
Abstract
BACKGROUND:
Human and equine patients are known to frequently develop vascular complications, particularly thrombosis both in veins and arteries as well as in the microvasculature.
OBJECTIVE:
The aim of the present study was to investigate and compare the angiogenic response of human and equine endothelial cells to lesions in an in vitro scratch assay.
METHODS:
Endothelial cells from human umbilical vein (HUVEC), abdominal aorta (HAAEC) and dermal microvasculature (HDMEC) as well as equine carotid artery (EACEC) and jugular vein (EVJEC) were cultured and an elongated defect was created (scratch or “wound”). Cultures were monitored over a period of 90 hours in a live cell imaging microscope.
RESULTS:
In the human endothelial cell cultures, there was a uniform and continuous migration of the cells from the scratch fringe into the denuded area, which was closed after 17 (HUVEC), 15 (HAAEC) and 26 (HDMEC) hours. In the equine endothelial cell cultures, a complete closure of the induced defect occurred after 17 (EVJEC) and 35 (EACEC) hours.
CONCLUSIONS:
In the equine arterial cells, the delay in closure of the denuded area seems to be the results of a disoriented and uncoordinated migration of endothelial tip cells resulting in slow re-endothelialization.
Keywords
Introduction
Thrombosis, the formation of a thrombus, is a major contributor to the global disease burden [1]. For different reasons thrombosis is seen less frequently in veterinary mammalian patients than in human patients [2]. However, there is an exception and this is the horse, that similar to humans, often displays severe thrombotic complications. Thromboses in human and equine patients can develop in veins [3, 4] arteries [5, 6] and in the microvasculature [7, 8]. A cell population with a central role in thrombosis is - besides platelets - the vascular endothelium [9, 10]. It covers the entire inner surface of blood vessels. Far from being a uniform “wallpaper” of the vascular lumen, endothelial cells (ECs) are a heterogeneous population of cells. Not only is it that the endothelial cells of the arteries and veins differ, but also in microvascular capillary beds of different organs, and even within one organ there are differences between endothelial cell populations [11, 12].
Thrombosis following endothelial injury due to a mechanical lesion has been described in humans [13] and horses [14, 15]. For example, the use of indwelling central venous catheters in human patients is associated with endothelial denudation, followed by activation of coagulation and thrombosis [16]. Similarly, horses often develop venous thromboses, especially of the jugular vein, after injections and catheterizations [4]. Unlike venous thrombi, arterial thrombi typically form under conditions of high blood flow and are initiated by endothelial erosions. They may lead to myocardial infarction, ischaemic stroke, and peripheral arterial occlusion in humans [17] and to acute thrombosis particularly of the horse’s limb arteries [18]. Thromboses in humans [19] and in the horse frequently occur in the microvasculature. For example, organ thromboses are life-threatening complications in horses with acute gastrointestinal disease associated with laminitis, a painful and potentially crippling condition of the hoofs that can be fatal to horses [7, 20].
In their review, Yau et al. (2015) describe the “endothelial cell control of thrombosis” where healthy endothelial cells express anticoagulant factors that prevent thrombosis. With thrombosis, endothelial cell proliferation and migration, the hallmarks of angiogenesis, seem to be involved in both, thrombogenesis as well as thrombolysis. Upon vascular injury, in a healthy individual, endothelial cells trigger fibrin formation and platelet adhesion to close the defect temporarily until cellular repair of the defect takes place. However, if endothelial cells do not proliferate and migrate effectively to close the defect, thrombogenesis may lead to the formation of a thrombus [10].
Recent investigations revealed that endothelial cells are also intimately engaged with thrombolysis by releasing pro-fibrinolytic agents that initiate fibrinolysis to degrade a thrombus [4]. Alias et al. [21], who investigated thrombolysis in mice, suggested that during the early phase of thrombus resolution endothelial cells migrate into the thrombus in order to resolve it. They examined mice with an endothelial-cell-specific conditional deletion of vascular endothelial growth factor receptor 2/kinase insert domain protein receptor (VEGF-R2/Kdr) and found that the defective angiogenesis of these animals delayed thrombus resolution [21].
So far, the horse has not been considered as a model for human vasculature and thrombosis research even though, in contrast to human tissues, samples for in vitro studies from healthy equine donors are readily available.
As endothelial cell migration seems to be a key factor in both, thrombogenesis as well as thrombolysis, the aim of the present study was to investigate and compare the migration pattern and migration time in human and equine endothelial cells in an in vitro scratch assay (also called wound healing assay). This in vitro assay measures endothelial cell migration after creating an artificial “wound” and mimics cell migration upon wounding in vivo [22]. The basic steps involve creating a “wound” in a cell monolayer via scratching away a proportion of the cells, capturing images at the beginning and at regular intervals during cell migration to close the scratch, and comparing the images to quantify the migration rate of the cells. Inverted phase-contrast microscopy and live cell imaging is used to characterize the mechanisms of angiogenesis involving proliferation, migration and three-dimensional organization of endothelial cells. This assay is well established for human cells [23], however it has never been used on equine endothelial cells. As one of very few research groups, we have isolated and identified equine macrovascular endothelial cells and established an angiogenesis assay in which the stages of the equine angiogenic cascade can be evaluated in vitro [24]. In continuation of this project, the present study presents the establishment of an in vitro scratch assay for equine endothelial cells and the comparison of the outcome of the scratch assay using human endothelial cells and equine endothelial cells.
