
Abstract
Lipid-containing adipocytes can dedifferentiate into fibroblast-like cells under appropriate culture conditions, which are known as dedifferentiated fat (DFAT) cells. However, the relative low dedifferentiation efficiency with the established protocols limit their widespread applications. In this study, we found that adipocyte dedifferentiation could be promoted via periodic exposure to cold (10°C) in vitro. The lipid droplets in mature adipocytes were reduced by culturing the cells in periodic cooling/heating cycles (10–37°C) for one week. The periodic temperature change led to the down-regulation of the adipogenic genes (FABP4, Leptin) and up-regulation of the mitochondrial uncoupling related genes (UCP1, PGC-1α, and PRDM16). In addition, the enhanced expression of the cell proliferation marker Ki67 was observed in the dedifferentiated fibroblast-like cells after periodic exposure to cold, as compared to the cells cultured in 37°C. Our in vitro model provides a simple and effective approach to promote lipolysis and can be used to improve the dedifferentiation efficiency of adipocytes towards multipotent DFAT cells.
Introduction
Dedifferentiated fat (DFAT) cells have become a promising cell source in stem cell therapy. They could be obtained via the process of dedifferentiation of mature adipocytes. DFAT cells express embryonic stem cell markers [1, 2] and show the multilineage differentiation potential [3–7]. The osteogenic and adipogenic potential of human DFAT cells was even higher than that of adipose tissue-derived MSCs [8] and similar to bone marrow-derived MSCs [9]. Notably, DFAT cells showed absence of cell senescence and low risk of tumor formation in vivo [10]. Given the abundance of adipose tissue in human body, DFAT cells may represent a great potential in clinical applications. However, there is currently no highly efficient method to promote lipid loss and adipocyte dedifferentiation in vitro, which limites the application of DFAT cells.
Adipocytes in fat tissue can regulate energy balance by storing and hydrolyzing lipids. The loss of lipids in mature adipocytes in vivo could lead to their transformation into beige adipocytes or dedifferentiation into fibroblast-like cells [11, 12]. It has been also demonstrated that dedifferentiation of mature adipocytes in vitro was accompanied by the loss of lipid [13]. Cumulative in vivo evidences have suggested that periodic exposure to cold could induce lipid loss and white adipocytes transformation into brown adipocytes [14, 15]. The decrease of white adipocytes and increase of brown or beige adipocytes have been observed in mice after exposure to a cold environment [16]. In addition, in vivo studies have shown that the occurrence of brown adipocytes in white adipose tissue during cold exposure was associated with mitochondrial activity to promote lipolysis [17, 18]. In vitro study has also demonstrated that mitochondrial uncoupling could promote lipid loss in adipocytes, and consequently induce adipocyte dedifferentiation [19]. This process involved the regulation of peroxisome proliferator-activated receptor gamma coactivator 1-alpha (PGC-1 α) and uncoupling protein 1 (UCP1) [20]. However, the effect of cold exposure on lipolysis and adipocyte dedifferentiation in vitro as well as the underlying mechanism are still unclear, which hided the large-scale production of DFAT cells.
In this study, we hypothesized that the periodic exposure to cold in vitro might be an effective approach to promote the lipolysis and dedifferentiation of the adipocytes. We investigated the lipolysis in mature adipocytes and the proliferation of the dedifferentiated fibroblast-like cells after periodic exposure to cold. The mechanism of the promoted lipolysis and dedifferentiation after exposure to cold was studied.
Materials and methods
Preadipocytes cultivation and adipogenic induction
The human adipose derived stromal cells were isolated from patient’s adipose tissue as described previously [21]. For cells maintenance, the isolated cells were cultured with Dulbecco’s Modified Eagle Medium (DMEM, Life Technologies, Germany) containing 10% (v/v) fetal bovine serum (FBS, Sigma-Aldrich, USA) and 100 U/ml penicillin-streptomycin (Merck Millipore, Germany), and incubated at 37°C containing 5% (v/v) CO2. The medium was changed every 2 days. For adipogenesis induction, the cells were cultured with adipogenesis differentiation medium (StemPro® Adipogenesis Differentiation Kit, Life Technologies, Germany) for 2 weeks and the medium was changed every 3 days.
