
Abstract
BACKGROUND:
The NCAM or CD56 antigen is a cell surface glycoprotein belonging to the immunoglobulin super-family involved in cell-cell and cell-matrix adhesion. NCAM is also over-expressed in many tumour types and is considered a tumour associated antigen, even if its role and biological mechanisms implicated in tumour progression and metastasis have not yet to be elucidated. In particular, it is quite well documented the role of the interaction between the NCAM protein and the fibroblast growth factor receptor-1 in metastasis and invasion, especially in the ovarian cancer progression.
OBJECTIVE:
Here we describe the isolation and preliminary characterization of a novel human anti-NCAM single chain Fragment variable antibody able to specifically bind NCAM-expressing cells, including epithelial ovarian cancer cells.
METHODS:
The antibody was isolate by phage display selection and was characterized by ELISA, FACS analysis and SPR experiments. Interference in EOC migration was analyzed by scratch test.
RESULTS:
It binds a partially linear epitope lying in the membrane proximal region of two fibronectin-like domains with a dissociation constant of 3.43
CONCLUSIONS:
According to our knowledge, this is the first completely human antibody able to interfere with this newly individuated cancer mechanism.
Introduction
The neural cell adhesion molecule-1 antigen (NCAM) or CD56 antigen is a membrane glycoprotein, a cell adhesion molecule structurally belonging to the immunoglobulin super-family with a molecular mass of 175 to 220 kDa under non-reducing conditions. NCAM is encoded by a single gene located on 11q23-24 that generates a wide variety of glycoprotein isoforms by alternative splicing. More than 24 distinct mRNAs of NCAM may be produced that are expressed preferentially in different tissues and stages of embryonic development.
The three major isoforms of NCAM differ mainly in overall size and in the type of membrane attachment. Two transmembrane-anchored glycoproteins of 140 and 180 kDa, respectively and a 120 kDa glycosyl phosphatidylinositol-anchored glycoprotein result from the use of different exons for the carboxyl terminus of the molecule. 120 kDa NCAM isoform is predominantly expressed in normal and well differentiated tissues, while both 140 and 180 kDa isoforms are found predominantly in less differentiated or malignant cell types [1, 2].
The extracellular domain has five IgG-like domains at the N-terminus and two fibronectin type III-like domains on the membrane proximal region and it is responsible for NCAM mediated cell-cell and cell-matrix interactions, by means of homophilic binding and various heterophilic interactions with other molecules.
NCAM is modified post-translationally by the addition of O-linked and N-linked oligosaccharides as well as
NCAM is expressed in several normal tissues, including natural killer (NK) cells, activated T cells, neurons, glial cells, neuroendocrine glands, skeletal muscle cells and cardiomyocytes. It contributes to a number of physiological processes such as embryogenesis and development, neuronal differentiation and synaptic plasticity [4].
The function of NCAM on human NK cells and T cells remains unclear, although its expression seems not to be essential for NK and T-cell development and function. Although a correlation exists between MHC-unrestricted cytotoxic function and expression of NCAM on lymphocytes, the NCAM antigen itself appears not to participate directly in NK killing of most target cells; notwithstanding, a recent report suggests that NCAM can mediate homotypic adhesion of NK cells to NCAM positive tumour cell lines and pathogen and tumour-cell recognition [5, 6, 7, 8].
NCAM is also considered a tumour-associated antigen, since it is overexpressed in almost all neuroblastoma [9], rhabdomyosarcoma [10], 98% of small cell lung cancer [11], 78% of multiple myeloma [12], and 53% of acute myeloid leukemia patients [13], some ovarian cancers [14], neuroendocrine cancers [15], pancreatic and gastric cancers [16], Wilms tumour [17] and Ewing sarcoma [18] patients also have elevated NCAM.
The functional role of NCAM in these tumours has to be elucidated, but actually a plethora of scientific data has emerged on the ability of NCAM to promote tumour progression and metastasis [14]. For example, NCAM upregulation subsequent to the loss of E-cadherin expression is implicated in the epithelial-mesenchymal transition (EMT) [19], a process that occurs when cancer cells disseminate from a primary tumour and infiltrate the surrounding tissue, leaving the tumour mass not only by gaining migratory and invasive capabilities but also by dissolving cell-cell contacts, known as tight and adherent junctions [20].
Moreover, recently, NCAM has been proposed to regulate the function of members of the fibroblast growth factor receptor (FGFR) family. These growth factor receptors are tyrosine kinases that trigger signaling cascades when bound by growth factors and other molecules, and aberrant FGFR signaling has been implicated in the progression of some cancers, including epithelial ovarian cancer, where its expression is associated with high tumour grade.
Published data have demonstrated that NCAM stimulates EOC cell migration and invasion in vitro and promotes metastatic dissemination in mice [14]. Several lines of evidence taken from in vitro and in vivo studies underline how this activity implicate the specific interaction between the adhesion molecule NCAM and members of the fibroblast growth factor receptor family, in particular FGFR1.
FGFR1 signaling induced by NCAM stimulation differs from the pathway initiated by other ligands such as fibroblast growth factor, FGF. In the absence of FGF, NCAM-mediated activation of FGFR specifically promotes characteristic cellular trafficking and receptor recycling that results in sustained FGFR signaling, leading to an enhanced cell migration with invasive and aggressive biological behavior [21].
NCAM-FGFR interactions are mediated by the second and the third Ig module of FGFR and the two Fibronectin-like type 3 (FN3) modules of NCAM. Even if the affinity of the NCAM-FGFR interaction (10
It was also presumed that association of NCAM to FGFR occurs when NCAM is polysialylated and has not a tight packing of homophilic interactions [23]. Moreover, the change in NCAM glycosylation in some tumours, in particular an increase of polysialic acids (PSA) levels within the fifth immunoglobulin domain of NCAM by two polysialyltransferases [24], appears to play a predominant role in the metastasis formation and tumour progression, by preventing NCAM interactions and downstream signaling [25, 26, 27].
NCAM over expression in different tumour types together with its role in promoting migration, invasion and metastasis, especially in EOC, make this antigen a potential novel drug development target. In this context, the development of an antibody-based target therapy could be a welcome addition to the available treatment choices.
In this paper, we describe the isolation and characterization of a novel, fully human antibody, in a ‘single-chain’ format specific for NCAM and also able to interfere with its interaction with FGFR1.
Materials and methods
Antibody phage library, NCAM antigen, human PBMC from healthy donors and cell lines
The IORISS naive human antibody library consists of a large array (more than 10
Peripheral blood mononuclear cells (PBMC) were isolated by Ficoll-Hypaque (Cedarlane, CL5120) density gradient centrifugation from heparinised blood samples of healthy donors (see Ethics approval and consent to participate section). Natural Killer cells were isolated via a magnetic cell sorter by using NK cell isolation kit II (Miltenyi Biotec, 130-092-657) according to the manufacturer’s protocol. The purity of isolated CD3-CD56
NK-92 is an interleukin-2 (IL-2, CellGS, GFH12) dependent Natural Killer cell line, and was a kind gift of Professor L. Moretta, (Director of the Istituto Gaslini, Genova, Italy) who obtained the cells from ATCC repository (ATCC; CRL-2407). The following human melanoma cells have different source: Me20842, Me30966, MelC and Mel501 are metastatic melanoma cell lines, LM2, LM3, LM4, LM5, LM7, INT10, INT18, INT30 and ATT are primary and metastatic tumour cells derived from lesions of melanoma patients and adapted to standard cell culture conditions. All these cells were kindly supplied by Dr Licia Rivoltini (Fondazione IRCCS Istituto Nazionale dei Tumori, Milan, Italy). Lymphoblastoid cells which include CCRF-CEM, HL60, K562, THP1 and MOLT-4 cell lines all obtained from the ATCC. A panel of carcinoma cell lines (breast, colon, and lung carcinoma) MCF7, HT29, LoVo, H69, H1299, H460, H292, H1650 and Calu were all purchased from ATCC. Cells were grown under standard conditions for mammalian cells cultures. The basic medium (BM) for cell culturing consisted of RPMI-1640 (Aurogene, AU-L0501) supplemented with 10% heat-inactivated foetal bovine serum (FBS, Lonza, DE14-801F), L-glutamine (L-Glu, Euroclone, EC13000D)), penicillin (100 U/mL)/streptomycin (100 U/mL) (P/S, Euroclone, 8ECB3055D).