Material and methods
Human endothelial cells
Human umbilical vein endothelial cells (HUVEC) and human abdominal aortic endothelial cells (HAAEC) were available from our cell and tissue bank and originally a gift, kindly provided by Prof. Robert Auerbach, Department of Zoology, Laboratory of Developmental Biology, University of Wisconsin, Madison, USA. Although of high passage, they displayed good proliferation and migration performance. Human dermal microvascular endothelial cells (HDMEC) were purchased from Lonza Sales AG, Verviers, Belgium.
Equine endothelial cells
Equine endothelial cells, from the jugular vein (EVJEC – one cell isolate from one horse) and the common carotid artery (EACEC – two cell isolates from two horses) were isolated from healthy donors in a previous study [24]. Briefly, to isolate endothelial cells, segments of the blood vessels were filled and incubated with collagenase solution. The cells obtained were seeded onto gelatine coated six-well plates. After cells in each well had formed islets of approximately 25 polygonal cells, these were scraped off and each islet was transferred separately to gelatine coated 24-well plates. They were then available from our cell and tissue bank. To substantiate endothelial identity of the primary equine endothelial cells, they were sorted using magnetically labeled antibodies targeting the human CD31 endothelial marker antigen (CD31 MicroBead Kit, Miltenyi Biotec GmbH, Bergisch Gladbach, Germany) according to the manufacturer's instruction.
Culture conditions
All cultures were grown with the same primary growth medium (Lonza EGM-2MV, see Table 1) at 37°C in a 5% CO2 humidified atmosphere. The medium was renewed every 3-4 days. For expansion, cells were cultured in 6-well plates until they formed a subconfluent monolayer. For the scratch assay, these cells were seeded onto gelatine coated 24-well plates and grown to confluency (for cell passages see Table 2). Then they were used for the scratch assay. The whole experiment was run three times using different parameters for seeding density and time of growth before a scratch was made (Table 2). After scratch closure in cultures of run 1, these cells were also used for run 2. Cells from run 1 (EVJEC, EACEC 1) or 2 (HUVEC, HAAEC, HDMEC, EACEC 2) were frozen and afterwards used for run 3.
Composition of cell culture medium
Composition of cell culture medium
Passages, cell density in monolayer, scratch closure time, seeding densities of cells and time before the scratch was performed in the 3 runs of the trial
P = passage; CD = cell density (cells/cm2) in monolayer at hour 0; SCT = scratch closure time in hours; nc = not confluent.
The migration capabilities of human and equine endothelial cells were evaluated using a scratch assay modified from Liang et al. (2007). For each cell line, three wells of a 24-well plate were used. In the confluent endothelial monolayer of each well and for all cell cultures at one scheduled time point, a “wound” was made by scratching the monolayer with a pipette tip (10 μl, Eppendorf, Cologne, Germany) in a straight line (the morphology of the resulting scratch using different pipette tips and scratching techniques, like a round or straight scratch, was tested in a pilot study in advance). To ensure repeatability all scratches were made by the same technical assistant. Any debris and displaced cells were removed by gently washing the well once with 1 ml of medium. Afterwards, the plate was placed into a temperature-controlled chamber (at 37°C with 5% CO2 and 50% humidity) integrated into an inverted Olympus IX 81 live cell imaging microscope (Olympus, Hamburg, Germany). To document cell migration and compare endothelial migration patterns as well as morphological variation in the samples, digital pictures from the centre of each well were taken at one-hour intervals for up to 90 hours at different magnifications (4x, 10x and 20x objective). A XM10 CCD camera (Olympus, Hamburg, Germany) was used for image-acquisition. The width of each scratch was measured with Olympus Cell R software (Olympus, Hamburg, Germany, V. 3.1). The cell density of the monolayer was calculated at the beginning of the test (hour zero) using Nikon NIS AR software (Nikon, Düsseldorf, Germany). For this purpose, two regions of interest, each 250000 μm2, one left and one right from the scratch, were defined in the pictures acquired with the 4x objective at hour zero. The cells in both regions of interest were counted manually. The time needed for closure of the scratch was defined as the hours until no cell free areas remained in the scratch area.
The statistical analysis of the scratch assay was performed with IBM SPSS 21 (IBM Deutschland GmbH, Ehningen, Germany). The scratch closure time was tested by ANOVA with the factor “isolate”. The level of significance was α= 0.05. Post-hoc-test (Games-Howell) was performed to compare the isolates with each other.
Results
Morphological characterization of the cell lines
HUVEC, HAAEC and HDMEC had similar morphologies in all three runs (Fig. 1 A-C). The cells grew in a cobblestone pattern. Most cells were polygonal or elongated in shape and had a single ovoid nucleus. Many cells of the HUVEC and HAAEC cultures developed long filopodia (100–200 μm) that overlaid those of other cells, within the confluent monolayer. A few cells (predominantly in the HUVEC cultures) were about triple the size of other cells, and had multiple nuclei. Both, HUVEC and HAAEC developed a completely closed contact fringe to adjacent cells.
