
Abstract
BACKGROUND:
Biological hydroxyapatite (BHA)-based bone grafting materials have been widely used for bone regeneration in implant surgery. Much effort has been made in the improvement of their osteogenic property as it remains unsatisfactory for clinical use. Osteoimmunomodulation plays a significant role in bone regeneration, which is highly related to active inorganic ions. Therefore, attempts have been made to obtain osteoimmunomodulatory BHA-based bone grafting materials with optimized osteogenic property by ion doping.
OBJECTIVE:
To summarize and discuss the active inorganic ions doped into BHA and their effects on BHA-based bone grafting materials.
METHOD:
A literature search was performed in databases including Google Scholar, Web of Science and PubMed, with the elementary keywords of “ion doped” and “biological hydroxyapatite”, as well as several supplementary keywords. All document types were included in this search. The searching period and language were not limited and kept updated to 2022.
RESULTS:
A total of 32 articles were finally included, of which 32 discussed the physiochemical properties of BHA-based biomaterials, while 12 investigated their biological features in vitro, and only three examined their biological performance in vivo. Various ions were doped into BHA, including fluoride, zinc, magnesium and lithium. Such ions improved the biological performance of BHA-based biomaterials, which was attributed to their osteoimmunomodulatory effect.
CONCLUSION:
The doping of active inorganic ions is a reliable strategy to endow BHA-based biomaterials with osteoimmunomodulatory property and promote bone regeneration. Further studies are still in need to explore more ions and their effects in the crosstalk between the skeletal and immune systems.
Keywords
Introduction
Bone defect caused by trauma, disease, surgery, and congenital malformations remains a major health problem worldwide [1]. Currently, the common approaches for the rehabilitation of bone defect depend on the use of either autologous bone or bone substitutes, such as allogenic and xenogeneic bone grafts as well as synthetic materials [1–3]. In implant surgery, biological hydroxyapatite (BHA) has been the most widely used xenogeneic grafting material for bone regeneration for years. However, its osteogenic property remains unsatisfactory and further improvement is of utmost importance. Recently, osteoimmunomodulation has become an attractive new strategy for developing advanced bone grafting materials. It emphasizes that biomaterials should be endowed with immunomodulatory capacity to modulate the local immune environment so as to favor the osteogenesis and osseointegration of implants [4]. It is also found that active inorganic ions are highly involved in osteoimmunomodulation. Therefore, as a safe, effective and cost-saving approach, the doping of various kinds of inorganic ions into BHA has been tried, with the attempt of enhancing its osteogenic property.
For a comprehensive understanding of the research status of ion doped BHA, literature searches have been done in databases including Google Scholar, Web of Science and PubMed, with the elementary keywords of “ion doped” and “biogenic hydroxyapatite”, and several supplementary keywords, including “ion doping”, “ion incorporated”, “ionic incorporation” and “biological apatite”, “biological hydroxyapatite”, “bovine bone” and “porcine bone”. All document types were included in the searches, and the searching period and language were not limited. A total of 1346 articles were found, out of which 121 related articles were filtered further based on their titles and abstracts. Finally, 32 articles focusing on ion-doped biological hydroxyapatite as xenogeneic bone grafting materials (or biomaterials in medication) were included, in which different aspects of BHA were investigated (32 for physiochemical properties, 12 for biological features in vitro and three for biological performance in vivo) (Table 1).
Research progress of ion-doped biological hydroxyapatite
Research progress of ion-doped biological hydroxyapatite
BHA is a carbonated, non-stoichiometric Ca-deficient hydroxyapatite, which is different from synthetic hydroxyapatite (HA) in composition, stoichiometry, crystal size, morphology, crystallinity degree, degradation rate, and overall biological performance [5–7]. It is usually extracted from biological sources or wastes [8–15] (Table 2).