Human osteosarcoma and Ewing’s Sarcoma cell lines, U-2OS, Saos-2 and SK-ES-1 were obtained from the American Type Collection. The TC-71 and 6647 cell lines were kindly provided by T. J. Triche (Children’s Hospital, Los Angeles, CA). IOR/BRZ was obtained from the Laboratorio di Oncologia Sperimentale, Istituto Ortopedico Rizzoli, Bologna, Italy. These cell lines are cultured in Iscove’s modified Dulbecco’s medium (IMDM, EuroClone, ECM0192L), supplemented with L-Glu (2 mM), P/S (100 U/ml) and 10% FBS. All cells in this study were maintained at 37
Human epithelial ovarian cancer cell lines, OVCAR3, Hey, A2780 WT, gently provided from Oncotech Tustin, CA, were cultured at 37
NCAM antigen used for biopanning strategies was a purified recombinant protein corresponding to the extracellular domain of human NCAM (neural cell adhesion molecule 1 isoform 1 precursor [Homo sapiens] NCBI Reference Sequence: NP_000606.3, aa 20-718 of the whole sequence) with 6xHis-tag (NCAMecd), produced in E. Coli by BiologicsCorp protein manufacturer (USA) on request.
Biopanning strategy for selection NCAM extracellular domain recombinant protein specific scFvB1
Specific antibodies in scFv format were isolated from IORISS phage library as previously described [28]. In brief, an aliquot of phages containing about 10
DNA characterization and sequences
Plasmid DNA from scFvB1 bacterial clone was digested with specific endonucleases; complementarity determining regions (CDR) were sequenced with an automated DNA sequencer (Eurofins Italia/, Italy) using scFvFor (5’TTGTCACGAATTCGGCCATGGCC 3’) and scFvRev (5’TTTGGGCTCGCTAGCTGCGG CCGC 3’) primers.
Soluble ScFv purification
Soluble scFv purification was performed as described by Ascione et al. [29]. For production of soluble scFvs, specific phage-TG1 E. coli infected cells were cultured at 37
NCAM extracellular fragments cloning and production
PBMC from a healthy donor were separated by Ficoll-Hypaque gradient centrifugation and then cultured in complete of RPMI 1640 medium with 20% FBS, 2 mM L-Glu, 100
After three and five days, the level of NCAM expression was evaluated by flow cytometry: an aliquote of cells was stained with the anti-CD56 PE (Becton-Dickinson, 555516) and anti-CD3 FITC (Becton-Dickinson, 349201) antibodies according the manufacturer’s instructions.
After five days, NK were isolated by magnetic separation, and mRNA was extracted using QuickPrep Micro mRNA Purification Kit (Analytik Jena, 845-KS-20300) according to the manufacturer’s instructions. mRNA (1
TG1 E.coli (supE hsd
Clones that showed the best induction were used for purification of proteins. Bacteria were grown in 100 mL 2
Elisa on recombinant NCAM proteins
96-well ELISA plates (ThermoFischer Scientific, 44-2404) were coated ON with 50
The reaction was visualized with 3,3’-5,5’- tetramethylbenzidine (BM blue, POD substrate, Roche, 10203700103) and stopped by adding 50
For the ELISA conducted in heat-denatured conditions, the plate was coated in parallel with 0.5
Determination of binding affinity
The determination of antibody affinity was performed by equilibrium saturation analysis using competition ELISA assay according to Rath et al. [30], with minor modifications as described by Gellini et al. [31].
A first ELISA step was performed to determinate the antibody concentration exhibiting 50% binding (EC50). Microtiter plates were coated with 50
A second ELISA step was performed to determinate the molar concentration of the free antibody at the equilibrium. Serial dilutions of the target antigen in solution were incubated in microtiter plates with the specific scFv at the determined EC50 (calculated from the first ELISA) and incubated for 1 hr to reach the equilibrium.
After the blocking step, the solution with the antigen at the varying concentrations and with the respective antibody at the determined EC50 (calculated from the direct ELISA) and allowed to incubate for about 60 min, was added to the antigen-coated wells together with a freshly prepared mixture consisting of an anti-FLAG M2, (Sigma, F3165) (12.5
Epitope mapping
Epitope mapping procedure was performed by Takis srl (Castel Romano, Italy).
A human NCAM peptide library was designed based on the sequence of NCAMecd 492-699 region and a total of 40 peptides were purchased from Genscript. The library was composed of peptides of 15 aa in length with 10 overlapping residues covering aa 492 to 699 of human NCAM1 precursor protein (NP_000606). Peptides were purchased in lyophilised form to avoid degradation.
Peptides were resuspended in 100% DMSO at 1 mg/ml concentration. Maxisorp 96 well ELISA plates were coated with the peptides diluted at 10 ug/ml concentration in 10 mM Carbonate buffer pH:9.6, in 100 ul/well volume, and incubated at 4
SDS-PAGE and western blotting
For analysis of purified scFvB1 one and five microliters were diluted in reducing SDS-PAGE loading buffer (NuPAGE
Purified NCAM fragments eluitions and corresponding bacterial lysates were fractionated by SDS PAGE. At the time of the choice of the best bacterial clone for protein production, bacterial lysates from not induced and 3 hours induced bacteria from different clones were fractionated by SDS/PAGE and transferred on nitrocellulose membrane. The membrane was blocked in 2% non-fat dry milk/PBS. After washing, the membrane was incubated 1 hr at RT with peroxidase conjugated anti-HIS antibody (Sigma). The reaction was detected with the DAB (3,3-Diaminobenzidine) reagent (Sigma).
For analysis of scFvB1 binding to recombinant NCAMecd protein in western blot assay, 0.5
NCAM detection on cell surface
The ability of scFvB1 to bind NCAM on the surface of cells was evaluated by flow cytometry investigations on intact/living PBMC and NK cells and a consistent panel of human cell lines which include CCRF-CEM and Molt4 (T lymphoblast), K-562 (chronic myelogenous leukemia), THP-1 (monocytic leukemia), HL-60 (promyelocytic leukemia), NK92 (Natural Killer cell line), H69 (small cell lung carcinoma), Calu (lung adenocarcinoma), H292 (lung epithelial), H1299 (nonsmall cell lung carcinoma), H460 (large cell lung cancer), H1650 (bronchoalveolar carcinoma), MCF7 (breast adenocarcinoma), HT29 and LoVo (colorectal carcinoma), U-2OS and Saos-2 (osteosarcoma), TC-71, 6647, SK-ES-1, IOR/BRZ (Ewing’s Sarcoma), Me20842, Me30966, MelC and Mel50 (metastatic melanoma cell lines), LM2, LM3, LM4, LM5, LM7, INT10, INT18, INT30 and ATT (are primary and metastatic tumour cells derived from lesions of melanoma patients), OVCAR3, SKOV 3, Hey, A2780 WT (ovarian cancer cell lines), Hela (cervix epitheloid carcinoma cell line).