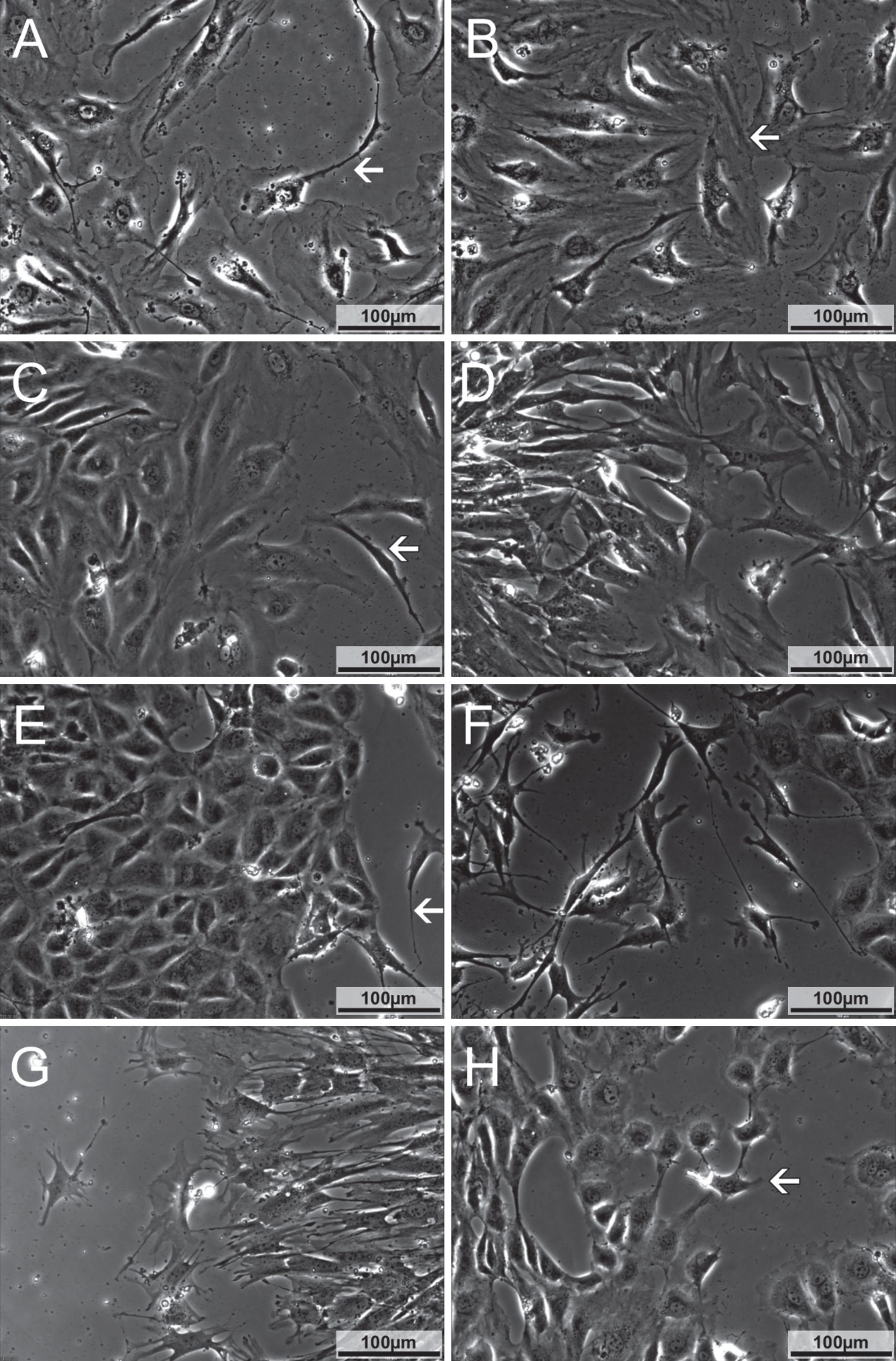
Morphological characterization of cells in the monolayer as well as migrating cells. A) HUVEC at hour 13 in run 3 shortly before closure of the scratch. Cells hold close contact to each other. Cells connected via long filopodia (arrow) can be seen. B) HAAEC at hour 10 in run 3 immediately before closure of the scratch. Cells are in close contact to each other. Leading cells (arrow) have broad contact zones to neighbouring cells. C) HDMEC at hour 34 in run 2. Cells migrate as a sheet, leading cells retain close contact to following cells. On the right side, two elongated leading cells can be seen (arrow). D) EVJEC at hour 23 in run 2. Cells from both sides of the scratch have made contact but still have star shaped morphology and are connected only focally to neighbouring cells. E) EACEC 1 at hour 55 in run 1, growing in a cobblestone pattern. Only few fibroblast like cells can be seen (arrow). F) EACEC 1 at hour 25 in run 1. Numerous fibroblast like cells can be seen, extending long filopodia to neighbouring cells. Only few endothelial like cells are located on the right side. G) EACEC 1 at hour 33 in run 2, with cells having a similar morphology to those in EVJEC, see 1D. H) EACEC 2 at hour 35 in run 2. Cells display a cobblestone pattern and have a completely closed contact fringe to neighbouring cells. Most of the cells are roundish to polygonal in shape. A small fraction of cells, especially the cells at the tips of branches (arrow), shows an elongated shape with multiple filopodia.