Origins of biological hydroxyapatite
Origins of biological hydroxyapatite
Yet, as a xenogeneic bone graft, the clinical performance of BHA is not satisfactory due to the lack of osteoinductivity [16,17]. Therefore, many attempts have been made to incorporate bioactive factors into BHA to make it osteoinductive [18–24]. Since 1991, initial histological evidence and further proof in vivo have indicated that, optimizing the physicochemical properties of biomaterials could endow them with osteoinductive ability rather than delivering proteins or cells [25–27]. Represented by trace elements and ion doping, the delivery of inorganic factors could not only optimize the properties of target materials, but also show lower cost and easier storage, compared with the incorporation of organic factors such as drugs, fibrins, cytokines, growth factors, etc. [17,28–30]. The existence of these incorporated ions may affect the crystallization, mechanical properties, degradation, and biological activity of BHA, and consequently affect its physiological function in vivo, such as the development of teeth and bone, whereas their absence could lead to bone loss or fragility [5]. Such naturally incorporated ions in BAH indicate the feasibility of foreign ion doping due to the tolerance of its crystal lattice. In BHA, OH− can be easily replaced by a small amount of F− and CO3 2−, while Ca2+ can be replaced by Sr2+, Mg2+ and Zn2+ [31–33]. The domino effect imposed on the finest levels of BHA structure by doping with foreign ions is to illustrate the potential of this approach to affect a number of microstructural parameters and, thus, properties exhibited by the material [34–39].
According to osteoimmunomodulation, bone regeneration is a complex dynamic remodeling process which depends on both the biological behavior of osteoblastic lineage and the coordination and regulation of immune and cardiovascular systems. As the representative immune cell in bone regeneration, macrophage has been found to be involved in the regulation of bone homeostasis. Its biological functions are heavily influenced by inorganic ions through some molecular signaling pathways, including but not limited to the toll-like receptors (TLRs), the mitogen-activated protein kinase (MAPK), the Wnt and the transient receptor potential cation channel member 7 (TRPM7), as well as the downstream signaling pathways intranuclear like nuclear factor-kappa B (NF-κB) (Fig. 1).
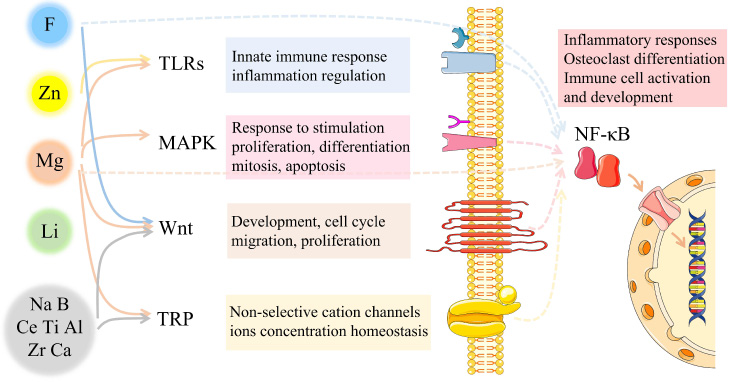
Potential molecular mechanism of osteoimmunomodulation caused by ion incorporation. Immune cells, especially macrophages, play a key role in bone homeostasis. Bioactive ions are closely related to both the bone remodeling process and the immune system. There are several bioactive ions related molecular signaling pathways that tightly link the bone regeneration process with the immune system.
In mammals, TLRs family are expressed on antigen presenting cells such as dendritic cells and macrophages, serving as key pattern recognition receptors (PRR) with central roles to sense diverse pathogen-associated molecular patterns (PAMPs) [40]. TLRs normally react to foreign implants and further stimulate the expression of inflammatory cytokine genes. However, some specific bioactive ions such as Zn2+ and Mg2+ could inhibit the expression of TLRs signaling pathway instead [41,42].
Consisting of famous ERK, JNK and p38 kinase families, MAPK cascades play a crucial role in the transduction of extracellular signals to cellular responses, the reaction to oxidation stress and inflammation stimulation [43]. By regulating the secretion of inflammatory factors, bioactive ions like Mg2+ could indirectly affect MAPK signaling pathway to further control the proliferation, differentiation and apoptosis of osteo lineage [44].
Wnt proteins constitute a large family of cysteine-rich secreted glycoproteins, which could activate at least three different signaling pathways: the canonical pathway, the planar cell polarity pathway, and the Wnt/Ca2+ pathway [45]. Bioactive ions such as F− and Mg2+ are related to the expression of canonical Wnt/β-catenin signaling, which can affect the entire osteoblastic lineage to control the bone remodeling process further [46,47].
TRPM7 is one of the non-selective cation TRP channels, which mainly controls the influx of Ca2+ and Mg2+. Located in the surface of macrophages and bone mesenchymal cells, TRPM7 senses mechanical stimulation and mediates cations influx to regulate inflammation reaction and bone regeneration consequently [48,49].