Cells in exponential phase of growth were collected, carefully washed in PBS and pelleted.
For screening of scFv clones that were reactive on cells, about 2.5
NK cells were purified from PBMC from healthy donors and activated (where indicated) with 100 ng/mL IL-2 (CellGS, GFH12) for three days. Cells were then washed and marked with LIVE/DEAD
For PBMC, 1
Competition assay with FGFR1-Fc
OVCAR3 or A2780 cells in exponential phase of growth were collected, carefully washed in PBS and pelleted. About 2.5
Surface Plasmon Resonance assay
Surface Plasmon Resonance (SPR) experiments were carried out using a SensiQ Pioneer system [33]. The sensor chip (COOH5) was activated chemically by a 35
The analytes (NCAMecd and scFvB1) were dissolved in PBS buffer
Modeling and docking
Modeling of scFvB1 three-dimensional structure was carried out using the server I-TASSER [34, 35, 36]. I-TASSER modelling starts from the structure templates identified by LOMETS from the PDB library. LOMETS is a meta-server threading approach containing multiple threading programs, where each threading program can generate tens of thousands of template alignments. I-TASSER only uses the templates of the highest significance in the threading alignments, the significance of which are measured by the Z-score, i.e. the difference between the raw and average scores in the unit of standard deviation; among the top-threading templates, I-TASSER used 5YD5A, 5WN9A, 5WN9H, 2GKI, 6UL6 and 4BUHA. For each target, I-TASSER simulations generate a large ensemble of structural conformations, called decoys. To select the final models, I-TASSER uses the SPICKER program to cluster all the decoys based on the pair-wise structure similarity, and reports up to five models, which corresponds to the five largest structure clusters. The confidence of each model is quantitatively measured by C-score that is calculated based on the significance of threading template alignments and the convergence parameters of the structure assembly simulations. C-score is typically in the range of [
The best scFvB1 model was docked to the structure of the two FN3 domains of NCAM (Protein Data Bank 2VKW, chain A) using ClusPro [37] and HADDOCK [38], yielding similar top clusters of docked models.
Wound healing assay
Cells (3
Statistical analysis
For all ELISA experiments the cut-off value separating positive from negative samples was calculated as 3 standard deviation values (SD) above the mean, of the values obtained using the negative controls.
For Wound healing assay and flow cytometry competition assay data are expressed as means
Isolation of scFvB1 antibody. A. Schematic representation of panning cycle for the isolation of anti-NCAM scFvs from IORISS phage library. B. Screening procedures to identify monoclonal antibodies positive for NCAM antigen and able to recognise the antigen on cells. On the top, anti-NCAM ELISA carried out with supernatants of 90 bacterial colonies isolated from a polyclonal population of phage from three rounds of selection. The plate was coated with 0.5 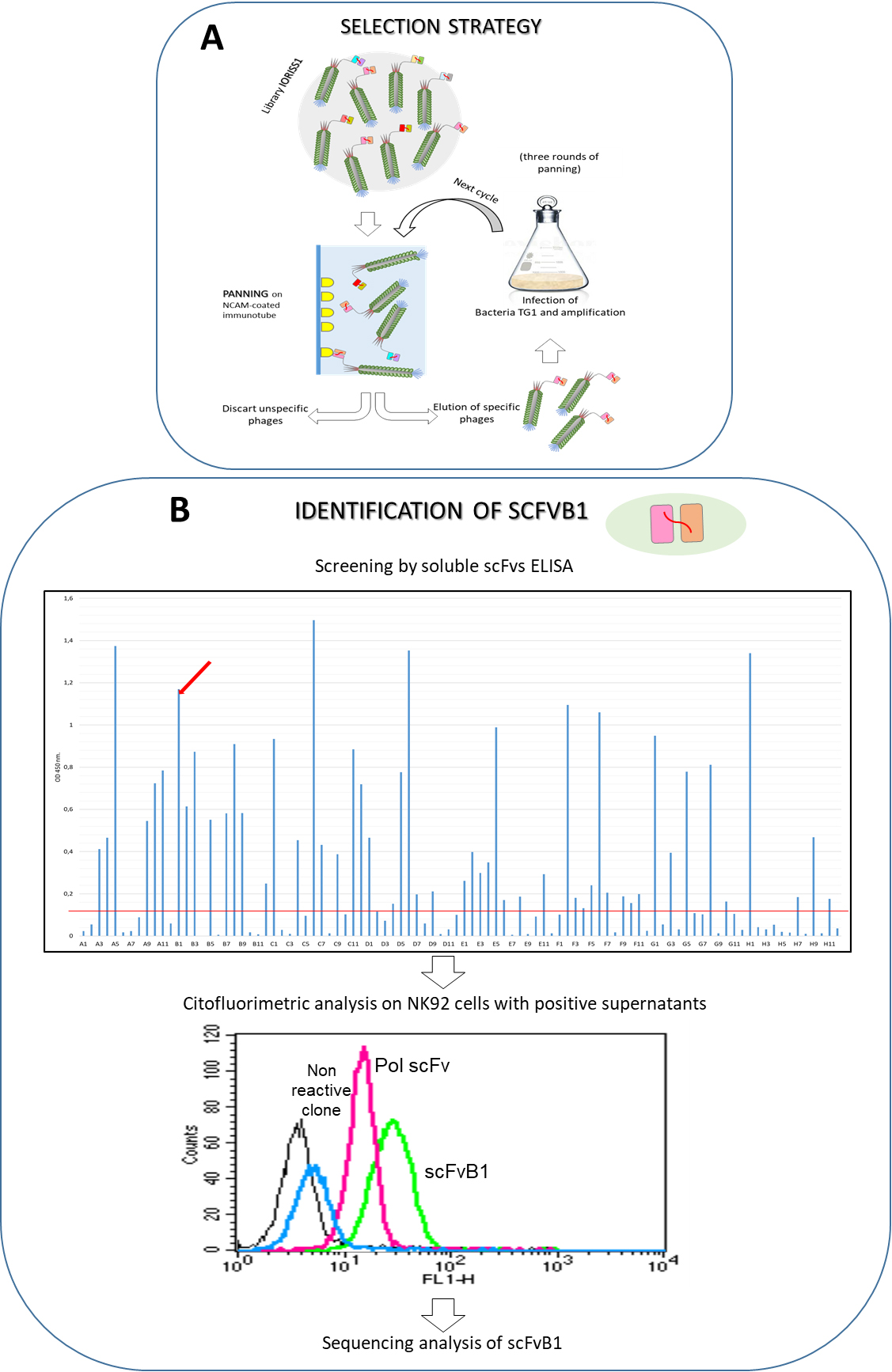