Overall, human endothelial cells were larger than equine cells. This is underlined by the cell density counts at hour 0 in the monolayer, in which human endothelial cells were in the range of 26200 and 47733 cells per cm2 and equine cells between 30067 and 114267 cells per cm2 (see Table 2).
EVJEC had a similar morphology in all three runs. The cells were triangular to elongated in shape. Many extended filopodia (100 μm) towards and over neighbouring cells. For most of the cells, a completely closed contact fringe to adjacent cells was not observed until a high cell density was reached. Instead they were connected in small isolated areas to other cells, resulting in star shaped outlines (Fig. 1D).
EACEC (isolate 1) displayed a heterogeneous morphology during the three runs. In run 1, the arterial cells had a cobblestone pattern with round to polygonal cell shapes. In some areas of the culture, their morphology was similar to that described for the EVJEC, being more elongated. Single cells extended long slender filopodia (100–200 μm) towards and over neighbouring cells. These EACEC did not build up a completely closed contact fringe to adjacent cells until a very high cell density was reached (Fig. 1 E-G). In run 2 and 3 their morphology was homogenous and similar to that described for the EVJEC.
The second isolate of EACEC from a different horse was run in parallel to isolate 1. Here the cells grew in a cobblestone pattern, held contact with their neighbours over a broad contact fringe and only a small number of star shaped and elongated cells were present. These were sprout leading cells (“tip cells”) or single migrating cells (Fig. 1H). Filopodia of sprout leading cells were few and short (50 μm).
Different pipette tips were tested and the 10 μl tip was found to be suitable in this assay, creating a scratch fitting with the 10x objective and the field of view of our system. The width of the resulting scratch was approximately 700 μm. The scratch was uniform in human cell lines but more uneven in width and outline in the equine cell lines.
In HUVEC cultures, single cells migrating out of the monolayer edges were followed closely by other cells. This resulted in a constant movement of a sheet of cells with a homogenous migration fringe, oriented towards the opposite side of the scratch. During this movement a few isolated cells were observed migrating within the scratch area. The constant migration towards the middle of the scratch area took place in all three runs of the experiment. The building of a monolayer before the scratch as well as the complete closure of the scratch was achieved in all 9 observed wells with a mean scratch closure time of 17 hours (Figs. 2–4 and Table 2).

Comparison of the scratch closure characteristics in the different cell populations as a time series in run 1. Human cultures show similar morphologies and grow in a cobblestone pattern. Human endothelial cells are larger than equine ones, the equine cells growing in a higher density. The different scratch closure times can be observed. E.g. while at hour 10 HUVEC, HAAEC and EVJEC have closed the gap, HDMEC, EACEC 1 and EACEC 2 have still an open scratch area. EACEC 1 presents a heterogeneous morphology with roundish cells growing in a cobblestone pattern as well as fibroblast like cells while in EACEC 2 the round cells dominate. Sprouts as well as single migrating cells can be seen in those isolates. Bar = 500 μm.

Comparison of the scratch closure characteristics in the different cell populations as a time series in run 2. Like in run 1, human cultures show similar morphologies, growing in a cobblestone pattern. They are larger than equine cells and the equine cells grow in a higher density. The different scratch closure times can be observed. E.g. while at hour 15 HUVEC and HAAEC have closed the gap, HDMEC, EVJEC, EACEC 1 and EACEC 2 have still an open scratch area. EACEC 1 presents a homogeneous morphology with elongated cells like EVJEC while in EACEC 2 the round cells still dominate. Sprouts can now be seen mainly in EACEC 2. Bar = 500 μm.

Comparison of the scratch closure characteristics in the different cell populations as a time series in run 3. Like in run 1 and 2, human cultures show similar morphologies, growing in a cobblestone pattern. They are larger than equine cells. The equine cells grow in a similar density but show a subconfluent monolayer. The different scratch closure times can be observed. E.g. while at hour 15 HUVEC, HAAEC and EVJEC have closed the gap, HDMEC has still subconfluent areas and EACEC 1 a bigger gap. EACEC 2 morphology shows round cells but does not form a monolayer until the end of the observation period. Bar = 500 μm.
HAAEC cultures showed a similar migration pattern as described for HUVEC. The constant migration towards the middle of the scratch area took place in all three runs of the experiment. The building of a monolayer before the scratch as well as the complete closure of the scratch was achieved in all 9 observed wells with a mean scratch closure time of 15 hours (Figs. 2–4 and Table 2).
In HDMEC cultures, the cell layer moved as a sheet to close the scratch; also single migrating cells as well as sprouts directed to the opposite side of the scratch could be observed. The constant migration towards the middle of the scratch area took place in all three runs of the experiment. The building of a monolayer as well as the complete closure of the scratch was achieved in 8 of the observed wells (one well fell dry during the experiment) with a mean scratch closure time of 26 hours (Figs. 2–4 and Table 2).
