
Abstract
Background:
Experimental studies have demonstrated that aluminum is an environmental toxin that induces neuroinflammation and the development of Alzheimer’s disease.
Objective:
In this report, we investigated the beneficial effect of a combination of resveratrol and curcumin to reduce aluminum-induced neuroinflammation.
Method:
We employed both an in vivo model of aluminum-induced neuroinflammation and an in vitro aluminum stimulated cultured PC-12 cells. Neuroinflammation in rats was assessed by measuring the expression of β-secretase, amyloid-β protein precursor, and γ-subunits (PS-1 and PS-2), along with the inflammatory COX-2, Il-1β, Il-1α, and TNF-α. Furthermore, we measured the expression profiles of neuro-protective Apurinic/apyrimidinic endonuclease 1 (APE1) protein and let-7c microRNA. In parallel, PC-12 cells were treated with 0.5 mM aluminum to induce a neuroinflammation-like state. In addition, curcumin effect, as a selective COX-2 expression inhibitor, was detected in a time course manner.
Results:
An overall significant attenuation of the inflammatory markers, as well as a decrease in the amyloidogenic mediators, was observed in resveratrol-curcumin treated rats. The therapeutic effect was also confirmed by transmission electron microscopic analysis of the brain cortexes. APE1 was significantly induced by resveratrol-curcumin combination. Both in vivo and in vitro studies indicated that Let-7c expression is significantly reduced after aluminum stimulation, an effect that was partially suppressed by co-addition of either resveratrol or curcumin and totally restored to the normal level by their combination.
Conclusions:
The present study clearly indicates the synergistic and therapeutic effect of a resveratrol-curcumin combination. We also show that both compounds exert beneficial effect either cooperatively or through differential molecular mechanisms in counteracting aluminum-induced neuroinflammation.
Keywords
INTRODUCTION
Alzheimer’s disease (AD) accounts for 60% to 70% of cases of dementia [1]. It is the most prevalent neurodegenerative disease of the elderly comprising complex neuro-biochemical and neuropathological events. It is characterized by regional specificity of neural aberrations associated with higher cognitive decline with a progressive loss of memory, extracellular amyloid-β deposits [2], and neurofibrillary tangles. The neurofibrillary tangles are formed by hyperphosphorylation and abnormal deposition of tau proteins [3]. The etiology of this disorder is multifactorial including genetic factors, head trauma, oxidative stress, inflammation, and environmental factors including aluminum (Al) toxicity. Al is the most abundant neurotoxic metal on earth, widely bioavailable to humans and repeatedly shown to accumulate in AD-susceptible neuronal foci.
Al induces neuronal loss, ultrastructural alterations in the different brain regions, and biochemical modifications that are implicated in AD development [4]. An elevated amount of deposited Al has been also reported in AD brain [5, 6]. Furthermore, it has been shown that Al crosses the blood-brain barrier through a specific transferrin receptor [7] and induces profound memory loss via disruption of various normal neuronal functions [8, 9]. At the molecular level, Al induces genetic and epigenetic modifications that alter genes expressions [10]. Phenotypic changes including protein misfolding and plaques depositions are involved in AD pathology [11].
Al augments intracellular oxidative stress by various mechanisms including potentiation of transition metals pro-oxidant activities [12] and distribution of the mitochondrial membrane potential [13]. Moreover, Al is a well-established neurotoxin for induction of neuroinflammation and consequently neurodegeneration in rodent models.
The efficacy of curcumin, a dietary hydrophobic polyphenol found in the curry spice turmeric, has received a lot of attention in several preclinical trials for the treatment of neurodegenerative diseases because of its safety and low cost. Curcumin is a pleiotropic molecule that has been shown to directly bind to and limit aggregation of specific amyloid-β that is characteristic of many neurodegenerative diseases. It also restores balance of the inflammatory system being an inhibitor of cyclooxygenase-2 (COX-2) expression, fosters the heat shock system, and induces the anti-oxidant elements [14]. In fact, curcumin effectively suppresses cytotoxicity and inflammation and inhibits amyloid aggregations [15], which are the characteristic features of AD. Furthermore, it has been reported that curcumin crosses the blood-brain barrier due to its lipohilic characteristics and reduces the existing senile plaques [16]. Resveratrol is also a potent antioxidant and anti-inflammatory molecule [14] against neurodegenerative diseases [17]. It has a molecular weight of 228 Da, therefore it crosses the blood-brain barrier via trans-membrane diffusion [18]. Resveratrol is reported to play a prominent role in the prevention of different neurodegenerative diseases [19]. Since oxidative stress appears to be closely related to major neuronal pathologies, resveratrol treatment has been tested with positive results in neurodegenerative disorders such as AD, Huntington’s disease, cerebral ischemia, Parkinson’s disease, epilepsy, and amyotrophic lateral sclerosis [20]. We and others have uncovered the role of DNA repair apurinic/apyrimidinic endonuclease 1/reducing Factor-1 (APE1/Ref-1) protein as a neuro-protective component against Al-induced neuroinflammation [21–24]. We have also shown that natural products like curcumin or resveratrol mediate neuroprotection, at least partially by maintaining high APE1 expression [21, 22].
Altered micro-RNAs (miRNAs) expression is now correlated with altered neuroinflammatory protein expression and hence altered functions that are implicated in development of neurodegeneration reviewed in [25, 26]. Let-7c, a highly expressed miRNA in the brain that plays a physiological role in normal development, has been recently reported to suppress microglial activation and protect against brain damage from ischemia [27]. We have previously showed that curcumin and valproic acid combination induce neuronal recovery in lipopolysaccharide (LPS)-induced rats through significant induction of five let-7 family members includinglet-7c [22].