NF-κB is a transcription factor found in all nucleated cell types. It is a vital component of the molecular processes such as the development and activation of immune cells, inflammatory responses, and osteoclast differentiation. It was reported that F− and Mg2+ could affect the expression of NF-κB signaling pathway both in macrophage and bone lineage, to regulate inflammation and bone remodeling, respectively. [49,50]. Currently, NF-κB signaling pathway is the most evident intranuclear signaling pathway in osteoimmunomodulation.
Based upon the literature, for every single ion, its effect on the osteoimmunomodulation of macrophages may involve more than one signaling pathway mentioned above, either fully or partially. Besides, some parts of signaling pathways remain unclear. Therefore, more work is needed to clarify the underlying mechanism of active ion-doping caused osteoimmunomodulation.
Fluoride
Fluoride (F) is one of the vital trace nutrient elements for human health. Through thermal treatment with hydrofluoric acid, F− was doped into BHA from bovine bone, resulting in several crystal structural changes including increased crystallinity and decreased crystal size [51]. F− could substitute CO3 2− in the HA crystal lattice to make it more resistant to acid [52]. Further investigation has proven that, the doping of F− may not only promote the crystallization of BHA (Fig. 2, [53]), but also enhance its osteogenic capacity in vitro and in vivo in a concentration-dependent manner [46,50,53–57].

The change of hydroxyapatite crystal structure by fluoride incorporation. SEM images: (a, b) FPHA (test group, F−: 1.0 mol/L, immersion period: 24 h, heat treatment); (c, d) PHA (heat treatment); (e, f) PHA (blank control) under 500× and 20,000× magnifications. Reproduced from ref. [53] with permission from The Japanese Society for Dental Materials and Devices, copyright 2012.
In previous studies, fluoride doped porcine bone (F-PHA) was prepared successfully by immersing PHA in sodium fluoride solution (NaF) with gradient concentrations and subsequent thermal treatment [50,53]. It was found that structural and functional changes of PHA were caused by the doping of F−. Specifically, the spheroid-like crystals of PHA were changed into rod-like after F− doping, resulting in F− concentration-dependent increased compressive strength, as well as decreased porosity and solubility [56]. In addition, it has also been found that F− could enhance the synthesis of bone-associated glycoproteins in bone mesenchymal stem cells (BMSCs), thereby promoting cell adhesion, protein adsorption and ALP activity, leading to the increase of mineral deposition in vitro [46,57].
Also, such effects were dose-dependent with F− [50 ]. In the low concentration (0.24–24 μM) of F−, in vivo vascularized bone formation can be achieved via finetuning the F− concentration, while the peak osteogenic effect was found at 2.4–24 μM [50]. On the basis of previous finding, animal experiments in SD rats and beagle dogs have been carried out to investigate the performance of F-PHA in bone augmentation [58]. 12 weeks later, the F-PHA groups showed a significantly higher alveolar ridge width than the PHA group on the macroscopic level [58]. Histologically, it was observed that F-PHA particles were surrounded by newly formed bone, where the proportion of non-mineralized tissue was significantly lower than that in the PHA group [58].
Further investigation revealed the molecular mechanism of F-PHA in osteoimmunomodulation. On the one hand, osteogenic activities of F-PHA may be associated with the activation of canonical Wnt/β-catenin signaling pathway, as well as the inhibition of receptor activator of nuclear factor-κB ligand/osteoprotegerin (RANKL/OPG) signaling pathway [46]. Moreover, F− was found to elicit a significant osteoimmunomodulatory effect in the modulation of inflammatory cytokines, and the expression of osteogenic factors such as BMP2, OSM, spermine and spermidine, as well as angiogenic factors including VEGF and IGF-1 during the early response [50], leading to more new bone formation in vivo [46] (Fig. 3).