EVJEC started to migrate in a constant movement towards the scratch area. Frequently, single endothelial cells detached from the edge of the cellular layer and migrated into the denuded area, resulting in an irregular appearance of the scratch edges. A constant migration towards the middle of the scratch area took place in all three runs of the experiment. The building of a monolayer as well as the complete closure of the scratch was achieved in all 9 observed wells with a mean scratch closure time of 17 hours (Figs. 2–4 and Table 2).
EACEC (isolate 1) had a heterogeneous migration pattern with focal sprouting of single cells trailed by other cells, moving out of the cell monolayer. Gradually multicellular sprouts formed and migrated/moved randomly through the scratch. There were areas of the scratch line, in which almost no sprouting occurred until very late or not at all (Fig. 5). In addition, single cells, especially cells derived from the tip of the sprouts detached from the cell complex and migrated into the denuded area. They remained there migrating without contact with other cells or after a few hours returned back to the cellular layer. Upon physical contact with cells at the scratch edge they again migrated back into the denuded area for a period until they again returned to the edge of the scratch. This movement pattern was repeated for up to 20 hours until these highly motile cells were integrated in the monolayer covering the scratch area. Sprouting was commonly observed in run 1 in areas with mainly roundish cells growing in a cobblestone pattern (Fig. 1E). In runs 2 and 3, in which cells having elongated to star shaped morphologies predominated, the cells had much less sprouting (Fig. 1G). Instead, they had a similar migration pattern to that described for the EVJEC, although they migrated more randomly and more slowly.
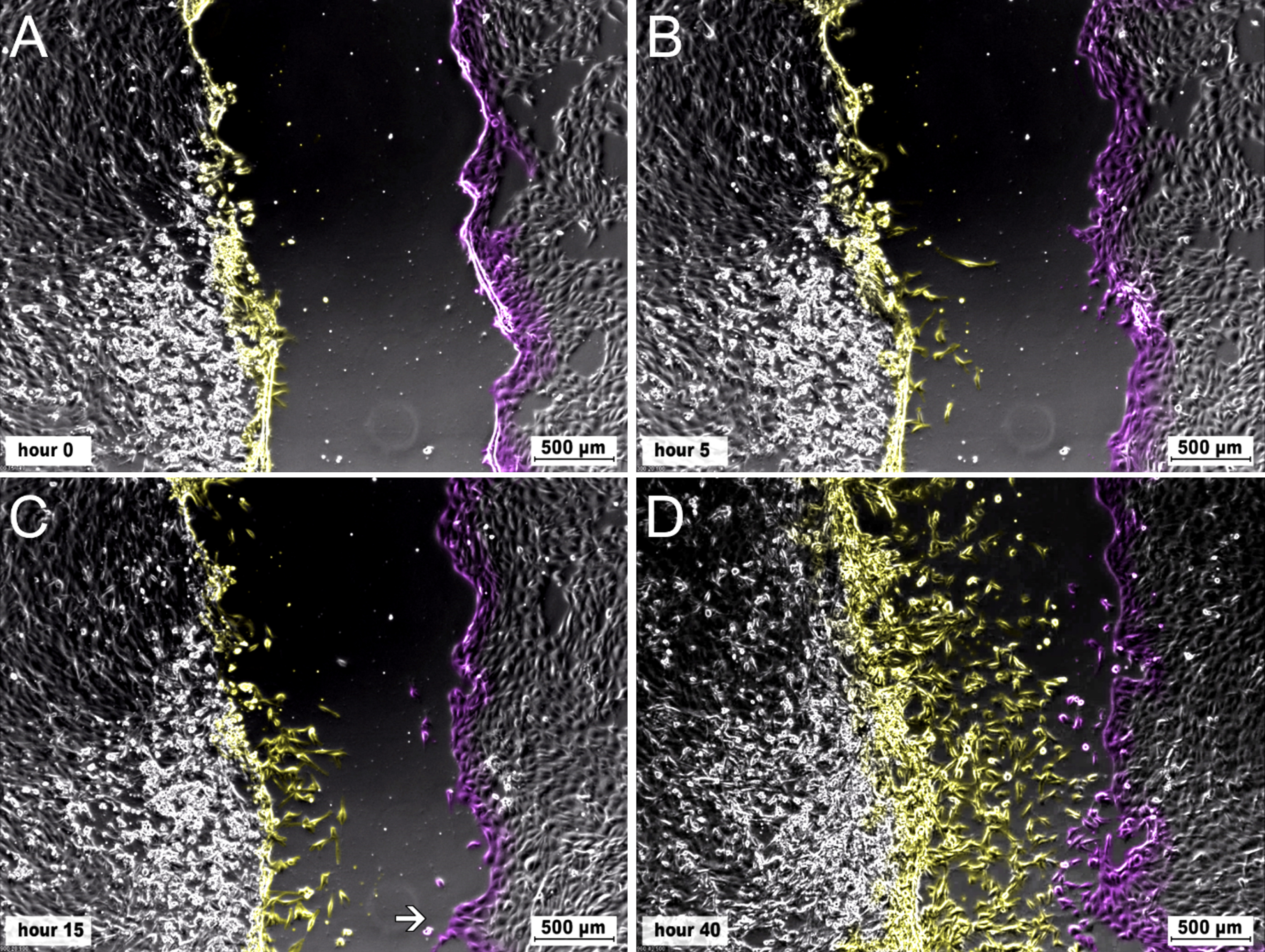
Time series of EACEC 1 from run 1. The opposing scratch borders and cells migrating from there are marked in different colours. A) The cells show endothelium like morphology with roundish cells, growing in a cobblestone pattern. The cells on the left side of the scratch are confluent, on the lower left side cells are very confluent with cells rounding up and detaching. On the right side of the scratch, the cells are partly subconfluent. B) A few cells from the left side start to migrate into the denuded area. They migrate as single cells and show an elongated morphology with multiple filopodia. C) Migration of single cells can be observed over the whole left side of the scratch. Only some single cells migrate from the right side of the scratch into the denuded area. On the lower right side a sprout has formed (arrow). D) While high migration activity of single cells into the denuded area can be seen from the left side of the scratch, most of the cells from the right edge of the scratch keep their position. The sprout (see 5C) did not grow substantially in length.