Adipocytes exposed to cold
To test the influence of periodic cooling on adipocyte dedifferentiation, the adipocytes obtained from adipose derived stem cell (ADSC) adipogenesis were cultured in a thermal chamber (Instec, USA) providing periodic cooling/heating for 7 days. The temperature in the chamber was cyclic changed from 37°C to 10°C, and the changing frequence was 1 time per hour with lasting for 30 minutes at 10°C and 30 minutes at 37°C. Cells cultured at 37°C were used as control group.
Oil Red O staining
Samples were washed with PBS and fixed in 4% (w/v) paraformaldehyde (Sigma-Aldrich, USA). After washing with PBS and water, the cells were stained with 0.1% (w/v) Oil Red O (Sigma-Aldrich, USA) in 60% (v/v) isopropanol for 15 minutes. The staining solution was removed and the samples were washed with water. The stained lipids in adipocytes were extracted using isopropanol and quantified by measuring the absorbance at 490 nm with a microplate reader (Infinite200 Pro, Tecan Group Ltd., Switzerland).
Free fatty acid assay
The supernatant of the cell cultures was collected. The free fatty acid released into the conditioned medium by lipolysis was quantified using a free fatty acid assay kit (Abcam, UK). In brief, 50μl of standard palmitic acid and 50μl of supernatant were added in 96 well plate. Then, Acyl-CoA synthetase was added and incubated at 37°C for 30 minutes. Finally, the enzyme mix was added and incubated at 37°C for 30 minutes. The optical density at 570 nm was measured using the microplate reader (Infinite200 Pro, Tecan Group Ltd., Switzerland).
Immunocytochemistry
The samples were washed with PBS and fixed with 4% (v/v) paraformaldehyde (Sigma-Aldrich, USA), permeabilized with 0.25% (v/v) Triton X-100 (Sigma-Aldrich, USA), and blocked using 5% (v/v) normal goat serum (Life Technologies, Germany). Samples were incubated with the primary antibodies overnight at 4°C and then incubated with the secondary antibodies (Life Technologies, Germany) for 1 hour at room temperature. The nuclei were stained with 4,6-diamidino-2-phenylindole (DAPI; Sigma-Aldrich, USA). The images were taken using confocal microscopy (LSM780, Carl Zeiss, Germany). The following primary antibodies were used: Ki67 (rat, Cell Signaling Technology, Germany), FABP4 (rabbit, Life Technologies, Germany), Perilipin-1 (goat, Life Technologies, Germany).
mRNA expression analysis
The mRNA level was quantified by qRT-PCR. The total RNA was isolated using the PureLink RNA Mini Kit (Life Technologies, Germany) according to the manufacturer’s protocol. The concentration of the total RNA was measured using the microplate reader (Infinite200 Pro, Tecan Group Ltd., Switzerland) with a NanoQuant Plate (Tecan Group Ltd., Switzerland). The cDNA was synthesized using the RT2 First Strand kit (Qiagen, Germany). Real-time PCR was performed using the StepOnePlus real-time PCR System (Applied Biosystems, USA), and RT2 SYSB Green ROX qPCR Mastermix (Qiagen, Germany) was used in the amplification. The GAPDH was used as a housekeeping gene. The primers in the following were used for each gene.
Statistics
Statistical analysis and plotting were performed using GraphPad Prism (Version 7.0) software. The results are presented as mean±standard deviation (SD). More than three independent experiments were used in the study. Statistical analysis was performed using independent-samples t-test (*p<0.05, **p < 0.01.)
Results
Periodic exposure to cold decreased the lipid droplets in adipocytes
To examine the relationship between periodic exposure to cold and lipid loss in vitro, we evaluated the lipid droplet formation in adipocytes. Bright field images indicated that the volume of the lipid in adipocytes was significantly decreased after periodic exposure to cold (Fig. 1A). Oil Red O (ORO) staining further showed that large lipid droplets were replaced by small lipid droplets, and more cells lost their lipids completely and turned into fibroblast-like cells after periodic exposure to cold, compared to adipocytes cultured in 37 °C (Fig. 1B). Compared to cells maintained at 37°C, the extracted ORO reduced approximately 59% in adipocytes after periodic exposure to cold (Fig. 1C). The results suggested that the periodic exposure to cold promotes lipid loss in adipocytes in vitro.