scFvB1 nucleotidic and amino acid sequence. In bold CDR1, CDR2 and CDR3 of VH (light blue) and VL fragments (light yellow). The sequence of the linker between the VH and VL fragments is in red.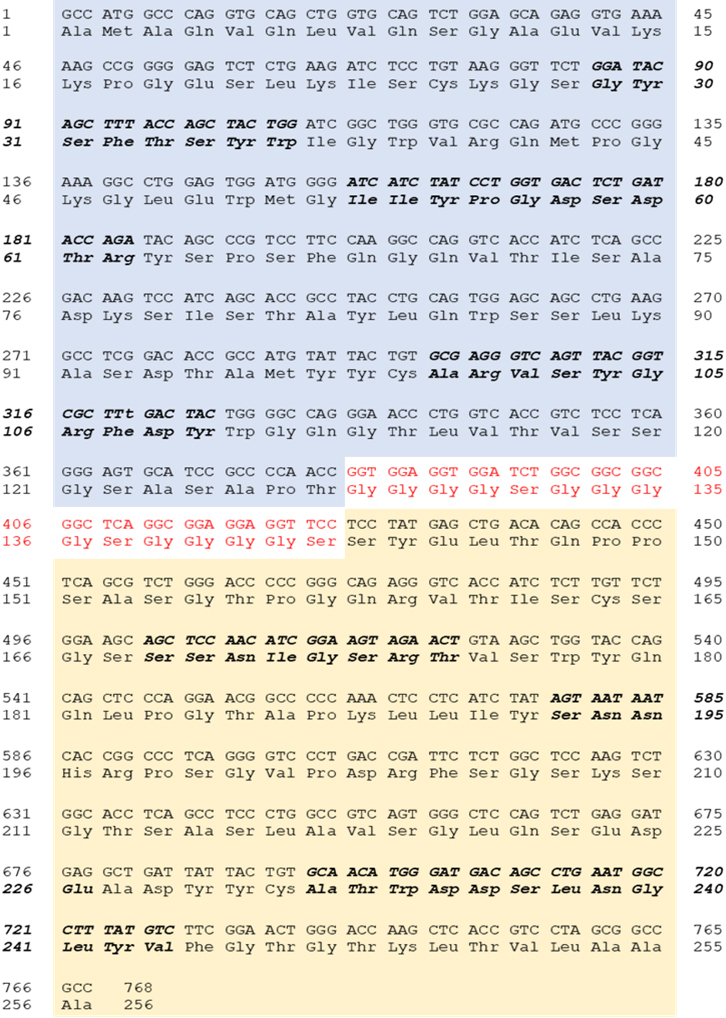
scFvB1 binding properties. A. Affinity binding (expressed as binding dissociation constant) calculated by a two steps ELISA, as described in methods. A first ELISA step (on the top) was performed to determinate the antibody concentration exhibiting 50% binding (EC50). Wells were coated with 0.5 ug of NCAMecd protein and then serial diluitions of scFvB1 (X axis, ng/ml) were tested. A second ELISA (down) was performed to determinate the molar concentration of the free antibody at the equilibrium. Wells were coated with 0.5 ug of NCAMecd protein and then serial dilutions of the same antigen (X axis, ng/mL) were incubated in solution with the specific scFv at the determined EC50 (calculated from the first ELISA). EC50 value from this second ELISA was used for the dissociation constant. B. On the top, schematic representation of the three cloned NCAM extracellular fragments (aa 20-307, aa 302-505, aa 492-699) and, down, results from ELISA of scFvB1 on the two available purified fragments (aa 20-307, aa 492-699). The plate was coated with 0.5 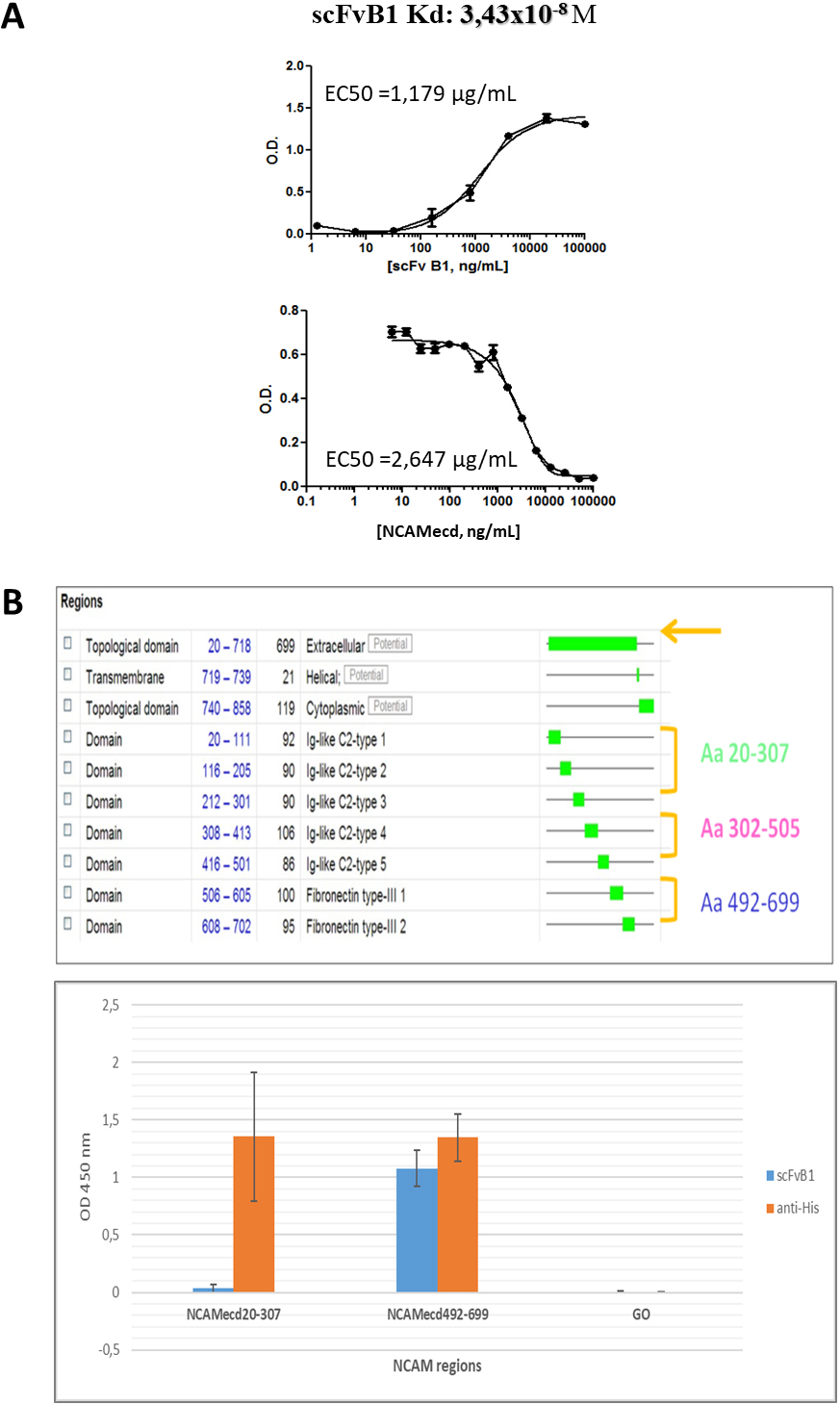
scFvB1 epitope analysis. A. WB analysis of ScFvB1 binding to NCAMecd antigen or irrelevant GO antigen in denatured conditions. NCAM antigen was revealed with anti-His antibody (lane 1) and scFvB1 (lane 4). GO antigen was only recognised by its specific scFvGO (lane 7). scFvB1 and scFvGO were detected with an anti-FLAG secondary antibody and then with anti-mouse-HRP conjugated antibody. Kilodaltons (kDa) of molecular markers (lane 3 and 8) are reported on the right. Nitrocellulose membrane was cut at level of the center of the two markers lane in order to perform different antibodies blotting and the reconstituted in the original manner for the figure. B. OD results from ELISA performed on 15 aa peptides (10 aa overlapping) mapping the NCAMecd 492-699 protein region coated at 10 ug/ml (NCAMecd 492-699 protein was coated as positive control). ScfvB1 was used at 10 ug/ml. scFvB1 was detected with anti-Flag AP conjugated (orange) or anti-6His HRP conjugated (blue) as secondary antibodies. w/o peptide: no coating. Control 
Flow cytometry reactivity. A. Dose-response binding of purified scFvB1 on NK92 cells. As control, NK92 cells were labeled with the secondary Abs alone (black). Green 0.25 ug/ml scFvB1, pink 2.5 ug/ml, light blue 25 ug/ml, blue 250 ug/ml, orange 1250 ug/ml. B. Flow cytometric analysis of purified scFvB1 (25 ug/mL) on purified native and IL-2 activated NK cells from healthy donor (right) in comparison with commercial anti-CD56 PE antibody (left). As control, cells were labeled with the secondary Abs alone or isotype control. C. Flow cytometric analysis of purified scFvB1 on PBMC isolated from healthy donor (left). The same sample was marked with commercial CD56 AlexaFluor647 and CD16PE in order to individuate the CD56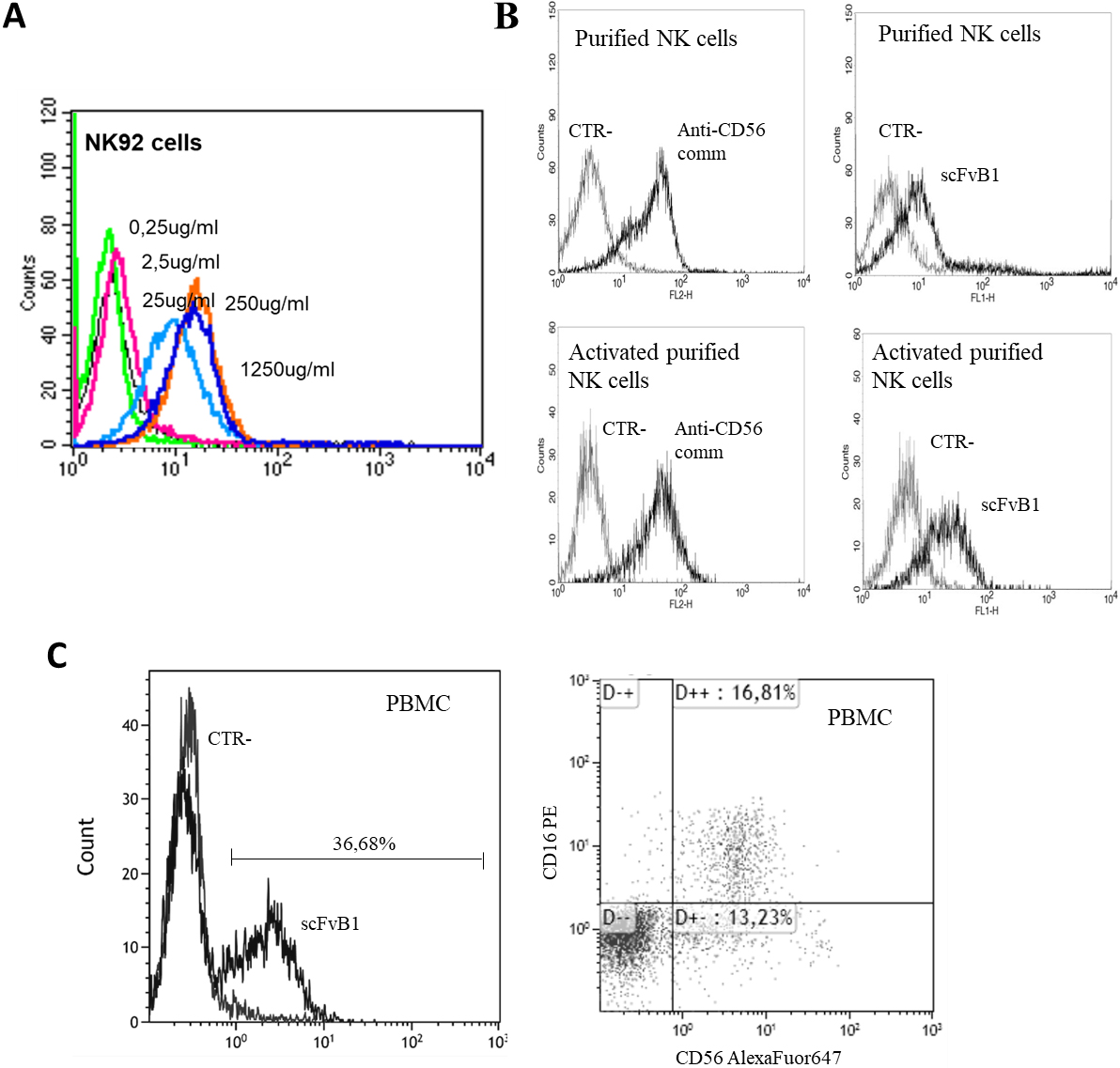