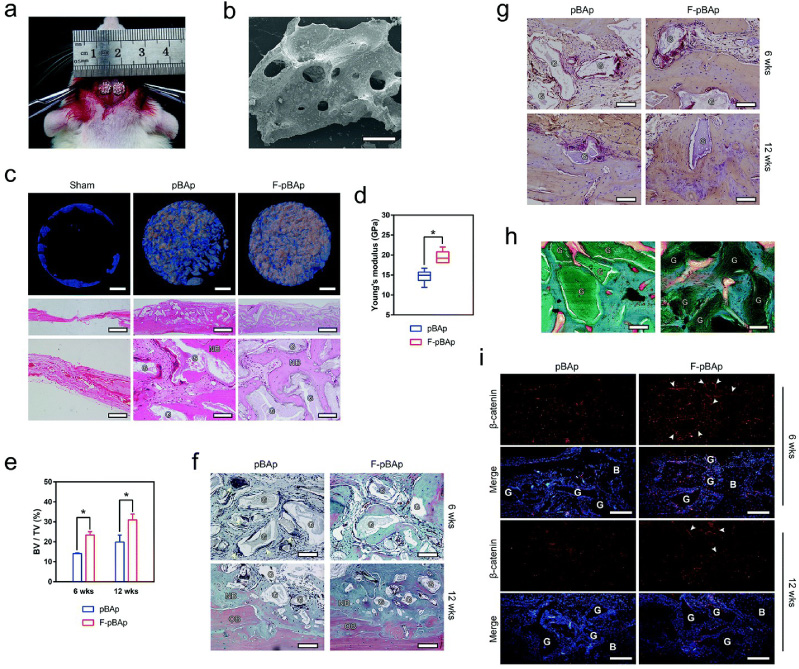
Fluoride incorporation leading to new bone formation in vivo. (a) The 5 mm diameter calvarial defects in rats grafted with pBAp (left) and F-pBAp (right). (b) Representative SEM image showing the grafted xenograft granules (scale bar = 300 μm). (c) Representative reconstructed micro-CT images (scale bar = 1 mm) showing bone growth (blue) around the grafted materials (white), as well as the H&E staining histology showing the ingrowth of bone tissues (low magnification: scale bar = 100 μm; high magnification: scale bar = 1 mm; G: grafted materials; NB: new bone). (d) Young’s moduli of newly formed bone round grafted pBAp or F-pBAp at week 12 (significant difference between groups was indicated as ∗ P < 0.05). (e) Quantification of the bone volume (BV)/total volume (TV) of the calvarial defect area grafted with pBAp or F-pBAp at week six and week 12 (significant difference between groups was indicated as ∗ P < 0.05). (f) Representative Goldner’s trichrome staining of the grafted areas at week six and week 12 showing the growth of new bone around the grafted materials (scale bar = 100 μm). (g) Representative TRAP staining showing osteoclastogenesis around the grafted pBAp and F-pBAp (scale bar = 100 μm). (h) Representative Goldner’s trichrome staining of the undecalcified tissue confirming the direct contact between the new bone and the grafted materials (scale bar = 100 μm). (i) Fluorescent IHC images showing the activation of β-catenin around the grafted materials (scale bar =200 μm). Reproduced from ref. [46] with permission from the Royal Society of Chemistry, copyright 2018.
On the other hand, F− participated in immune response and inflammation regulation in a dose-dependent manner. In detail, the low concentration of F− promoted the polarization of macrophages to the M2 phenotype by increasing the expression of IκB-𝛼 (NF-κB inhibitor 𝛼) to inhibit nuclear factor kappa light chain enhancer of NF-κB signaling pathway [50]. This was demonstrated by the increased expression of cellular surface markers including arginase and CD11c, as well as the decreased expression of CCR7 and iNOS [50]. Whereas, the high concentration of F− over 24 μM showed evident cytotoxicity, inducing the secretion of inflammation factors including IL-1β, TNF-𝛼, M-CSF, and stimulating RANKL/OPG signaling pathway to inhibit bone regeneration (Fig. 4) [50].

Effects of fluoride on bone regeneration and underlying mechanism. In BMSCs, F− in the range of 0.24–2.4 μM could regulate the formation of vascularized bone in vivo [50]. The peak osteogenic effect of F− was found at 2.4–24 μM, which may be associated with the activation of the canonical Wnt/rmβ-catenin signaling pathway, and the inhibition of RANKL/OPG signaling pathway [46]. During the early response, F− increased the expression of osteogenic factors and angiogenic factors [50]. In macrophages, a low concentration of F− promoted the polarization of macrophages to the M2 phenotype by stimulating NF-κB signaling pathway [50]. High concentrations of F− over 24 μM started to show cytotoxicity, induced the secretion of inflammation factors, and stimulated RANKL/OPG signaling pathway to inhibit bone regeneration [50].