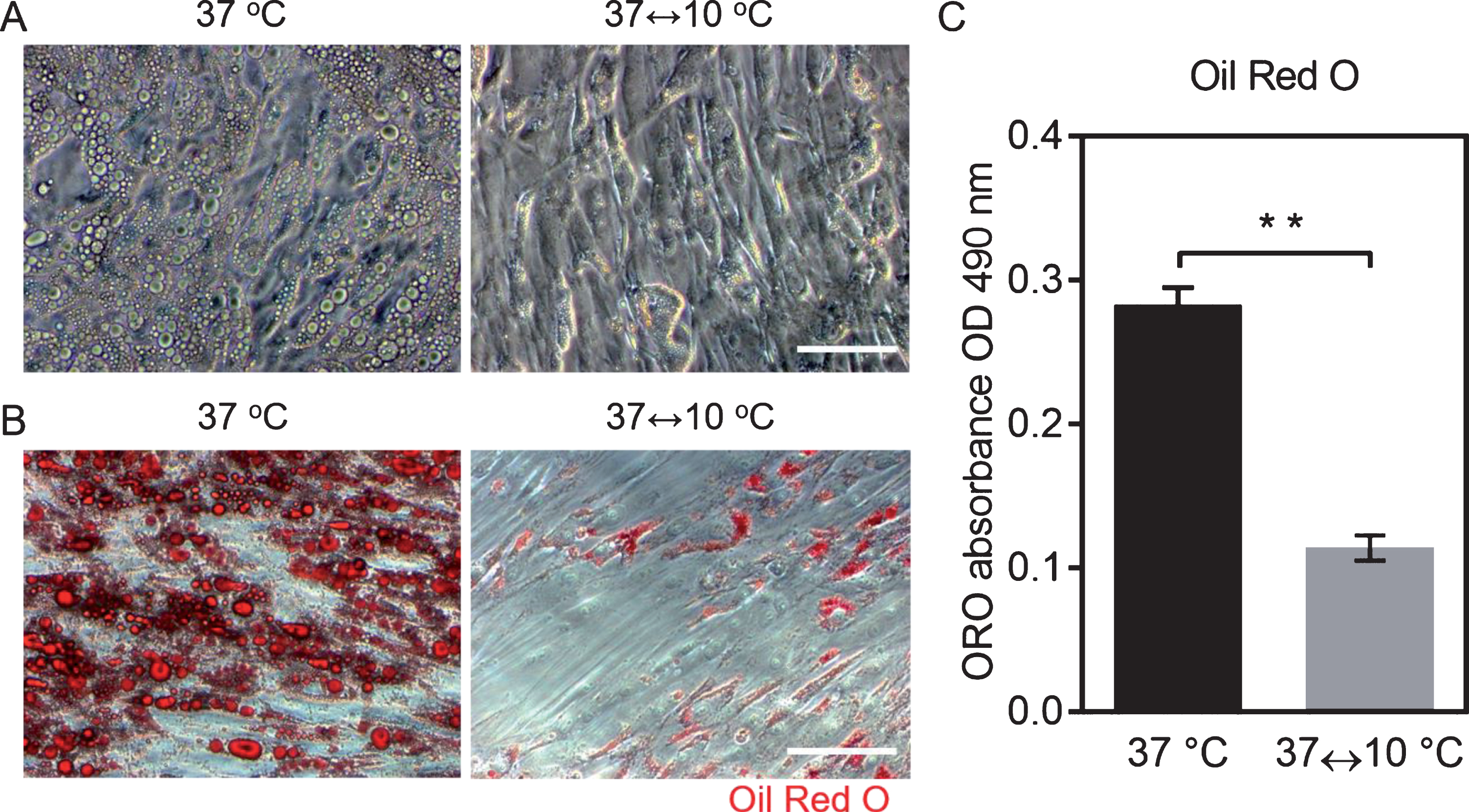
Decreased lipids in adipocytes after periodic exposure to cold. A. Bright field microscopy images of cells on TCP after periodic exposure to cold and to 37°C for one week, scale bar = 100μm. B. Oil Red O staining for lipids in adipocytes after periodic exposure to cold and to 37°C for one week, scale bar = 100μm. C. Quantification of Oil Red O (n = 6; mean±SD; p < 0.01).
