer 500 μm of the NP region, which was not observed in any other group (Fig. 4C). Discogenic AF cells deposited a greater quantity of collagen between the layers of electrospun polycaprolactone (PCL) fibers comprising the AF region of the eDAPS (Fig. 4C). This collagen penetrated more deeply into the polymeric sheets of the AF, attributable to the discogenic AF cells’ migration into the PCL, whereas both gMSCs and hMSCs remained confined to the interlamellar spaces (Fig. 4D; Supplementary Fig. S6). Discogenic NP cells were distinctly smaller in size and occupied smaller pericellular spaces than did the MSCs (Fig. 4E; Supplementary Fig. S7).
These histological trends were supported by biochemical analyses. DNA content was higher in NP Att. and NP Susp. eDAPS, most prominently in the AF regions when compared with MSC-seeded eDAPS (Fig. 4F). GAG deposition was significantly higher in the NP of NP Att. eDAPS when compared with the AF of gMSC eDAPS (p = 0.0248), the AF of hMSC eDAPS (p = 0.0232), and the NP of hMSC eDAPS (p = 0.0248), although GAG levels were also elevated in the AF and in NP Susp. eDAPS (Fig. 4G). Collagen content was elevated in the AF of both discogenic-seeded eDAPS as well as in the NP region of gMSC-seeded eDAPS, closely mimicking the histological findings (Fig. 4H). Increased fibrocartilaginous matrix deposition led to increased toe modulus (Fig. 4I), increased linear modulus (Fig. 4J), decreased transition strain (Fig. 4K), and decreased compressive strain (Fig. 4L) of discogenic eDAPS compared with both gMSC and hMSC constructs. All eDAPS experienced similar creep deformation (Fig. 4M; see Supplementary Fig. S8 for creep behavior).
Immunohistochemistry (Supplementary Fig. S9) confirmed that the vast majority of all collagen produced by all cell types was collagen II (Fig. 5A and B, 5F, 5J; Supplementary Fig. S10). Collagen I was also deposited, albeit to a lesser extent, by all discogenic cells, as well as goat MSCs, in both NP and AF analog regions (Fig. 5C, G, K; Supplementary Fig. S11). While goat MSCs produced similar levels of collagen I in the NP and AF analogs, discogenic AF cells deposited significantly more collagen I than discogenic NP cells (for AF of Att. eDAPS vs. NP of Att. eDAPS: Fluorescent area p = 0.0258; Mean Fluorescence Intensity (MFI) p = 0.0134; for AF of Susp. eDAPS vs. NP of Susp. eDAPS: Fluorescent area p = 0.0041; MFI p = 0.0029), and discogenic NP cells deposited significantly more collagen II than discogenic AF cells (for NP of Att. eDAPS vs. AF of Att. eDAPS: p < 0.0001; MFI p = 0.0013; for NP of Susp. eDAPS vs. AF of Susp. eDAPS: Fluorescent area: p < 0.0001; MFI p = 0.0004). The hMSCs deposited scant amounts of collagen I and II overall, especially in the NP region. Both gMSCs and hMSCs secreted collagen X throughout the constructs (Fig. 5D, H, L). In contrast, collagen X deposition was muted from discogenic AF cells and virtually nonexistent from discogenic NP cells (Supplementary Fig. S12). All cell types made chondroitin sulfate at significant levels (Fig. 5E, 5I, 5M; Supplementary Fig. S13), but differences between groups were less pronounced in comparison to differences in collagen I and II levels.
Discussion/Conclusion
This study demonstrated the successful use of DiscGenics’ discogenic NP and AF cell products in a tissue-engineered total IVD and marks the first instance in which human-sized eDAPS were fabricated using a scalable human IVD cell source. Our data suggest that discogenic cells may be a superior alternative to MSCs in the eDAPS construct.
Adipose-derived hMSCs are widely used in cell therapy applications due to their abundance, ease of isolation, differentiation potential, and resilience in short-term culture. However, several limitations hinder their application in tissue engineering. Populations of MSCs are heterogeneous, both between donors, 30 and between subpopulations within each donor. 31 Critically, these variations lead to inconsistent therapeutic outcomes in a clinical setting, many of which remain unreported. 32 The gene expression and ECM secretion profiles of hMSCs also make them less advantageous for disc tissue engineering. Our data demonstrate that matrix production of hMCSs is dominated by collagen I, with discogenic cells producing more collagen II and aggrecan compared with hMSCs. Taken together, these challenges render hMSCs suboptimal for large-scale manufacturing operations where consistency and fibrocartilaginous matrix production are crucial.
Discogenic NP cells previously treated IVD degeneration in a recently completed randomized, double-blinded, placebo-controlled phase I/II study. 19 Analytical testing of discogenic NP cells here matched previous findings reported by DiscGenics,18,20 adding support to the characterization profile of these cells, in addition to providing insight into the properties of AF discogenic cells, which have not been previously characterized. Flow cytometry showed that discogenic AF cells have higher GD2 expression and lower CD166 expression compared with discogenic NP cells. The elevated presence of CD24 in both NP and AF tissues from donor PD101 is likely due to the natural donor-to-donor variability in this marker related to factors such as age, obesity, or immune system activation.33–36 Protein and gene expression assays confirmed that discogenic AF and NP cells produced matrix reflective of their disc microenvironments, with discogenic AF cells producing more collagen I and discogenic NP cells producing more aggrecan and collagen II, revealing the unique properties of each cell type.