Isolation of human scFvB1 single chain antibody specific for recombinant and cell surface-exposed human NCAM by phage display technology
Human recombinant antibodies against NCAM were obtained by an in vitro affinity enrichment process commonly known as biopanning; in essence, an aliquot of phage-displayed scFvs representing the complexity of the naive IORISS library [28], has been subjected to cycles of absorption and elution to recombinant NCAMecd protein – used as a bait immobilised on immunotubes (Fig. 1A).
After three rounds of selections, we were able to isolate a phage-antibody population specifically recognizing NCAMecd. Figure 1B (top) reports results of monoclonal ELISA screening of soluble scFvs from single colonies obtained after plating bacteria TG1 infected with the enriched phages. Several of them proved to be specific for NCAMecd protein, while no signal was detected against recombinant Glucose Oxidase (GO) protein used as irrelevant antigen (data not shown).
Then, we analysed positive supernatants containing soluble scFvs for their reactivity on physiologically expressed NCAM, by flow cytometry on NK92 cells. Only one of tested scFvs was able to recognise NCAM on cell surface (Fig. 1B), therefore, hereafter, we will refer to this single NCAM-specific scFv, as scFvB1.
The DNA encoding for scFvB1 was sequenced; Fig. 2 shows scFvB1 DNA and amino acid sequence.
ScFvB1 was produced in bacteria and purified by metal chromatography using the six-histidine tag positioned at the C-terminal end of the antibody. After purification, scFvB1 was checked by SDS-PAGE (Supplementary Fig. 1) and the protein presents the expected molecular weight of about 27–30 KDa. An additional higher band at about 70 KDa is present, probably corresponding to scFv aggregates.
Evaluation of scFvB1 binding affinity and recognized epitope
First, the purified scFv was tested against the recombinant antigen used for selection panning in ELISA, in order to determine its binding affinity, that was calculated as Kd: 3.43
Unfortunately, we were able to obtain producing bacterial clones only for the first NCAM aa 20-307 and the third NCAM aa 492-699 fragments. These two NCAM fragments were produced and purified (Supplementary Fig. 2).
ELISA tests on these two recombinant regions showed that scFvB1 binds the region 492-699, that is close to the cellular surface (Fig. 3B) and involved in the interaction with the second and third Ig-like domain of FGFR1.
Since scFvB1 was able to recognize NCAMecd antigen also in denatured conditions in western blot analysis (Fig. 4A), we supposed that it binds to a linear epitope.
For this reason, we performed an epitope mapping with linear peptides of 15 amino acids with 10 amino acids overlapping regions covering the NCAMecd 492-699 region, recognised by the scFvB1.