According to this background, we designed the current study to investigate the neuro-therapeutic effect of the resveratrol/curcumin combination against Al-induced oxidative damage, molecular alterations, and neuroinflammation in rats.
∥
MATERIALS AND METHODS
Animals
Sixty-seven male adult Wistar rats (100–150 g) were used to conduct the present study. Animals were purchased and housed at the Medical Research Institute at Alexandria University, in which the European principles of laboratory animal care ethics were followed in all experimental protocols. Rats were maintained under controlled temperature (25±2°C) and constant photoperiodic conditions (12:12-h daylight/darkness). The dams had free access to water and standard commercial chow containing 20% protein, 54% carbohydrate, 4.5% fiber, 4% lipids, 7% ash, and 10% moisture.
Neuroinflammation was induced in male Wistar rats using 25 mg/kg aluminum chloride (Al) daily by oral gavage for one week then the dose increased to 150 mg/kg for three weeks. Al was administered gradually to avoid acute systemic toxicity and animal death. In parallel to the induced group, 15 rats received an empty vehicle and represented the mock-treated group. Induced rats were subsequently divided into 4 major groups (10 to 14 each) and received oral gavage treatment as follows: empty vehicle (saline solution), resveratrol (200 mg/kg), curcumin (200 mg/kg), or their combination three times/week for four weeks.
Assay of oxidative stress related markers
Total antioxidant capacity assay (serum)
Total antioxidant capacity in sera samples were assayed following Koracevic et al., [28] protocol.
Measurement of total lipid peroxidation
Part of dissected mid-brain tissue (10% w/v) was washed with saline solution, minced, and homogenized in ice-cooled potassium phosphate buffer (50 mM pH 7.5). Level of lipid peroxidation was determined according to Ohkawa et al. [29]method.
Glutathione reduced determination (GSH)
Briefly, part of the brain tissue (10% w/v) was washed with saline solution, minced, and homogenized in ice-cooled buffer containing: 1.15% KCl, 0.01 M sodium phosphate buffer pH 7.4. Concentration of GSH was determined as described byEllman [30].
Glutathione-s-transferase (GST) activity
Brain tissue was homogenized in ice-cooled buffer (100 mM potassium phosphate, pH 7.0 containing 2 mM EDTA) per gram tissue. GST was assayed according to the Habiget et al. [31] method.
Superoxide dismutase determination (SOD)
The assay was preformed according to the method described previously [32].
Catalase activity
Part of brain tissue was homogenized in 5 to10 ml of cold homogenization buffer (50 mM potassium phosphate, pH 7.4, 1 mM EDTA, and 1 ml/L Triton X-100) per gram tissue. Catalase activity was assayed according to the Aebi [33] procedure.
Assessment of acetylcholine esterase activity
The Ellman et al. [34] method was applied to assay acetylcholinesterase (AChE) activity. Briefly AChE reacts with acetylthiocholine iodide (first substrate) and 5, 5-dithiobisnitrobenzoic acid (second substrate) to give a yellow color that is read at 405 nm. Absorbance was measured at 405 nm every two minutes. AChE activities were calculated and represented as mU/mg protein.
Electron microscopy of brain tissue
Specimens assigned for transmission electron microscopic examination were fixed in 4F, 1G solution. Dehydration in ethanol series was followed by applying propylene oxide (transition solvent) then infiltration and embedding in a liquid resin (epoxy). Resin blocks were then ultra-sectioned. Fifty to seventy nano meter thick sections were mounted onto metal mesh grids and stained with uranyl acetate and lead citrate.
Preparation of whole-cell lysates
Part of frozen mid-brain tissues or PC-12 cells was homogenized in total cell lysis buffer that composed of HEPES (50 mM pH 7.4), Triton X-100 (1% v/v), EDTA (4 mM), sodium fluoride (1 mM), sodium orthovanadate (0.1 mM), tetrasodium pyrophosphate (1 mM), PMSF (2 mM), leupeptin (10 μg/ml), and aprotinin (10 μg/ml). The cells’ homogenate was vortexed and frozen in ethanol/dry ice bath. Cycles of thawing/freezing were repeated twice. Homogenate was divided into aliquots of 300 μl each and stored at –80°C for further analyses.
Western blotting analysis
According to the method described by Burnette [35], APE1, COX-2, Aβ40, and β-actin immunoblots were performed on prepared tissue extracts. Briefly, 50 μg protein lysates from each sample were mixed with 2x loading buffer (130 mM Tris-HCl, pH 8, 30% (v/v) glycerol, 4.6% (w/v) SDS, 0.02% Bromophenol blue, 2% DTT), boiled for 5 min then cooled at 4°C for 5 min. Samples were separated on 12% SDS-PAGE mini gel and run at 100V. Proteins were transferred to nitrocellulose membrane at 70V for 1 h at 4°C. The membrane was washed three times with TBST (50 mM Tris-base, pH 7.5, 150 mM NaCl, 0.05% Tween 20) and incubated in blocking buffer (TBST containing 5% carnation nonfat dry milk) for 1 h at room temperature then incubated overnight in primary antibody to either APE1 (sc-17774), COX-2 (H-62), or Aβ40 (NB300–225). After washing with TBST, antibody binding was detected following appropriate secondary antibody application using chemiluminescence detection. Equal loading was confirmed by probing with β-actin (sc-81178) monoclonal antibody.
Induction of neuroinflammation like state in PC-12 using aluminum chloride
PC-12 cells were grown on complete culture medium containing DMEM, horse serum (7%), calf serum (3%), and penicillin/streptomycin (50 units/50 μg of each per ml).