Before eDAPS seeding, discogenic AF and NP cells cultured in suspension initially produced greater amounts of aggrecan than cells cultured via attachment. However, eDAPS seeded with Susp. or Att. cultured cells were similar in every outcome measured. It is likely that Susp. and Att. cells performed similarly once cultured within the NP hydrogel, as NP cells expanded in monolayer developed normal NP phenotypes following 3D culture. 37 In addition, our data demonstrate that the NP and AF regions of discogenic eDAPS are phenotypically distinct, with discogenic AF cells producing more collagen I and discogenic NP cells producing more collagen II. In contrast, the NP and AF regions of MSC-seeded eDAPS matured similarly, comparable to eDAPS seeded with primary bovine AF and NP cells in our prior work. 15 The NP and AF regions of the eDAPS received the same culture media, so the regional distinction observed in discogenic eDAPS was driven solely by the NP and AF progenitor cells themselves and their interactions with their discrete substrates and not by independent chemical signals.
The lack of regional distinction achieved in the MSC-seeded eDAPS is defined by the collagen-rich outer edge of the NP, indicating that MSCs struggled to deposit matrix when nutritionally challenged at the center of the eDAPS. Unfavorable diffusion gradients often create hostile environments (low glucose and oxygen) for cells toward the center of large implants that result in cell apoptosis and heterogeneous matrix deposition,38,39 as observed in the MSC-seeded eDAPS here. Preconditioning MSCs could prolong their survival in hypoxic, nutrient-deficient environments but may not overcome the heterogeneity inherent to MSC populations, as the effect of preconditioning also seems to be donor-dependent. 40 Conversely, proteoglycan and collagen deposition was both more homogeneous and more pronounced in the NP of discogenic eDAPS, indicating that discogenic cells more readily thrived in the unfavorable environment of a human-sized disc even when compared with other native disc cells. 15
It is well known that MSCs, both adipose- and bone-marrow-derived, produce collagen X, a marker of chondrogenic hypertrophy, when exposed to chondrogenic growth factors. 41 While MSCs from both human and goat sources deposited collagen X throughout the eDAPS, discogenic cells’ deposition of collagen X was minimal or nonexistent. The presence of collagen X and the resulting consequences of osteogenesis and subsequent mineralization following in vivo implantation 42 are undesirable for a tissue-engineered IVD replacement where the maturing tissue needs to remain fibrocartilaginous. The absence of collagen X in the discogenic eDAPS further bolsters the superiority of these cells in the development of an eDAPS able to successfully function in vivo.
This study was limited by its low sample size. Human-sized tissue-engineered constructs require tens of millions of cells to seed, and the number of samples was ultimately constrained by our ability to successfully expand adipose-derived hMSCs to the scale required as well as by the high cost involved in producing DiscGenics’ cell products. Future work will focus on challenging discogenic eDAPS via exposure to hypoxic, low glucose, and inflammatory conditions in vitro and in vivo to better understand how these tissue-engineered IVD replacements fare in more physiologically relevant environments. Overall, our data demonstrate the feasibility of utilizing discogenic cells—a cell product currently under clinical investigation—to generate human-scale living whole disc replacements that are compositionally and functionally superior to MSC-seeded constructs.
Authors’ Contributions
Study conception and design: S.E.G., R.L.M., H.E.S., and I.E. Investigation: S.S.F., M.F., G.D., R.R., and A.B. Obtaining of Funding: S.E.G., R.L.M., and H.E.S. Visualization: S.S.F. and M.F. Drafting of the article: S.S.F., M.F., G.D., and S.E.G. Revising the article for critically important intellectual content: all authors. Final approval of the version to be submitted: all authors.
Footnotes
Acknowledgments
Special thanks to Dr. DongHwa Kim for fabricating the electrospun PCL scaffolds used in this study and to DonorConnect Salt Lake City, Utah, for supplying the human tissue used to isolate Discogenic NP and AF cells.
Funding Information
The research was partially funded by DiscGenics, which may stand to gain from the findings through the advancement of new products and the improvement of its existing product line. This work was supported by the Department of Veterans Affairs Rehabilitation Research and Development Service, award numbers I01 RX002274 and IK6 RX003416, as well as a BRAVE Award from the VA Technology Transfer Program. This work was also carried out in part with support from the Penn Center for Musculoskeletal Disorders (NIH/NIAMS P30AR069619).
Data Availability
Disclaimer
The authors affirm that the conclusions drawn in this article are based solely on the research conducted and are independent of any external influence. The contents do not represent the views of the U.S. Department of Veterans Affairs or the U.S. Government.
Disclosure Statement
Discogenic cells were provided for the eDAPS studies under a Cooperative Research and Development Agreement between DiscGenics, Inc. and the Corporal Michael J. Crescenz VA Medical Center.
Employees of DiscGenics contributed to the study’s design, data collection, analysis, interpretation, and article preparation. In addition, the company supported the submission of the article.
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.