As a result, migration of the cells towards the middle of the scratch was discontinuous and heterogeneous over the three runs of the experiment. For this isolate, the building of a monolayer was achieved in 7 of the observed wells and the complete closure of the scratch was observed in 6 of the wells. The wells, which showed the heterogeneous scratch closure and also the prolonged time to form a monolayer, were the ones of run 1. The induced lesion in the EACEC monolayer was closed after 35 hours (Figs. 2–4 and Table 2).
EACEC (isolate 2) had a similar migration pattern as described for isolate 1 in run 1. The building of a complete monolayer was achieved in 1 of the observed wells at the scheduled time of scratching and the complete closure of the scratch was observed in 3 of the wells (the rest of the wells became confluent after the scratch was made). The ability to form a monolayer and to close the scratch decreased rapidly from run 1 (passage 5) to 2 (passage 7) to 3 (passage 9). In run 3, the cells of isolate 2 built net-like structures right from the start instead of a monolayer (Figs. 2–4). Since this isolate did not readily form monolayers and the scratch closure time exceeded the observation window, it was excluded from statistical evaluation.
Statistical evaluation showed that the arterial equine isolate (EACEC 1) had a significantly longer scratch closure time than the human isolates HUVEC, HAAEC as well as the venous equine isolate (EVJEC) with a mean difference of 18 hours. Scratch closure of microvascular cells (HDMEC) with an intermediate scratch closure time took significantly longer than HAAEC (Fig. 6).
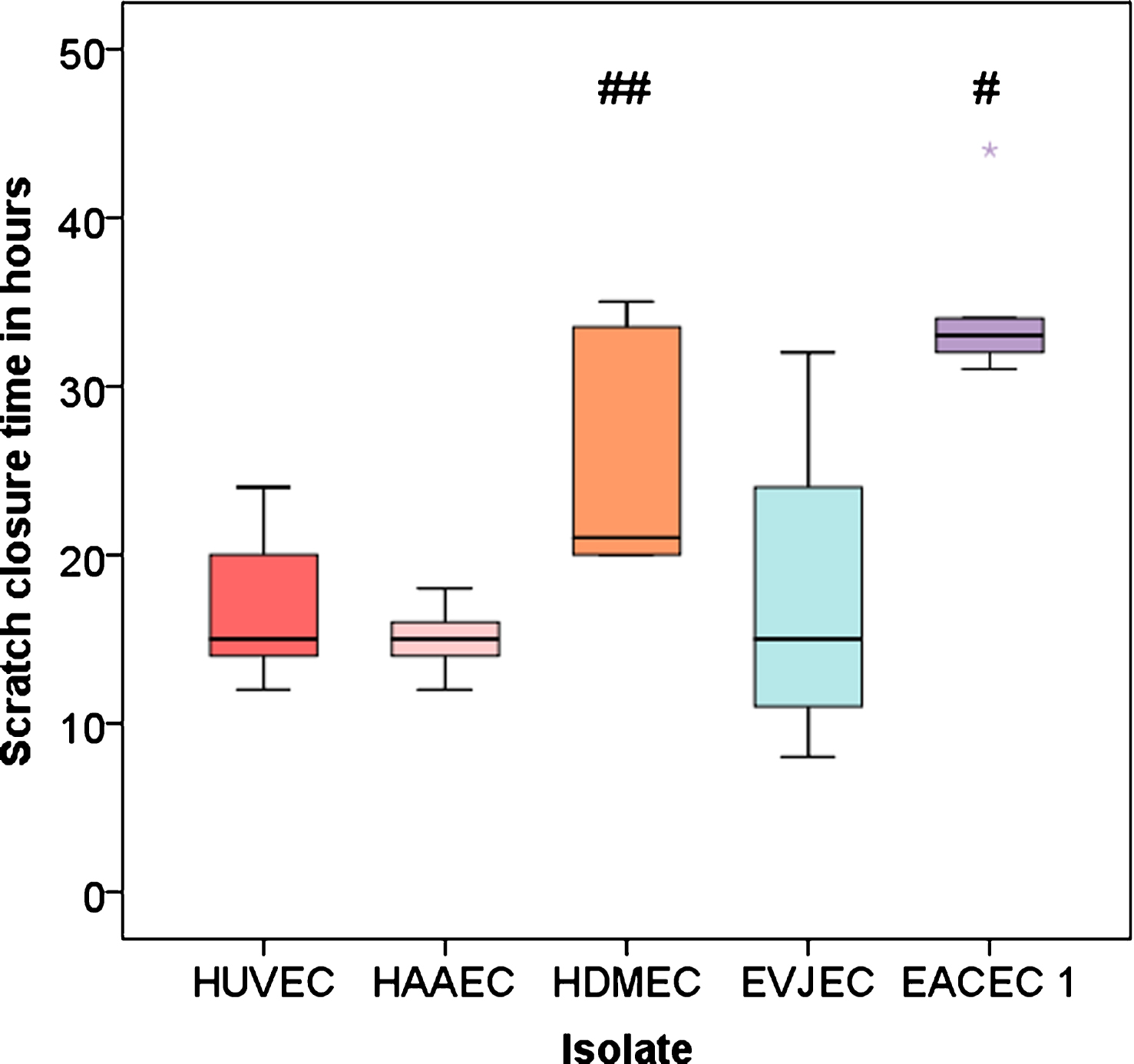
Boxplots for overall scratch closure time. # = P < 0.05 vs HUVEC, HAAEC and EVJEC; # = P < 0.05 vs HAAEC.
The objective of this study was to investigate and compare migration patterns and migration times of human and equine endothelial cells in response to scratch lesions in an in vitro scratch assay. After creating a defect by scratching the endothelial monolayer, cultures were monitored in a live cell imaging microscope until closure of the defect.
Experimental setting
As the required number of cells to build a confluent monolayer depends on both the particular cell type and the size of plates [22], these factors have to be adjusted appropriately. Different seeding densities were tested in the live cell imaging runs. For scratch assays usually confluent endothelial monolayers are used, whereas migration and proliferation assays mostly use subconfluent cultures [25–27]. In our study, the human cell lines showed a homogenous ability to form confluent monolayers with a stable cell density over all tested seeding densities. In the equine isolates, the highest seeding density (50000 cells/well) resulted in highly confluent cell growth, before scheduled scratching. This was reflected by a high cell density in the monolayer at hour 0 and may have had an unfavourable effect due to it being a mature and therefore more inactive cell population [28]. The lowest seeding density (20000 cells/well) as well as the intermediate seeding density (40000 cells/well) resulted in a subconfluent