Cells were stimulated with different concentrations of Al for 6 and 24 h, respectively. The optimum sub-lethal concentration of Al that induced oxidative stress was 0.5 mM. PC-12 cells were treated with 0.5 mM Al and treated with either resveratrol (100 μM), curcumin (100 μM), or their combination for 6 and 24 h. Treated cells were collected and lysed for total RNA and protein extraction. Experiments were done under standard conditions in the presence of proper controls.
Isolation of total RNA, reverse transcription, semi-quantitative, and quantitative (Q) PCR
Total cellular RNA was extracted from dissected brain tissues that were stored in RNAlater solution as Chomczynski and Sacchi described [36], using Biozol RNA Isolation reagent.
Alterations in the steady-state mRNA levels of neuroinflammation-associated genes were determined by reverse-transcriptase PCR analysis. Reverse transcription kit, Thermoscientific RevertAid, was used for converting total mRNA into cDNA. The cDNA synthesis reaction was performed as described in the pamphlet. The synthesized cDNA was then used for the amplification of target genes using specific primer sets as shown in Table 1. The primers were then subjected to PCR cycles; each cycle consists of denaturation, annealing, and extension steps. Primers set combination for the studied genes and the corresponding accession numbers are shown in Table 1.
Sequences of the primers of the analyzed genes with the corresponding accession numbers
Extracted miRNA was reverse transcribed using Ncodetrademark Vilotrademark miRNA cDNA synthesis kit. The resulting cDNA was amplified by PCR using miRNA specific primers and NCodetrademark SYBR® GreenERtrademark.
The relative levels of miRNA let-7c, COX-2, and APE1 expressions were calculated from the relevant signals by normalization with the signal of U6 for let-7c and of β-Actin expression for other genes. The fold change of let-7c and t expression were calculated based on the threshold cycle (CT) value using the following formula: 2–ΔΔCT [37].
Statistical analysis
We analyzed in vivo samples individually without pooling or mixing different samples. Each experiment was repeated at least two to three times independently and illustrated graphs represent the mean and the slandered errors. The difference between groups was analyzed by one-way analysis of variance (ANOVA), SPSS program, and the difference considered significant when compared to control either at p < 0.01, p < 0.001, or p < 0.0001.
RESULTS
Curcumin and resveratrol combination improves the ultrastructure and biochemical markers in rat brains
Resveratrol and curcumin are well studied as antioxidant compounds in different experimental models. The efficacy of this combination was assessed by analyzing the brain ultrastructure as well as by measuring the antioxidant capacity and AChE activity.
Experimental rats were treated with Al for 4 weeks by oral gavage and consequent pathological alterations in the brain cortex were assessed at ultrastructural level using electron microscopic examination. The results, as shown in Fig. 1, indicated progressive neuroinflammation and accumulation of autophagic vacuoles in induced brains compared to mock-treated ones. Using resveratrol or curcumin alone as a treatment partially ameliorated Al-induced damage when administered for four weeks, while their combination clearly restored the brain structural organization, almost close to mock-treated animals with even detectable attempts at regeneration(Figs. 1 and 2).

Transmission electron microscope (TEM) analysis of the brain cortex after induction and treatments. A) Control/Empty vehicle (note regular nuclear outline and normal mitochondrial distribution). B) Aluminum-induced case showing distorted nuclear contour. C) Subtle nuclear membrane irregularity and reformed nucleolus. D) Rare autophagic vacuole within a dystrophic neurite (arrow). C,D) Induced with aluminum then resveratrol treated cases both showing reduced autophagosome counts. TEM, original magnification X2000-X2500.

A-C) Aluminum-induced then curcumin-treated cases. A,B) Arrows pointing to few residual autophagic vacuoles (APVs) remaining after curcumin treatment. C) Aluminum-induced then curcumin-treated case showing a binucleated cell denoting regenerative activity. D) Aluminum-induced case co-treated with resveratrol and curcumin featuring budding “regenerating” nucleus and no evidence of APVs. TEM, X2000-2500.
Ultrastructural examination points to different mechanistic strategies underly the effects produced by both agents. Whereas resveratrol mainly targets autophagy (Fig. 1B-D), the electron microscopic hallmark of neurodegeneration was observed. On the other hand, curcumin was noted to augment regeneration (Fig. 2C). This notion suggests a synergistic action of both agents and explains why the best effects (Fig. 2D) where produced by a combination thereof.
Furthermore, chronic administration of Al induced biochemical modulations in the mid-brain cortexsections. Our results uncover significant increase in the lipid peroxidation and AChE activity as well as reduced glutathione reductase and catalase activities and reduced glutathione level as oxidative stress indices. Administration of the resveratrol-curcumin combination attenuated lipid peroxidation and ameliorated the biochemical changes significantly compared to resveratrol or curcumin alone(Table 2).
Profile of oxidative stress markers in mid-brain cortex tissue sections
Significance from the mock-treated at *p < 0.0001, **p < 0.001 or ***p < 0.01.
The curcumin and resveratrol combination regulates the expression of neurodegeneration-associated markers
Neurodegeneration-associated proteins including BACE1, AβPP, and γ-secretase subunits as well as inflammatory enzyme COX-2 expressions were also measured. As shown in Fig. 3, Al administration induced a highly significant increase (p < 0.0001) in the expression of BACE1 and AβPP as well as the inflammatory cytokines (IL-1β, IL-1α, and TNF-α) associated with a reduction in presenilins 1 and 2 (γ-secretase subunits). Moreover, we observed that treatment with either resveratrol or curcumin exerted a significant positive effect on the analyzed parameters compared to normal expressions. Interestingly, the resveratrol-curcumin combination induced an anti-inflammatory effect that resulted in the restoration of all measured parameter levelsback-to-normalization.