The content of zinc (Zn) normally ranges between 0.012 and 0.025 wt.% in human bone, which is relatively higher than that in whole fat-free adult tissues and plasma [59]. Zn plays a crucial part in a variety of physiological functions, markedly being a co-factor in hundreds of enzymes involved in bone functions and metabolism [35,60].
It was previously reported that among the tested materials, BHA derived from bovine bone doped with 5% ZnO (Zn-BHA) showed the best mechanical properties [59,61,62]. The microstructure of 5% Zn-BHA comprised well-developed ZnO-like crystals dispersed on the surface and pores of the BHA matrix, which enhanced the microhardness of BHA [59]. It should be emphasized that ZnO did not increase the compressive strength of BHA but the hardness only [59]. Besides, the incorporation of ZnO seemed to narrow the sintering temperature range of BHA up to 1200 °C, when Zn-BHA exhibited the best densification behavior [59]. Once the sintering temperature further increased, Zn-BHA became vulnerable, and decomposition occurred.
Based on such structural optimization, Zn-BHA showed concentration-dependent bioactivity, antibacterial, anti-inflammatory and osteogenic abilities [63,64]. In addition, HA doped with less than 1% of Zn showed effective antibacterial properties, while the maximum inhibitory effect of 2% Zn on Gram-positive and Gram-negative bacteria was (50 ± 5)% and (77 ± 5)%, respectively [65,66]. The cytocompatibility of Zn2+ was often described as a non-toxic level [67]. In some situations, however, the toxicity was observed with Zn-HA and different Zn-based compounds as well, which was probably due to the mechanical damage caused by a sedimentation of Zn-HA particles over cell membranes [68,69]. Indeed, it was reported that Zn-HA which formed the largest particles with irregular shapes and hard edges, would induce the highest lethality due to the mechanical damage to cell membranes [68].
Moreover, it was found that Zn2+ could promote osteoblast proliferation and osteocalcin production, accelerate bone matrix maturation, and inhibit osteoclast activity [62]. The deficiency of Zn2+ in vivo could alter the number and function of neutrophil granulocytes, monocytes, natural killer-, T-, and B-cells [70]. The potential mechanism might be closely related to the effect of Zn2+ on osteoimmunomodulation, as Zn2+ homeostasis was tightly controlled by a transporter family called Zn signaling [70–72]. Besides, Zn2+ could increase the release of anti-inflammatory IL-10, and reduce the expression of TNF-𝛼 and IL-1β, which may be due to the regulation of TLR-4 signaling pathway [41,73,74] (Fig. 5), although the detailed molecular mechanism needs to be clarified further.
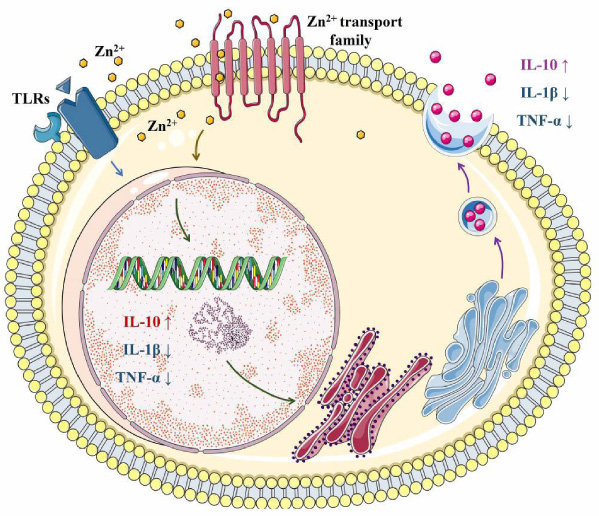
Magnesium (Mg) is the fourth most abundant cation in the human body, and its deficiency can lead to bone loss. Thus, the doping of Mg seems to be a potential approach to improve the biological properties of bone grafting materials [47,75].
When BHA was doped with MgO, SiO2, Al2O3, ZrO2 and Y2O3, respectively, the crystal lattice of BHA had a higher capability to accommodate Mg2+ than other cations attributed to the difference in cation size [78]. Mg-BHA was the only one following the general trend of better sintering performance at higher temperature, and the best one with sintering performance and density [78]. It is worth noting that when sintered at 650–1000 °C, Mg doping might induce a partial decomposition of HA into tricalcium phosphate (TCP), forming Mg-HA/TCP compound [79]. It contained a distinctly different microstructure from single phase Mg-HA, including mixed needle-like and plate-like crystalline particles, with improved specific surface areas over the range of 91.0–104.3 m2/g [79].