to confluent monolayer at the time of scratching and was thus suitable for the assay. Although a higher seeding density was used in run 3 than in run 2, the cell density was generally lower in run 3. A possible explanation would be the effect of cold storage [29]. Trauma to the jugular vein from catheterization and repeated intravenous injections by drug users are the leading causes of thrombosis in humans [30]. Similarly in horses, especially the left jugular vein is often traumatized and damaged due to frequent intravenous injections [31]. It has been found that repeated endothelial lesions enhance the thrombogenic process [32]. In order to mimic frequent endothelial lesions, cells were transferred from one run to the other and scratch assays were performed, even if the endothelial cultures were subconfluent at the scheduled time of scratching. A 10 μl pipette tip was chosen to make an optimal scratch width of 700 μm that was the best fit for the field of view of our system. Another parameter determined empirically for the particular cell type used was the time frame for incubation [22]. In our experiment, depending on the cell isolate and seeding density, the observation time frame had to be at least 50 hours.
Scratch closure time
Defect closure is dependent on endothelial proliferation and migration, both hallmarks of the angiogenic response and regulated by a plethora of angiogenic stimulators and inhibitors. Normally, the turnover of endothelial cells is low but under certain conditions including mechanical stress or hypoxia, endothelial cells start to proliferate and migrate [33]. In a scratch assay, the behaviour of migrating cells is characterized by a) migration speed, b) migration direction, and c) migration persistence, i.e., the ability of a cell to maintain its migration direction [34].
Migration speed
In this study, the time until scratch closure was similar in human arterial and venous and in equine venous endothelia, i.e., 15–17 hours. In contrast, the equine arterial cells needed 35 h until the scratch was closed, i.e., defect closure took more than double the time. The human microvascular endothelial cells had an intermediate scratch closure time of 26 hours. This shows that in comparison to the other endothelial cells investigated, migration capacity of EACEC was slower. Although the human EC had an even lower cell density at the level of confluence, their time needed to close the defect was shorter but within the range described in current literature [35–37]. It should be noted though, that e.g. HUVEC after passage 15–20 are considered as senescent by some authors [29, 38]. This might explain the heterogeneous morphology with smaller and bigger cells and low cell density found in this study. In senescent HUVEC, bigger cells compensate for reduced migration speed [35]. Proliferation of endothelial cells already changes in much earlier passages. Krüger et al. examined the influence of the number of cell passages on the endothelialisation of a polystyrene surface. They emphasized that only by using HUVEC of the first passage, cell densities occurred, which were comparable to cell densities in vivo [39].