Neuroinflammation markers expression measured by semi-quantitative RT-PCR. Expression levels of neuroinflammatory markers BAC1, AβPP, and γ-secretase subunits (PS-1 and PS-2) were altered significantly in induced rats versus mock-treated (Mock-tr) ones. A) mRNA profile of the different parameters. Data indicated that curcumin treatment (Cur-tr) exerted positive significant effect in restoring the inflammatory markers levels (Il-1β, Il-1α, and TNF-α) close to normalization compared to resveratrol-treated (Rsv-tr) ones. Adding resveratrol to curcumin (Rsv & Cur-Tr) for treatment potentiated its effect significantly as indicated by the detected alterations in both inflammation-induced proteins and cytokines expressions versus mock-treated group. B) The relative expression levels as the ratio of each gene to β-actin calculated using Quantity One (ImageJ software). The p value calculated for each group compared to the control.
Previously we demonstrated that high APE1 expression is implicated in brain neuroprotection against both aluminum chloride and LPS-induced neurotoxicity [21, 22]. To test if a similar or different mechanism is involved in resveratrol-curcumin mediated therapeutic effect, we measured APE1 expression at both mRNA and protein levels under all experimental conditions. We observed that the significant Al-induced reduction (p < 0.01) of APE1 is counteracted by resveratrol or curcumin treatments (Fig. 4). The observed differential effect of both compounds on APE1 expression indicating that curcumin is more powerful as a reactive oxygen species scavenger, which is further confirmed by an in vitro study (Fig. 4). The detected alterations in the soluble Aβ40 protein levels indicated marked reduction in the induced versus mock-treated rats (Fig. 4). Treatments induced significantly the elevation in the intracellular level of Aβ40 as detected by western blotting analysis of total cell extracts prepared from brain cortex areas. Although the exact role of Aβ40 in the central nervous system is not well understood, some reports highlighted that Aβ40 exerts antioxidant activity against stress and regulates the functions of some transcription factors. Consistently herein, we postulate that under therapeutic condition and tissue recovery from chronic inflammation, an elevated Aβ40 might play an essential role in counteracting oxidative stress and in genetic remodeling. Curcumin has been