In order to understand the lipid metabolism after periodic exposure to cold, we determined the amount of the lipid storage protein perilipin-1 and hydrolysis of triglycerides. After periodic exposure to cold, the perilipin-1 expression was decreased in adipocytes, and the lipid droplets were smaller compared with the adipocytes maintained at 37°C (Fig. 2A). Lipolysis induces the lipid droplets hydrolysis into glycerol and free fatty acids, which is important for adipocyte dedifferentiation. Therefore, the amount of free fatty acids in the culture medium was evaluated. The result showed that the amount of free fatty acids in the medium was almost doubled after the periodic exposure to cold (37↔10°C: 78.94±16.26μM), as compared to the control group (37°C: 38.12±5.21μM) (Fig. 2B). These results suggested that the periodic exposure to cold promoted the lipolysis, which led to a decrease of lipids in adipocytes.

Periodic exposure to cold improved lipolysis in cells. A. Perilipin-1 immunostaining after periodic exposure to cold and to 37°C for one week, scale bar = 100μm. B. Quantification of free fatty acid in culture medium after one week periodic exposure to cold and to 37°C (n = 7; mean±SD; p < 0.01).
To confirm adipocyte dedifferentiation, the mRNA expression level of mature white adipocyte related genes (fatty acid-binding protein 4 (FABP4) and Leptin) were determined. It was found that FABP4 and Leptin were down-regulated significantly in the adipocytes with periodic exposure to cold for 1 week, compared to the adipocytes maintained at 37°C (Fig. 3). The mRNA expression levels of the transcription factors PPARγ and C/EBPα did not showed significant differences between these two groups (Fig. 3).

Expression of adipogenic genes after one week of culture in 37 °C and periodic exposure to cold (n = 6; mean±SD; **p<0.01).
In order to explore the relationship between increased lipolysis, decreased mature adipocytes population, and periodic exposure to cold, we examined the mitochondrial activity, which plays a central role in heat production in order to overcome periodic exposure to cold. The mRNA expression of UCP1 and PGC-1α were up-regulated significantly after periodic exposure to cold for one week, compared with adipocytes maintained at 37°C. In addition, the treatment of periodic exposure to cold enhanced the mRNA expression of PR domain containing 16 (PRDM16) (Fig. 4). The high expression of PRDM16 accelerated the lipids loss and promoted the mature white adipocyte transformation into brown adipocyte. The up-regulation of UCP1, PGC-1α and PRDM16 after periodic exposure to cold could enhance the lipolysis in adipocyte via mitochondrial uncoupling, which further improved the dedifferentiation process.

Expression of mitochondrial uncoupling and brown adipocyte transformation genes was up-regulated after periodic exposure to cold for one week (n = 6; mean ± SD; *p<0.05, **p<0.01).
To evaluate the proliferation activity of dedifferentiated cells from mature adipocytes, the expression of proliferation marker Ki67 and adipocyte marker FABP4 were studied. The immunostaining images displayed more Ki67 positive cells after periodic exposure to cold, compared to cells maintained at 37°C (Fig. 5). The Ki67 high-expressing cells showed the low expression of FABP4. These results suggested that the mature adipocytes dedifferentiated with a higher proliferation potential after exposure to cold.