Surprisingly, no signal was detected on any peptides in ELISA assay performed with 10 ug/ml of scFvB1 (Fig. 4B).
So we decided to conduct an ELISA on the heat-denatured NCAM protein (by boiling). In this case, the OD signal with scFvB1 was significantly lower but not abolished in respect to native NCAM protein, whereas the signal with anti-HIS antibody was almost the same on the two antigen (Fig. 4C).
Based on these results, we can state that the epitope recognized by scFvB1 is not completely linear, it being at least partially dependent on structure that SDS Page and boiling do not completely disassemble.
Evaluation of scFvB1 binding on tumor cells expressing NCAM antigen
First of all, a pilot flow cytometric analysis on NK92 cells with different concentrations of purified scFvB1 was performed to choose the optimal concentration for binding studies on different tumour cell lines. This experiment confirmed the ability of scFvB1 to bind naturally expressed NCAM on the surface of cells expressing the antigen and let us to identify the most balanced concentration of scFvB1 (25 ug/ml) for the successive wider cytofluorimetric studies (Fig. 5A).
scFvB1 was also tested on NK cells purified from a healthy donor before and after IL2-driven activation in order to promote a higher expression of NCAM, thus demonstrating the ability of scFvB1 to effectively detect target antigen level variations on cells surface (Fig. 5B). scFvB1 was also able to bind NCAM expressing cells in a more variegated population, that is PBMC purified from a healthy donor (Fig. 5C); in this case the percentage of NK population we found in PBMC was similar to the percentage detected with commercial fluorochrome-conjugated antibodies, thus confirming the good affinity and specificity of scFvB1.
Then, scFvB1 was tested on a large panel of different tumour cell lines available in our cellular bank. As shown in Table 1, scFvB1 was able to recognize NCAM as the commercial antibody control or better as for osteosarcoma cell lines and colon carcinoma. In particular, scFvB1 recognized 5/13 melanoma cell lines, 2/2 osteosarcoma cell lines, 3/4 Ewing sarcoma cell lines, 2/6 lung carcinoma cell lines (in particular small cell lung carcinoma), 1/2 colon carcinoma. Noteworthy, scFvB1 was positive on 1/5 EOC cell lines tested, OVCAR3 cells, according to literature data which report a percentage of NCAM-positive EOC cells of about 23% [14]. Supplementary Fig. 3 shows scFvB1 binding profiles to NCAM positive cell lines.
Flow cytometry determination of the binding level of scFvB1 on different tumor cell lines
Flow cytometry determination of the binding level of scFvB1 on different tumor cell lines
A commercial anti-CD56PE was used as positive control, while an scFv against glucose oxidase antigen (GO) was used as negative control. Stained cells:
Since scFvB1 was able to recognize the region of NCAM involved in the interaction with FGFR1, we tried to verify whether it was also able to interfere with this binding.
Since NCAM-FGFR1 interaction, intracellular signaling and subsequent effects on migration are well documented for epithelial ovarian cancer (EOC), we focus our experimental evaluation on the NCAM-positive OVCAR3 cell line.