recently reported as a selective inhibitor for COX-2 expression; therefore, we tested the possible implication of this activity in our model. COX-2 expression, as a pro-inflammatory enzyme, elevated significantly (p < 0.0001) after Al induction by around 4.5 folds, and resveratrol treatment did not reduce it significantly (its expression level is around ∼4 fold). Although treating induced rats with curcumin alone reduced COX-2 expression by around 2.5 times, the resveratrol-curcumin combination exerted more potent synergistic effect by reducing its mRNA and protein levels to around1.5-fold (Fig. 4).
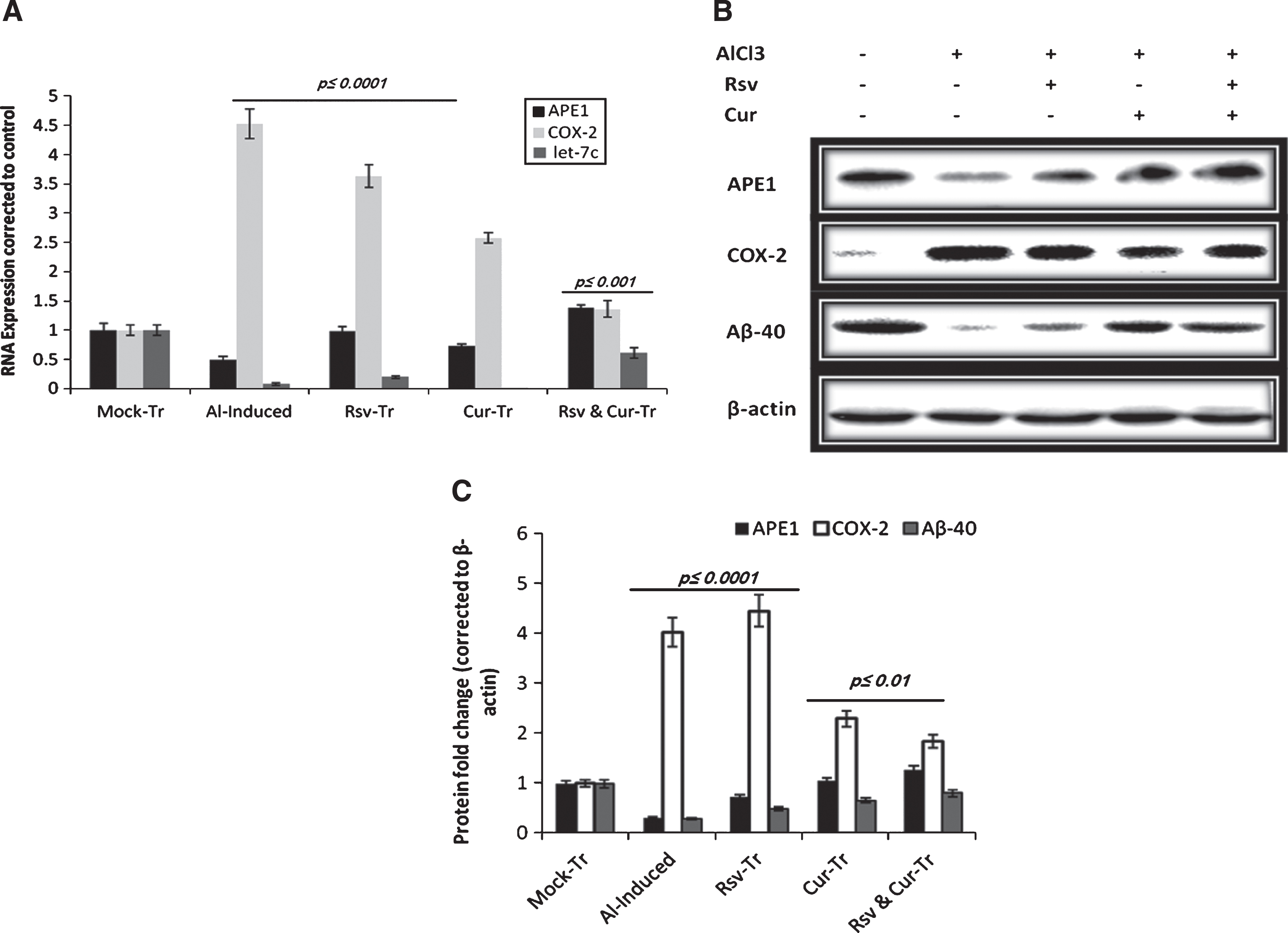
In vivo Let-7c, APE1, and COX-2 RNA levels as well as the western blotting analysis of some neuroinflammation associated proteins. A) We detected significant reduction in let-7c microRNA expression by Al induction. Although treatment with either resveratrol (Rsv) or curcumin (Cur) alone induced let-7c expression significantly, their combination (Rsv & Cur-Tr) exerted more significant effect on let-7c induction. B) Development of neurodegeneration-like state was further confirmed by the detection of tissue Aβ40 soluble protein levels in induced versus treated rats. Moreover, inflammatory enzymes APE1 and COX-2 levels were significantly modulated by aluminum chloride stimulation. We observed significant reduction in APE1 (A) mRNA and (B) protein levels; on the contrary, COX-2 expression was significantly induced. C) Western blotting quantification is calculated as a relative expression of each group from control.
MicroRNA Let-7c, which is highly expressed and plays an essential role in brain development, was significantly decreased (∼50%) in the cortex of induced rats (Fig. 4). Resveratrol treatment induced significant let-7c expression while curcumin did not, which indicate the different mechanistic actions of the two compounds. Furthermore, treating induced rats with the resveratrol-curcumin combination for four weeks induced back-to-normal restoration of let-7c expression as shown in Fig. 4.
Addition of resveratrol, curcumin, or their combination protects PC-12 from aluminum toxicity
In a follow-up pattern, we measured the expressions of APE1, COX-2, and let-7c as major molecular targets at early and late stages in the response to Al-induced toxicity in cultured PC-12. One of the important points to uncover was the differential effect of resveratrol and curcumin, alone or in combination. PC-12 cells were treated with Al at the concentration of 0.5 mM and cells were collected at two time intervals of 6 and 24 h (several pilot experiments were run to standardize Al dose and follow-up time).