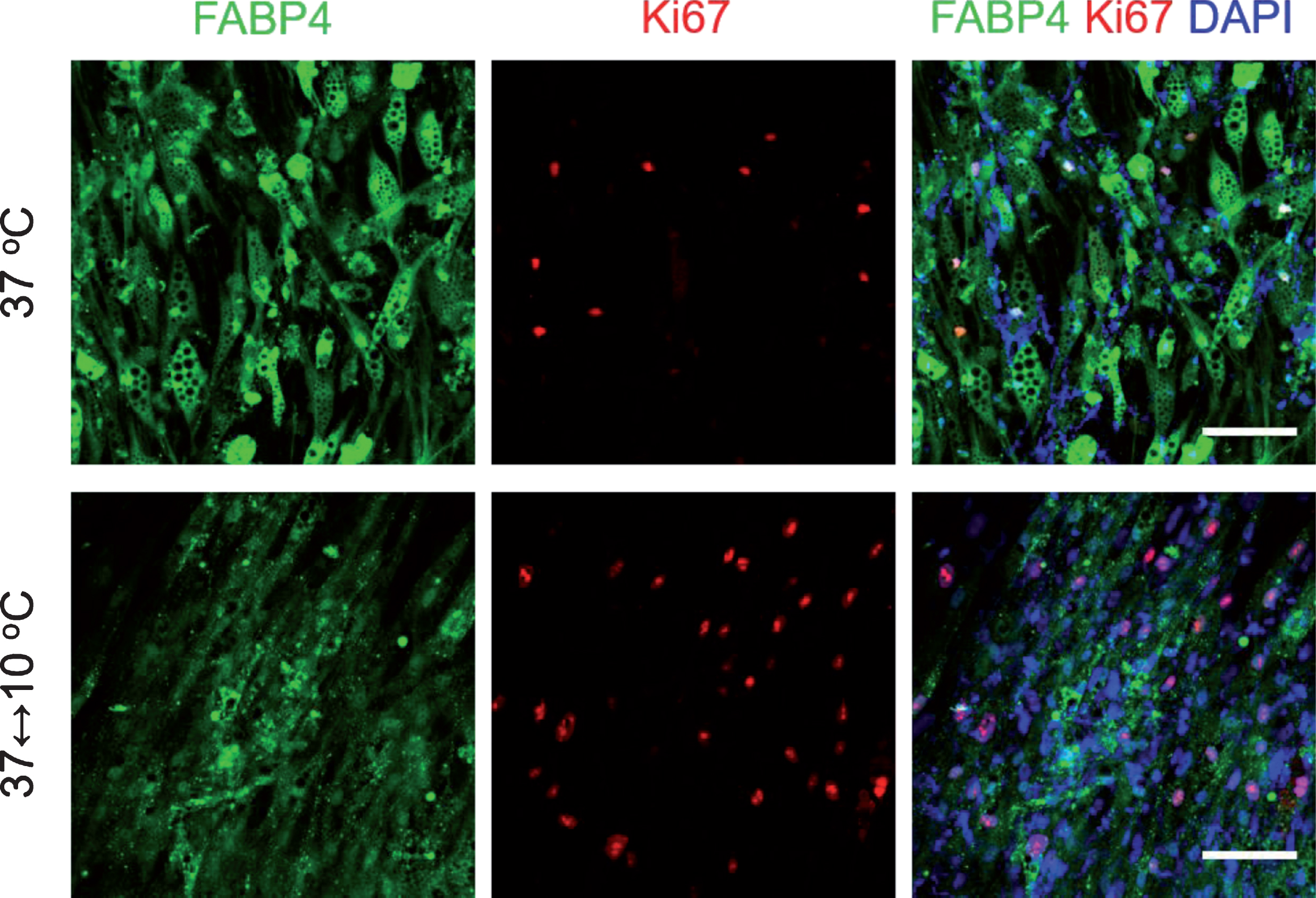
Periodic exposure to cold for one week decreased expression of mature adipocyte marker and increased cell proliferation activity. Cells were immunostained to visualize adipocyte marker FABP4 (green), proliferation marker Ki67 (red) and DAPI (blue). Scale bar = 100μm.