It is known that Mg shows a high degree of biocompatibility with living cells, and plays an important role in bone health by stimulating osteoblast proliferation in the early stages of osteogenesis [79]. It has been demonstrated that Mg could enhance BMSCs adhesion and further stimulate osteoblast proliferation [80,81]. Mg was found to promote new bone formation on dental implants, control the initial degradation rate and guide bone regeneration [82]. Mg2+ doped biological hydroxyapatite (Mg-BHA) has a similar composition, morphology, and crystallinity to that of biological apatite, without cytotoxic effects at low doping concentrations of Mg2+. In contrast, a higher concentration of Mg2+ at 5 mM may impair osteoblast activity and contribute to bone diseases [76,77]. A previous study modified BHA thin film with MgF2 or MgO by pulsed laser deposition technique, which could reinforce HA hexagonal-type phase (P63/m (176) space group) and increase the spheroidal crystal particulates over the thin film to enrich the surface morphology [83]. This Mg-BHA thin film could not only promote cell adhesion and further improve the osseointegration properties of implants, but also show good performances in cell proliferation, osteogenic and antibacterial activities [83].
The underlying mechanism of Mg2+ in bone regeneration and immune modulation seems closely related to the stimulation of BMSCs and macrophages both. In BMSCs, it was reported that Mg2+ induced the accumulation of active β-catenin, and further up-regulated the expression of Wnt/β-catenin signaling pathway to stimulate the osteogenic differentiation [84,85].
For macrophages, it was found that Mg2+ suppressed the production of TNF-𝛼, IL-6 and other inflammatory cytokines by inhibiting TLR signaling pathway, which ulteriorly promoted the polarization of macrophages to the M2 phenotype [42,86]. In addition, Mg2+ promoted the expression of IL-10, BMP-2, transforming growth factor β1 (TGF-β1) and other osteogenic related factors [86]. TGF-β, which played a significant role in osteoimmunomodulation, could further inhibit the inflammation progress in bone tissue and enhance the recruitment and mobilization of BMSCs [87,88]. Most recently, it was reported that Mg2+ may act in a previously unknown biphasic mode in bone repair [49]. In the early stage of inflammation, Mg2+ contributed to an upregulated expression of TRPM7 in macrophage, which caused Mg2+ influx and consequently led to the cleavage and nuclear accumulation of TRPM7-cleaved kinase fragments (M7CKs) [49]. Then, M7CKs triggered the phosphorylation of histone 3 at serine 10 (H3S10) in nucleus, stimulated the promoters of inflammatory cytokines and therefore resulted in the primary formation of pro-osteogenic immune microenvironment [49]. In the later remodeling phase, the continuous Mg2+ exposure upregulated NF-κB signaling pathway, which may increase osteoclastic-like cells and inhibit bone regeneration [49] (Fig. 6). Hence, Mg2+ seems to be a crucial regulator worthy for further exploration, as it locates at the central intersection of complex cell signaling pathway network for bone regeneration and immune regulation.
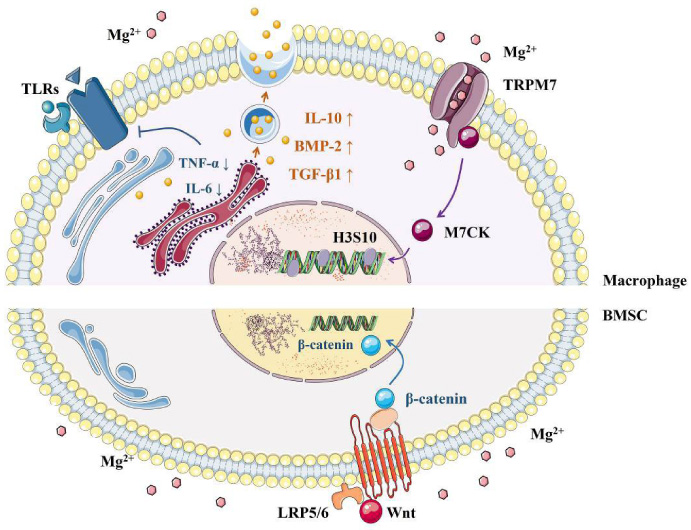
Effects of magnesium on bone regeneration and immune modulation. In BMSCs, Mg2+ induced the accumulation of active β-catenin and further upregulated the expression of Wnt/β-catenin signaling pathway [84,85]. For macrophages, Mg2+ suppressed the production of inflammatory cytokines by inhibiting TLR signaling pathway [42,86]. In addition, Mg2+ promoted the expression of osteogenic related factors [86]. In the early stage of inflammation, Mg2+ upregulated the expression of TRPM7, which led to Mg2+ influx [49]. Then, M7CKs triggered the phosphorylation of H3S10 in the nucleus, which stimulated the promoters of inflammatory cytokines [49]. In the later remodeling phase, continuous Mg2+ exposure upregulated NF-κB signaling pathway, which may increase the proliferation of osteoclastic-like cells and inhibit bone regeneration [49].