First, we tested the ability of scFvB1 to compete with exogenous FGFR1-Fc recombinant protein for the binding to OVCAR3 cells. As reported in Fig. 6 and Supplementary Fig. 4, the incubation of OVCAR3 cells with scFvB1 reduces the binding of FGFR1-Fc to the cell’s surface, thus suggesting a competition between scFvB1 specific for NCAM1, and FGFR1-Fc. The competition effect seems to be antigen-specific since we didn’t observe the same effect with the irrelevant antibody scFvGO [32]; on the contrary, we observed the opposite effect, such as an increase of FGFR1-Fc signal. This is a counterintuitive effect we observed in other occasions (data not shown), probably caused by a binding stabilization in turn due by interactions between the proteins present in the sample. Moreover, On the basis of these results, we evaluated the ability of scFvB1 to interfere between NCAM and FGFR1 by studying the protein-protein interaction by surface plasmon resonance experiments. First we confirmed literature data of about 10
FGFR1-Fc competition assay by flow cytometry on OVCAR3 cells. OVCAR3 cells were marked with FGFR1-FC protein (10 ug/ml, 0.125 uM) and anti-human IgG FITC as secondary antibody, alone or in presence of scFvB1 or scFvGO (50 ug/ml, 1.66 uM). Represented histograms were the average of four independent experiments; Bars represent standard error. Star represents statistically significant difference, 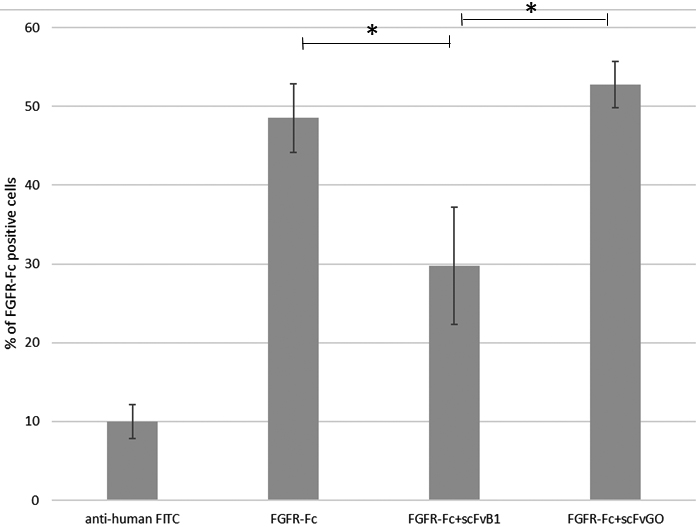
SPR competition experiment. Interaction between NCAMecd and immobilised FGFR1-Fc in the presence of scFvB1. Sensorgrams measured upon injection of 5 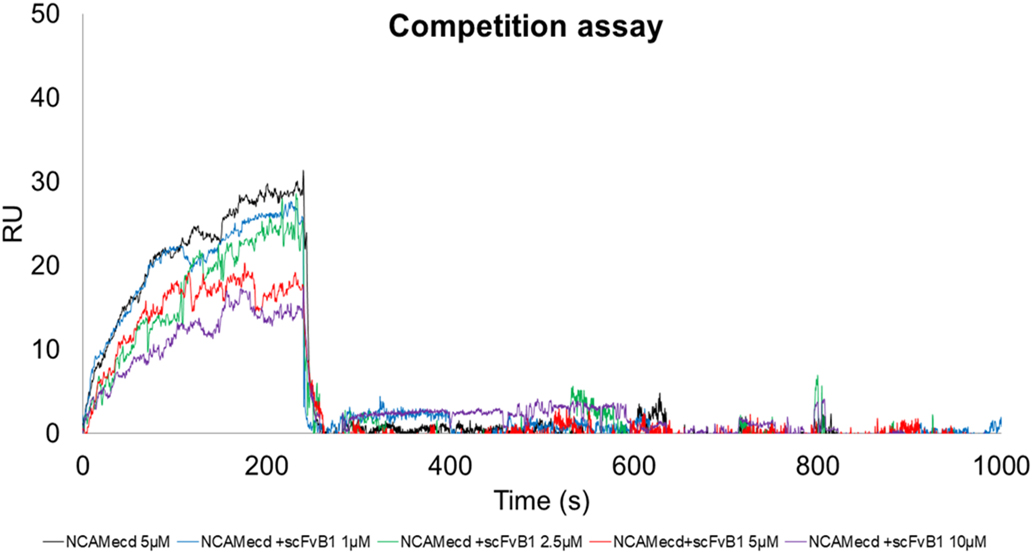
Docking analysis. A. Docking of NCAM-FN3 structure (blue) and scFv model (green), performed with HADDOCK 2.0. B. Close-up of the interface of the NCAM-FN3 – scFv docking, with the residues involved in the interaction between the proteins: residues Q507, E509, Y511 and Q516 belonging to the FRM region of 1FN3, and Y672, Q679, K681and K683, belonging to the FGL region of 2FN3, are among the residues establishing interactions with scFv. Figure designed with Pymol.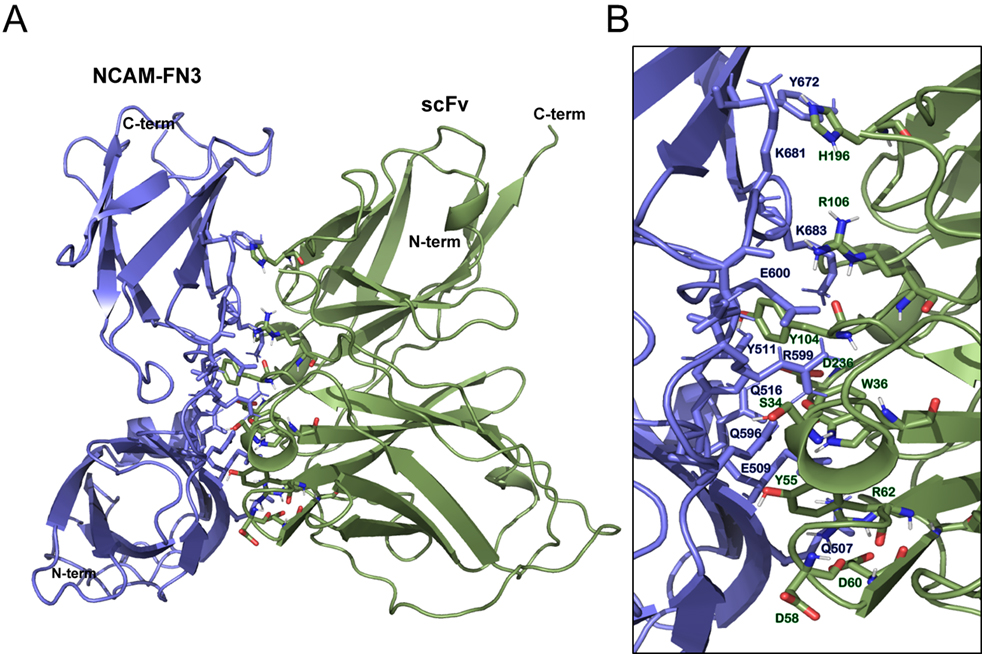
Wound healing assay. A. Graphic representation of percentage of scratch area repopulation at 48 hrs of OVCAR3 cells treated with scFvB1 and scFvG0 (40 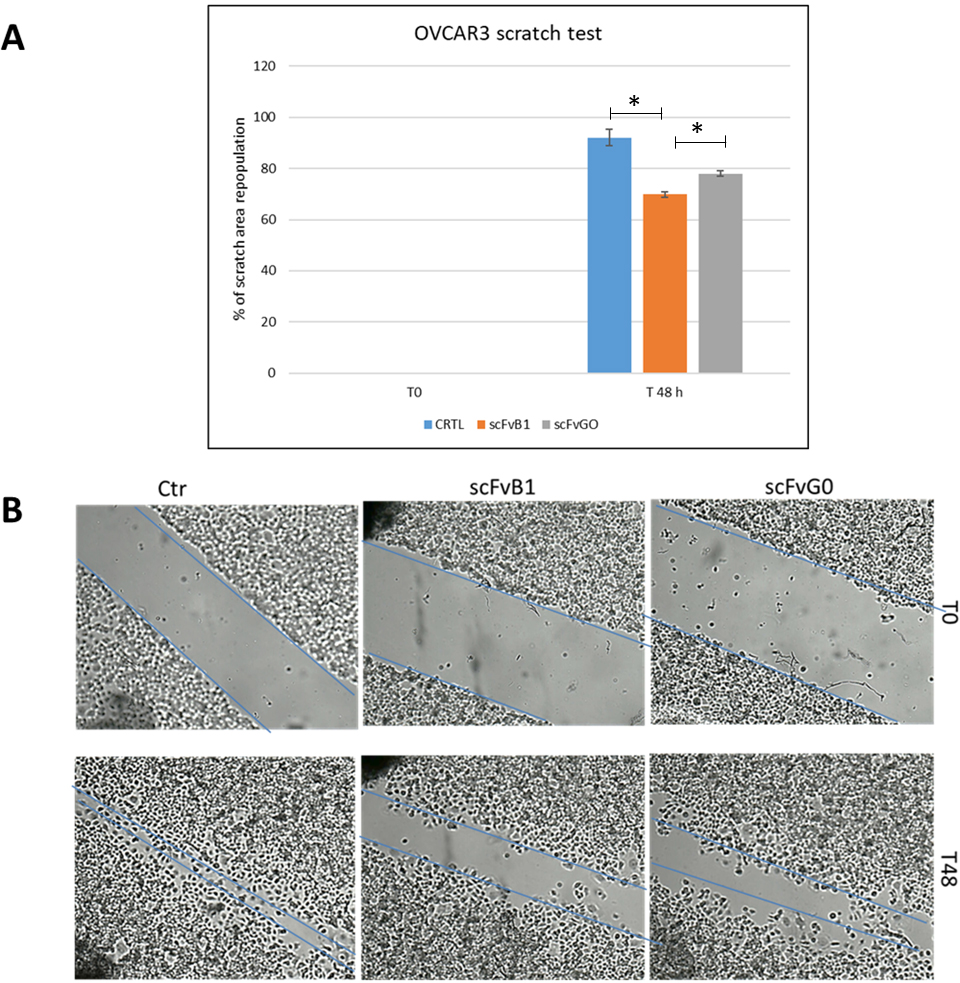
In order to further investigate the interesting ability of scFvB1 to interfere with NCAM-FGFR1 binding, we performed a docking computational study. The top model (model 1) of scFvB1 three-dimensional structure predicted by I-TASSER had a C-score of 0.94, estimated TM-score of 0.84
The best scFvB1 model was docked to the structure of the two FN3 domains of NCAM; the best cluster of structures (cluster 7), has a HADDOCK score of
scFvB1 interferes with migration of EOC cells in vitro
Finally, we decided to evaluate whether the molecular interference in the interaction between NCAM and FGFR1 mediated by scFvB1 translates in an inhibition of EOC cell migration. OVCAR3 cells were scratch wounded and treated with scFvB1 or scFvGO as described in materials and methods. First significant results were obtained after 48 hrs; as reported in Fig. 9, repopulation of the scratched area was significantly lower when cells were treated with scFvB1 compared to the other controls, suggesting a partial inhibition of cells migration process. Experiments were conducted in triplicate; Fig. 9 reports images from a representative experiment.
Discussion
NCAM has been shown to have a high level of expression in a variety of malignancies and has a role in tumour biology, in particular migration and metastasis [39]. This aspect makes this cell surface molecule an attractive antigen target for immunotherapy and diagnostic approaches in spite of its expression also on some normal tissue and cells including NK, cardiomyocytes and peripheral neural tissues. Indeed, investigators were not impeded to pursue an anti-NCAM approaches for cancer therapy by this potential drawback, as evidenced by a steadily rising number of indications and strategies are being published.