Adipocytes can dedifferentiate both in vitro and in vivo, while the underlying mechanism is still unclear. In vivo studies suggested that exposure to cold could enhance the lipid loss and brown adipogenesis through the mediation of β-adrenergic receptor [14, 22]. It is found that cold, rather than sympathomimetics, is the major factor to activate mature white adipocyte transformation into brown adipocyte in vivo. Nevertheless, the in vivo system relying on animals is different from humans, and it is hard to clarify the molecular mechanism at single cell level. In this study, we investigated the periodic cooling effects on adipocyte dedifferentiation in vitro. The results provided an evidence that the periodic exposure to cold could induce lipolysis in adipocytes and promote mature adipocyte dedifferentiation. The cell model in vitro used here excluded the influences of the neuroendocrine system.
Mitochondria uncoupling plays a key role in the process of lipid metabolism and adipocyte dedifferentiation [19]. UCP1 and PGC-1α are genes related to lipid consumption by mitochondrial uncoupling [24, 25]. We found that the periodic exposure to cold up-regulated the UCP1 and PGC-1α mRNA levels and improved the loss of lipids, which was consistent with the previous work [26]. FABP4 and Leptin are significantly expressed in mature white adipocytes [27, 28]. The down-regulation of FABP4 and Leptin and the up-regulation of PRDM16 suggested the transformation of mature adipocytes into brown adipocytes. Our results confirmed that the periodic cooling could enhance the mitochondrial activity, which was highly related to the dedifferentiation process. The transcriptional factors PPARγ and C/EBPα are involved in adipogenesis as well as the process of transformation or dedifferentiation of white adipocyte. The PPARγ could regulate the white and brown adipogenesis, the transformation from white to brown adipocyte, and lipolysis [29–31]. C/EBPα played a central role in white adipose development, but its function in brown adipocyte transformation is still controversial [32–35]. In our study, the mRNA levels of PPARγ and C/EBPα did not show significant differences between the dedifferentiated and non-dedifferentiated groups, suggesting that their expression levels were not closely associated with the dedifferentiation process.
Accumulation and consumption of lipids are important for adipocyte differentiation and dedifferentiation. The lipids in mature adipocyte become unstable in the dedifferentiation process. The surface of the lipid droplets was coated with perilipin-1 protein in adipocyte, which is important to regulate the lipid accumulation [36]. In this study, the perilipin-1 expression was decreased with the increase of lipolysis after periodic exposure to cold. Previous studies using perilipin-null mice [37, 38] and gene transfection technique [39] have shown that high expression of perilipin can inhibit lipolysis during white adipocyte differentiation, which was consistent with our results. However, it is still unclear how the periodic cooling could affect the activity of perilipin. Previous studies proposed that phosphorylation of perilipin could induce lipolysis via PKA pathway [40, 41]. Therefore, further study focusing on the phosphorylation of perilipin in periodic cooling condition might be helpful to address this question.
In this study, the adipocytes differentiated from ADSCs were used as a cell model to test the effects of periodic cooling on adipocyte dedifferentiation. Further studies should be performed with freshly isolated mature adipocytes to verify the effects of periodic cooling on adipocyte dedifferentiation. This might provide an innovative methodology to improve the production of DFAT cells from fat tissue.
Conclusion
Periodic exposure to cold promoted lipolysis and mature adipocyte dedifferentiation into fibroblast-like cells with high proliferation potential. This approach could be applied as a potential strategy to increase the efficiency of dedifferentiation from mature adipocytes. The DFAT cells generated from human fat tissue with high-efficiency may serve as a new cell source in stem cell therapy and regenerative medicine.
Footnotes
Acknowledgments
This work was financially supported by the Helmholtz Association through Helmholtz Graduate School for Macromolecular Bioscience (MacroBio, VH-GS-503), Helmholtz Cross Programme Initiative “Technology and Medicine - Adaptive Systems”, Helmholtz Virtual Institute “Multifunctional Biomaterials for Medicine (VH-VI-423), and programme-oriented funding.
This communication was presented at the Joint Meeting of the European Society for Clinical Hemorheology and Microcirculation, the International Society for Clinical Hemorheology, and the International Society of Biorheology, 2–6 July 2018, Krakow, Poland.