Lithium (Li) is present in human body in trace amounts as an essential element [89]. Li+ is smaller than Mg2+ and Zn2+, so that it can easily pass through biological membranes to reach target sites [90], indicating its medical potential as an active ion. It has been proven to be a reliable pharmaceutical ingredient with few side effects [91].
It was reported that doping Li2CO3 could increase the porosity of BHA [91,92]. Besides, Li-substituted BHA thin films with PLD using Li2CO3 and Li3PO4 as dopants showed obvious surfaces with a rough and irregular morphology, consisting in general of spheroidal particles, which improved the angiogenesis capability and osteo-conductivity of BHA [92]. Additionally, Li-BHA films elicited improved wettability due to proper porosity and roughness, which could further induce rapid bone regeneration compared with BHA coatings [92]. This was supported by an in vivo evaluation on the osteogenic performance of Li-BHA coatings, where more new osseous structures were detected. It was believed that the incorporation of Li might create a delivery vehicle for bioactive agents to promote, even accelerate osseointegration in close relation with an improved anchorage of bone metallic implants [5]. However, for Li+, there is still a lack of detailed molecular mechanism, especially in promoting bone regeneration and osteointegration.
Others
The physiochemical and biological features of some other ion-incorporated BHA have also been investigated. Both positive and negative effects have been observed.
BHA incorporated with Na4P2O7 (NP) was synthesized in 1999. With the increase of NP content, BHA gradually transformed into different crystalline phases composed of TCP/HA, TCP/HA/NP or TCP/NP, forming natural porous biomaterials with better bioactivity than pure BHA [93]. Besides, it was reported that BHA doped with CeO2 showed reinforced microhardness and compression strength [62]. Other reports demonstrated that the osteogenic activity of Ce was related to TGF-β and BMP-2/Smad signaling pathways [94,95]. Additionally, the expression level of ERK signaling pathway was also correlated to Ce in a time-dependent manner [96]. Moreover, the incorporation of titanium (Ti) caused the cracking of BHA crystals and enhanced porosity [91]. Recently, BHA thin films doped with Ti6Al4V were synthesized by selective laser melting technology, demonstrating better biomineralization capacity and lower degradability than the control [97]. The incorporation of boron (B) into BHA led to improved compressive strength [61], whereas the doping of SiO2 resulted in a structural degradation of BHA [78].
In contrast, it was reported that Al2O3 doping showed no physiochemical enhancement on BHA, which was probably due to the large gap between the optimal sintering temperature of Al2O3 and BHA [78,98]. The doping of ZrO2 into BHA showed a microstructure similar to Al2O3 doping, indicating poor improvement of the mechanical characteristics of BHA [78].
Conclusions
In the hope of improving the osteogenic property of BHA-based bone grafting materials, a new concept, namely osteoimmunomodulation has drawn much attention and been followed. Among several approaches, the doping of active inorganic ions into BHA has been demonstrated to be a reliable, reproducible and cost-saving strategy. Also, enhanced osteogenesis has been observed for such ion doped BHA in vitro and in vivo. Although some molecular signaling pathways have been confirmed in the process of ion-doping induced osteoimmunomodulation in macrophages, further studies are still needed as multiple signaling pathways seem involved for each single ion simultaneously. Moreover, it is also of great necessity to explore the effects of multiple ionic co-doping into BHA and the potential mechanism. Additionally, it is of more crucial significance to carry out more translational researches of ion-doped BHA, so as to truly promote the development and application of bone grafting materials.
Footnotes
Acknowledgements
This work was supported by the National Natural Science Foundation of China (NSFC 81970975, 81901055).
Conflict of interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this article.