Consistent with the in vivo data, we found that resveratrol alone exerted protective effect by significantly inducing APE1 and maintaining high level of let-7c expression both at 6 and 24 h (Figs. 5 and 6), while curcumin, which acts as a potent free radical scavenger, reduced both APE1 and COX-2 expressions significantly (p < 0.0001) compared to Al- and resveratrol-treated cells at 6 h post stimulation (Figs. 5 and 6). The significant toxicity of Al and its consequent effect on the inflammatory genes expression, especially on COX-2 level, was reduced by around 50% upon curcumin co-addition. The combination of resveratrol-curcumin significantly inhibited COX-2 and induced APE1 and let-7c levels compared to induced rats (Figs. 5 and 6).
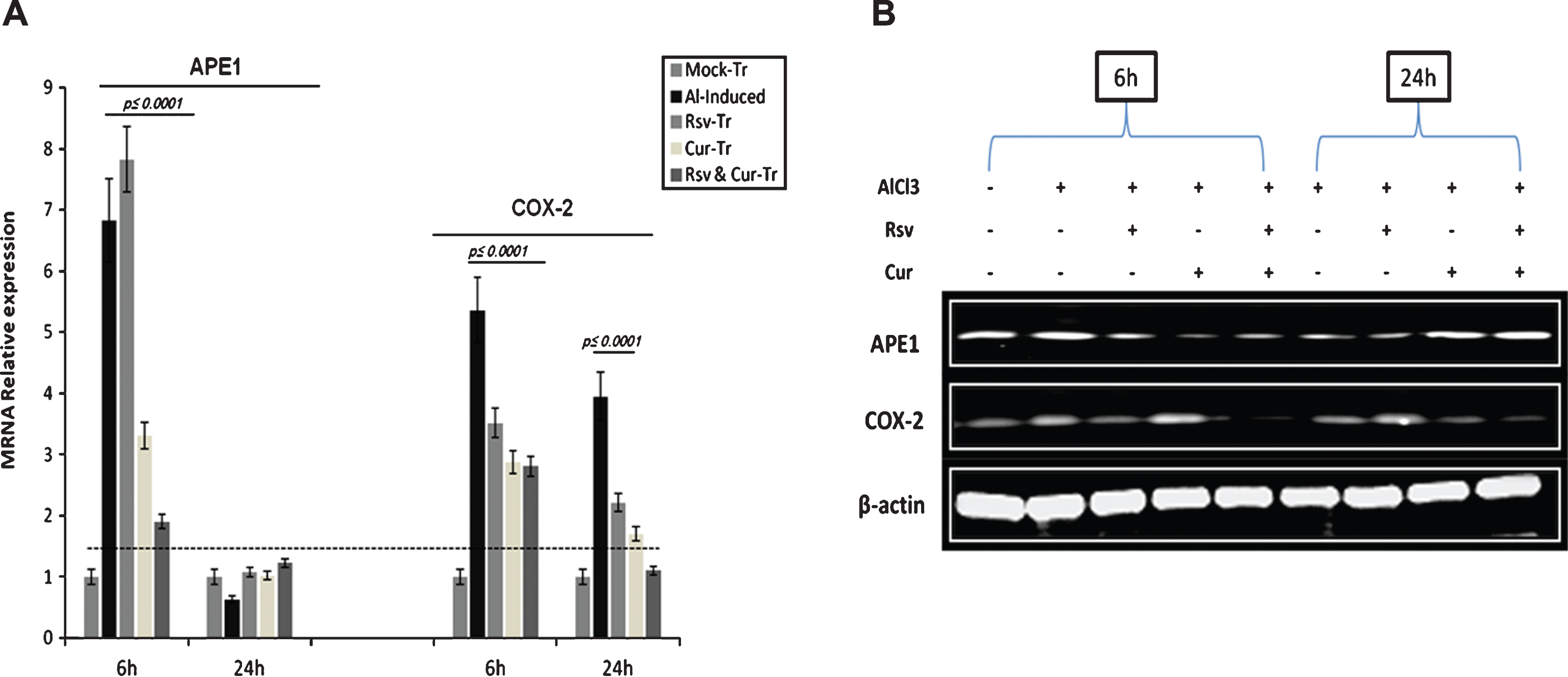
QRT-PCR and western blotting analysis of APE1 and COX-2 expressions in PC-12 cells treated with Al. A) mRNA expression levels of APE1 and COX-2 under the different experimental conditions. The results indicated that resveratrol (Rsv) and curcumin (Cur) act differentially to counteract Al-induced stress, with profound antioxidant effect in curcumin-treated (Cur-Tr) cells. B) In parallel to mRNA levels, western blotting analysis indicated that curcumin co-treatment induced marked reduction in COX-2 level, while resveratrol co-treatment induced significant elevation in APE1 expression at 6 h as part of the cells initial response to Al-induced stress.
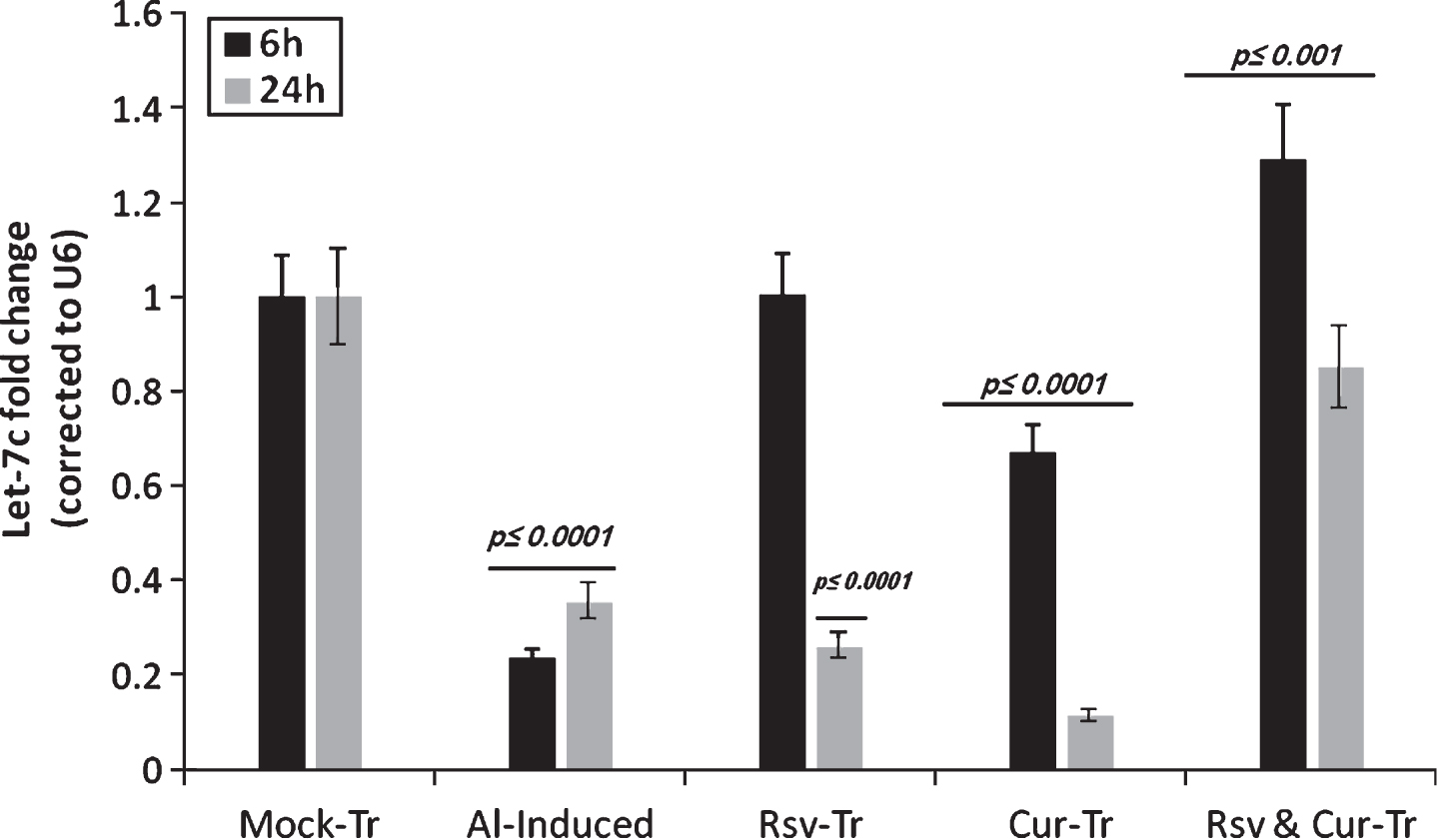
Let-7c expression profile in PC-12 cells induced by Al at 6 and 24 h, respectively. The expression of the miRNA let-7c after 6 and 24 h of Al stimulation indicated significant reduction compared to mock-treated (Mock-Tr) cells. On the other hand, co-addition of resveratrol (Rsv) attenuated Al-induced effect, in part, as indicated by maintaining normal expression of let-7c at 6 h. At 24-h of Al stimulation, let-7c level was reduced significantly upon Al stimulation alone or under the co-addition of either resveratrol or curcumin. On the contrary, PC-12 co treatment with both resveratrol and curcumin (Rsv & Cur-Tr) after Al stimulation showed normal let-7c expression compared to mock-treated.