To date, only three non-mouse anti-NCAM antibodies are actively described, all of them developed for an antibody-drug conjugated (ADC) approach:
Lorvotuzumab (huN901, humanized N901) linked to the cytotoxic maytansinoid effector molecule DM1 via a disulfide linkage (mertansine) (LM, huN901-SPP-DM1, BB-10901, IMGN901) that is being evaluated in clinical trial for different solid tumours [40] and for multiple myeloma [41] with good results [42]. m900 and m906 conjugated with pyrrolobenzodiazepine dimer (PBD) (m900PBD and m906PBD), both human scFvs tested on neuroblastoma cell lines, which exhibited different killing activities, correlated both to their different ability to be internalised and to the heterogeneity of dynamics of NCAM on the different cell lines [43]. Promiximab, a chimeric anti-NCAM antibody linked to two different drugs, which proved to be a good efficacy in small cell lung cancer (SCLC) cell line xenograft models, even if not all SCLC cell lines were bound by the antibody [44, 45].
However, recently a great interest is arising on the interaction between the NCAM1 and FGFR1 and the consequent induction of cell migration and metastatic dissemination, with particular attention to the EOC. Clearly, the interplay between an adhesion molecule and a growth factor may represent a new target for novel drug development for EOC, or other tumors that share this mechanism at the basis of their genesis and development, including during EMT [46, 47, 48].
These observations open the possibility to evaluate another mechanism of action for anti-NCAM antibody-based therapeutic strategies. In this context, the generation of a number of different highly specific antibodies against human NCAM is a prerequisite for the development of appropriated therapeutic strategies.
Here, we describe a new NCAM-specific fully human single chain antibody named scFvB1, endowed with good binding affinity and capable to selectively recognize various NCAM expressing tumor cell lines.
Our data indicate that the scFvB1 binds to a complex epitope that is not completely linear; in fact, even if scFvB1 was able to recognize the antigen in both WB and ELISA under reduction conditions, no reactivity was observed on overlapping linear peptides derived from the primary NCAM sequence. To this regard, it is important to point out that some antibodies recognising linear epitopes will bind to a specific spatial conformation of the linear epitope and thus it still recognises a conformational-specific epitope in the case of native proteins [49].
At the same time, there are different examples of antibodies toward conformational epitopes that showed ability, even if limited, to bind the target protein in Western blot [50].
Interestingly, we demonstrated that scFvB1 recognised the region of the two fibronectin like domains on extracellular domain of NCAM which is involved in the interaction with FGFR1 [21, 22].
This binding was also proved to translate in an interference into the molecular interaction between NCAM and FGFR with two different competition assays. The fact that scFvB1 doesn’t completely abolish, but only reduces the fluorescent signal of FGFR1-Fc, in spite of the higher binding affinity of scFvB1 for NCAM compared to FGFR and NCAM, can have different explanations. It could be the effect of a partial competition, which means that the epitope of scFvB1 is involved in the interaction but not limited to it. However, the phenomen may also be determined by the fact that FGFR1-Fc binding to OVCAR3 is not strictly limited to the NCAM molecule; this is a reasonable explanation since it is known that FGFR forms homophilic and heterophilic bindings with other molecules on cells surface.
Importantly, treatment of OVCAR3 cells with scFvB1 caused a partial inhibition of cells migration.
Docking analysis provided us the possible structural explanation for the competition ability of scFvB1, since the best obtained model of the interaction between scFvB1 and the structure of the two FN3 domains of NCAM displays a series of intermolecular contacts that involve amino acids of FRM and FGL regions of NCAM that are responsible for FGFR1 interactions.
Interestingly, scFvB1 amino acids involved in antibody-NCAM antigen interactions are located almost all in the VH chain; this finding is in line with several previous studies pointing out the central role of the heavy chain in antigen binding and specificity [51, 52, 53].
According to our knowledge, scFvB1 is the first completely human antibody described so far, with the potential to specifically target and interfere with the NCAM/FGFR interplay, known as involved in multiple progression cancer mechanisms. In the context of innovative therapeutic strategies, this context, a blocking antibody specific for NCAM could probably be more safe in respect to an inhibitor of FGFR1 receptor, since NCAM expression occurs specifically in transformed cancer cells while no antigen was observed in corresponding normal tissues.
Clearly, a deeper study of scFvB1 is necessary, but, it is important to underline that the structural flexibility of the scFvB1 format allows to further optimize the properties of scFvB1, for example by improving its affinity or steric hindrance and its competitive function; furthermore, scFvB1 can be considered a starting block to develop antibody constructs with a more balanced molecular weight in order to satisfy both the requirements of adequate pharmacokinetics and effective tumor penetration.
These properties make it a starting molecule for the development of potential therapeutic applications.
Recent studies are underlying the role that NCAM can have in cell migration and metastatic dissemination, via FGFR1 receptor. Different papers also examined in depth the structural basis for this interplay. We isolated a completely human scFv, named scFvB1, that was able to specifically bind NCAM molecule and interfere with the FGFR1 binding as demonstrated by functional and computational results. The structural flexibility of the scFv would allow to optimize its functional and kinetics properties, while its completely human origin is already an essential starting point for the development of therapeutic agents.
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Authors’ contributions
MF isolated scFvB1; GC performed SPR experiments and docking analysis; AA performed affinity experiments and ELISA for region mapping; MLD carried out characterization of scFvB1 on cell lines; EP and RR carried out OVCAR3 scratched tests; MA was on charge with FACS analyses; CR performed SDS PAGE and western blot analysis; MG was in charge to purify scFvB1; AB analyzed scratched tests images; SB performed recombinant protein purification; SV supervised the study design on the basis of the scFv isolation; AM designed experiments, wrote the paper and coordinated the study. All authors have seen and agreed to the final version of the paper.
Ethics approval and consent to participate
Human specimens used in some experiments described in our manuscript are PBMCs from healthy donors. Multiple stocks of human peripheral blood mononuclear cells (PBMCs) isolated from a large number of healthy donors (
They were volunteers; accordingly to the Italian law on this matter (Legislative Decree of the Italian Ministry of Health of January 25, 2001, published on the Official Gazette of April 3, 2001) donor ages ranged between 21–60 years for women and 21–65 years for men. There have been no restrictions for blood donors as to sex and ethnic background, except for children, mentally disabled individuals and pregnant women. All donors were healthy, non-medicated consulting adults, and were prescreened for exposure to infectious agents. Blood was obtained by venipuncture following standard medical procedures using sterile techniques and a compression bandage in the area of blood removal. A record of the donor and hematocrit result is stored on file at the Transfusion Center. Research investigators do not have any access to personal data or other information concerning the donors in agreement with the Italian law Number 675 of December 31, 1996.
Our study did not require an ethical approval for the use of described cell lines for the evaluation of the ability of our scfv to recognize CD56 on their surface, since: the first legislation that is applicable to our study was emanated by the Data Protection authority in 2012 (General authorization for the treatment of personal data for research uses. Official Gazette of Italian Republic n. 72 published on March 26, 2012) and cell lines described in our study were obtained in a previous time. Moreover, there is not possibility to find the identity of the patients from which cell lines originated.
Funding
This work was supported by research funds of Italian Ministry of Health with the Grant number: GR-2010-2306432.
Supplementary data
The supplementary files are available to download from https://dx-doi-org.web.bisu.edu.cn/10.3233/HAB-200431.
Footnotes
Acknowledgments
We thank Dr. Massimo Sanchez and Dr. Valentina Tirelli of the ISS Service of instruments and core facilities for their technical support.
Conflict of interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.