Migration direction
Contrary to the other endothelial types investigated, in the EACEC multicellular sprouts developed at the defect line that were not oriented towards the centre of the scratch but rather grew more randomly. The undirected migration of sprouts in the EACEC could be interpreted as a deficiency in the tip cells. These cells are critical in the first step of angiogenesis that occurs with the formation of a new sprout. The tips of these sprouts are formed by specialized endothelial cells called tip cells. Tip cells are migratory and extend numerous filopodia to sense their microenvironment and determine the direction of the new sprout’s growth. Following behind the tip cells, other endothelial cells become so called stalk cells that proliferate and form the trunk of the new blood vessel [40, 41]. It appears plausible that dysfunction of endothelial tip cells may lead to disturbed angiogenesis. Recently it was found that directed migration seems to be under the control of ephrin-A1, which is involved in migratory direction and speed. Silencing of ephrin-A1 leads to a faster but more disorientated migration in wound-healing assays [42]. Because of their essential role, tip cells are a potential therapeutic target for anti-angiogenic therapies [40, 41] and e.g. the role of ephrin-A1 should be investigated in future studies.
Migration persistence
In addition to their random growth pattern in sprouts at the defect line, single EACEC displayed poor migration persistence as shown by uncoordinated migration between the denuded area and the cellular fringe of the lesion for up to 20 hours. A potential explanation for this may be the absence of adequate vascular endothelial growth factor (VEGF) guidance for these cells. Tip cells are guided by gradients of matrix-bound VEGF. They explore their environment by extending slender motile filopodia, which express VEGF receptor (R)-2 and -3 and transduce the migratory signal initiated by VEGF binding [43]. The medium used for EACEC was the same as used for the other cultures, containing the same amount of VEGF. However, VEGF concentration levels for different endothelial populations differ. Noren et al. [44] found that endothelial cells decode VEGF-mediated Ca2 + signalling patterns to produce distinct functional responses. They used different VEGF concentrations and triggered distinct and mutually exclusive cellular behaviours and accurately predicted cell responses depending on VEGF gradients [44]. So far little is known about demands for VEGF of different endothelial populations including equine endothelial cells. Additionally, growth factors seem to be a two edged sword in wound healing, since the application of most growth factors has the potential of scattering cells, thus compromising barrier function [45].
In summary, the delay in closure of the denuded area in the EACEC seems to be a combination of all three parameters. Not only was migration speed lower in EACEC but also their migration direction and persistence resulted in slower re-endothelialization.
Morphological features of migrating cells - Collective migration and endothelial to mesenchymal transition?
EVJEC as well as the later runs of the EACEC 1 had a different morphology to that of human endothelial cells as well as EACEC 2, being more spindle shaped and resembling fibroblasts. EACEC 1 was a morphologically heterogeneous cell population. In the beginning of run 1 the typical endothelium like phenotype, with roundish cells growing in a cobblestone pattern, dominated. During the experiment, the spindle shaped cells took the lead. In run 2, in which the cells from run 1 were seeded anew, they were the prevalent cell population. A general pattern was noted: cells that grew in a cobblestone configuration migrated either collectively by forming sprouts or as a sheet with leading cells maintaining close contact with those following. Fibroblast like cells migrated predominantly as single cells. Collective migration is one of the hallmarks of successful wound healing of many types of epithelia and the endothelium, during which cells migrate collectively as a coherent sheet, maintaining intercellular connection and relative positions. To maintain proper cell-cell adhesion is especially important during wound healing so that the barrier is not compromised [45]. Therefore, another prominent difference found in our study, the heterogeneous contact zone to neighbouring cells, seems to be of importance. While the human cells and EACEC 2 had contact to their neighbours all over their cell membrane, EVJEC and EACEC 1 (spindle shaped type) had only focal attachments to neighbouring cells until a high cell density was reached, again reminiscent of fibroblasts. A possible explanation for this fibroblast-like appearance of endothelial cells would be endothelial-to-mesenchymal transition. This phenomenon is connected to an alteration in the endothelial morphology, i.e., endothelial cells acquire mesenchymal characteristics such as an elongated, fibroblastoid morphology, increased motility and cell-to-cell junction rearrangement [46]. In this context it is remarkable that endothelial to mesenchymal transition is found predominantly in the arterial tree. It is characterized by a build-up of intraluminal plaques that can lead to thrombosis both in humans [47] and horses [18]. Markers of endothelial-to-mesenchymal transition have been found in thrombotic microangiopathy [48].
It is concluded that collective migration and endothelial-to-mesenchymal transition would be imminent future study targets. So far, a morphological description as well as the general migration pattern of the cells can be given. The collective migration of endothelial cells is modularly controlled to regulate cell motility, directed migration, cell-cell coordination, and cell density [49]. These parameters should be examined in more detail.
Footnotes
Acknowledgments
The authors thank Sophie Richter, graphics designer at the Institute for Veterinary Anatomy for support with pictures.