DISCUSSION
The role of environmental pollutants such as toxic metals, pesticides, and phenols in aging, neurodegenerative disorders, and neuronal apoptosis is not conclusive. Al is one of the most copious earth elements that play an active role in the pathogenesis mechanisms of neurological disorders [38, 39]. The development of neurodegenerative disorders is among the most common problems arising from the chronic exposure to Al. Administered Al was reported to predominantly accumulate in different brain regions including hippocampus and cortex [39, 40]. Furthermore, recent novel studies have indicated that aluminum chloride induces loss of spine density in hippocampal CA1 and dentate gyrus regions [41].
In the current study, we decided to investigate the controversial role of Al with respect to its relation with AD and neuroinflammation disorders using both in vivo and in vitro models.
Our results disclosed that Al induces cytotoxicity over time as shown by abnormal alterations in the ultrastructure of brain cortex and assessed by transmission electron microscopic (TEM). Neuronal loss and accumulation of autophagic vacuoles were observed in Al-induced brain sections analyzed by TEM. Four weeks of rat treatment with either resveratrol or curcumin induced partial tissue recovery compared to the mock group. Interestingly, adding resveratrol to curcumin induced marked tissue recovery as it appears by TEM analysis.
Al toxicity to the neurons is mediated mainly by increasing cellular oxidative stress. Rats were orally administered total amount of 3.325 g/kg aluminum chloride over four weeks. We assessed oxidative stress-related biochemical markers including sera total antioxidant capacity, tissue MDA/GSH contents, and SOD/catalase/GST activities. Consistent with previous reports [21, 42–45], our results indicated significant reduction in the tissue antioxidant capacity in association with progressive neuroinflammation.
Furthermore, we detected significant increase in AChE activity at the end of induction period in parallel to what has been previously shown [46]. AChE is one of the brain’s highly active enzymes that is widely distributed and has several functions in cholinergic and neuromuscular synapses. One of the reported non-classical roles of AChE is its ability to accelerate the assembly of the Aβ peptide into amyloid fibrils [47–49] hence AChE/Aβ complexes showed enhanced neurotoxicity compared with fibrils containing only Aβ [50]. This could explain our detected significant elevation of cortex AChE activity that may, in part, contribute to cellular neurotoxicity and enhanced cell death. Moreover, we investigated the possible synergistic effect of adding resveratrol to curcumin for treatment of aluminum chloride-induced neurodegeneration on male rats. Natural products exert protective effects mainly through free radical scavenging mechanisms. Resveratrol and curcumin are well studied as antioxidant compounds in different experimental models. However, many reports have discussed the neuroprotective effect of curcumin against brain-induced toxicity [22, 52] and others illustrated the defensive role of resveratrol in induced neuroinflammation and neurodegeneration models [21, 53–55], but there is no documented work on the effect of their combination. Furthermore, although treatment of induced rats with either resveratrol or curcumin alone inhibited partially Al-induced elevation in AChE activity, which is in agreement with what was previously published [56], their combination significantly restored its activity in the cortex tissue, indicating their synergistic effect.
Protein misfolding and aggregations are considered key players in the pathogenesis of neurodegenerations. Abnormal expression of AβPP and its processing enzymes β- and γ-secretases occur as part of Al-induced chronic neuroinflammation that is triggered by glial cells activation [21, 58].
Herein we determined the neurodegeneration-associated indices including AβPP and γ-secretase as well as the activated glial cells produced inflammatory cytokines (IL-1β, IL-1α, and TNF-α) expressions. As shown, Al stimulation for four weeks induced significant increase in the expressions of BACE1 and AβPP as well as inflammatory cytokines associated with a reduction in presenilins 1 and 2 (γ-secretase subunits). Treating induced rats with either resveratrol or curcumin alone recovered the expression of all parameters by around 20% compared to induced group.
Interestingly, the resveratrol-curcumin combination was effective as an anti-inflammatory formula that resulted in restoration of all measured parameter levels almost back-to-normalization. Convergent with previous studies [21, 58], we postulatethat resveratrol-curcumin combination acts in arapport that is mediated by the interference in different cellular mechanisms independently and/or synergistically.
APE1, a key enzyme in base excision repair pathway, is reported to play an essential role in the mechanism of cell survival versus death upon stimulation with cytotoxic agents. Therefore, it has emerged as an attractive therapeutic target for many diseases including neurodegenerative disorders [reviewed in 59].
Consistent with our previous work [21, 22], treatment of the induced rats with either resveratrol or curcumin alone partially attenuated the induced brain damage. The detected alterations in the soluble Aβ40 levels during induction and treatment may attribute not only to the regulation of gene expressions but also to the association between APE1 and different intracellular proteins that play crucial role in Aβ-mediated signaling pathway [60].
Curcumin is the principal curcuminoid of the spice turmeric (Curcuma longa), a member of the ginger family [61]. Besides having a number of pharmacological properties [62], it exerts anti-inflammatory activity by inhibiting a number of different molecules that participate in the process of inflammation [63]. At the molecular level, curcumin is a toxic natural inhibitor of NF-κB and as a result modulates the expression of various genes such as cyclooxygenase-2, matrix metalloproteinase-9, inducible nitric oxide synthase, interleukin-8, and anti-apoptotic proteins, which are regulated by NF-κB [64]. The resveratrol-curcumin combination significantly attenuated aluminum chloride-induced oxidative damage; ‘This antioxidant property of curcumin has been attributed to the presence of chain breaking or hydrogen donating phenolic groups in its molecular structure’ [61].
A major question of our study was to address the possible involvement of the small regulatory elements, miRNA modulation in the pathogenesis and treatment of our established in vivo and in vitro models. miRNAs have been recently reported to be implicated in neurodegenerative disorders including AD [65]. Let-7c belongs to one of the important miRNA families, let-7, that are highly expressed and play an essential role in brain development[66].
We have previously showed that let-7c is one of the modulated targets in LPS-induced brain damage in rats and it is one of the miRNAs that are implicated in curcumin-mediated brain protection [22]. In harmony with our previous report, we detected a significantly reduced expression of Let-7c in the cortex of induced rats. Treatment with resveratrol alone induced let-7c expression by around 20% of the mock-treated rats, while curcumin did not. In parallel, the resveratrol-curcumin combination for four weeks induced significant re-elevation in let-7cexpression.
Because until now, there has been a lack of a suitable in vitro model for neural cell replication, and there is a limitation in the available number of approved therapeutic agents that help in treatment or recovery of the injured nerve functions. Therefore, we decided to further elucidate the mechanism by which the proposed compounds exert their therapeutic effect by conducting an in vitro study using PC-12 cells. PC-12 cells have an embryonic origin from neural crest of the rat adrenal medulla and represent an established model for in vitro study of neurological disorders.
We decided to test the alterations in the inflammatory enzymes APE1 and COX-2 in addition to let-7c miRNA as targets for our natural combination. Because cultured cells are rapidly growing and lack the need of the in vivo system complicated networking to operate, we ran several pilot experiments to optimize the proper time and concentration of the Al (data not shown). An inflammation-like state was established in PC-12 using Al treatment at different time intervals; 6 h (to mimic the initial defensive response) and 24 h (that represent the chronicstate).
Stimulation of PC-12 with Al triggered a quick and strong APE1 expression at 6 h. Co-addition of curcumin to Al restrained APE1 induction by around 50% while resveratrol induced more APE1 expression. In parallel to our findings; the sudden induction of APE1, as a redox sensor, in response to genotoxic stimulation is well documented before reviewed in [58]. One of the important points to uncover was the differential effect of either resveratrol or curcumin alone in addition to their combination on the expression levels of APE1, COX-2, and let-7c. Cells co-treated with Al and the resveratrol-curcumin combination had higher APE1 expression at 24 h compared to Al-stimulated cells. Both resveratrol and curcumin protected PC-12 at late time by maintaining high APE1 level. COX-2 expression is observed to be significantly reduced at 24 h by all treatment modalities compared to Al-stimulated cells. In addition, we observed that curcumin exerted more inhibitory effect on COX-2 than resveratrol, which is compatible with what has been previously reported [67]. Interestingly, adding resveratrol to curcumin potentiated its effect on COX-2 inhibition, possibly through indirectmechanisms.
In parallel to the in vivo results, the let-7c role as a protective miRNA is confirmed. Al induction reduced intracellular let-7c levels by around 80% and 65% compared to the mock-treated cells at 6 and 24 h, respectively, an effect that is countered by co-addition of either resveratrol or curcumin to Al-stimulated cells. Obviously, our results indicate that resveratrol acts, in part, via induction of let-7c miRNA as shown by maintaining normal let-7c level at 6 h while curcumin did not. The positive synergistic effect of adding resveratrol to curcumin is further confirmed by the detected high expression of let-7c at both 6 and 24 h. Controversial explanations are proposed to clarify the exact mechanism by which miRNA are involved in the pathology and recovery conditions.
AβPP-binding family B, member 3 (APBB3) is a cytoplasmic protein that is reported to bind to AβPP as well as to other AβPP-like proteins, and hence play a role in AD pathology [68]. APBB3 (NM_053957) is one of the let-7c miRNA targets in the database of microRNA (http://www.microRNA.org) –Targets and Expression with calculated mirSVR score of –0.1324 and PhastCons score of 0.6695. Therefore, we suggest a positive correlation between let-7c inductions, increased Aβ40 soluble protein, and reduced AβPP expression in treated rats’ cortex tissues in accordance with let-7c role as classical suppressor of APBB3 expression.
We introduce let-7c as a novel target for the action of resveratrol. The effect of resveratrol on let-7c expression might be attributed to different mechanisms including but not limited to the effect of resveratrol as a pharmacological compound that alters different intracellular transcription factors functions and/or activities (Fig. 7).

Illustration of the differential effects of resveratrol (Rsv) and curcumin (Cur) on the molecular targets in injured brain tissues.
Conclusion
Herein we show that oxidative stress, genetic/epigenetic alterations, and neuronal loss are implicated in Al-mediated neuronal toxicity. Based on the results emerging from cell culture and animal studies, our findings clearly indicate that both resveratrol and curcumin are exceptional candidates as therapies against neuroinflammation and other related degenerative disorders, ready to enter clinical trials within a short time frame.