
Abstract
Background:
Animal models of dementia associated with metabolic abnormalities play an important role in understanding the bidirectional relationships between these pathologies. Rodent strains develop cognitive dysfunctions without alteration of peripheral metabolism following intracerebroventricular administration of streptozotocin (icv-STZ).
Objective:
We aimed to estimate the effect of icv-STZ on cognitive functions and peripheral metabolism in Lewis rats, which are rarely used for the induction of cognitive abnormalities.
Methods:
Inbred adult Lewis rats were treated with single icv-STZ (3 mg/kg). Cognitive functions were assessed using Morris water maze (MWM) test and locomotion by the Open Field test. Metabolic alterations were studied using histological and biochemical analysis of brain and peripheral tissues.
Results:
The icv-STZ induced rapid weight decline during the first two weeks. Thereafter, the rats showed an accelerated weight gain. Three months after the icv-STZ treatment, the rats were severely obese and revealed fatty liver, pancreatic islet hypertrophy, significantly elevated levels of blood insulin, leptin, and adiponectin, but intact peripheral glucose homeostasis. The icv-STZ rats expressed amyloid-β deposits in blood vessels of leptomeningeal area, microgliosis, astrogliosis, and spongiosis in fimbria-fornix area of hippocampus. Locomotor activities of icv-STZ treated and sham-operated rats were similar. In the MWM test, the icv-STZ treated rats demonstrated severely impaired spatial learning during both acquisition and reversal phases.
Conclusions:
Icv-STZ treated Lewis rats develop severe dementia associated with obesity and peripheral metabolic abnormalities. This animal model may be useful for exploring the pathophysiological relationship between obesity and dementia and provides a new tool for development of effective therapy.
Keywords
INTRODUCTION
The increased prevalence of dementia associated with obesity-related metabolic dysfunctions is a growing public health concern. However, the relationship between dementia and obesity is still controversial. Several studies have found that being overweight or obese during middle age increases the risk of dementia in later life [1, 2]. On the other hand, a retrospective cohort study has found that obese people actually have a lower risk of dementia than those with a healthy weight, and being underweight in middle and old age apparently increases the risk of dementia [3].
Until now, only few studies have examined the association between body mass index (BMI) changes in patients with Alzheimer’s disease (AD) and the rate of clinical progression of dementia. Approximately 30–40% of patients with mild to moderate AD lose weight after disease diagnosis [4, 5]. Moreover, accelerated weight loss may precede diagnosis in dementia [6, 7]. In patients with amnestic mild cognitive impairment, weight loss was associated with faster progression of the disease [8]. Interestingly, in those cases where BMI declines before the clinical AD onset, it levels off after clinical AD onset, and might even increase in advanced stage of the disease [9]. So far, the biological mechanisms of such associations between weight changes and dementia are poorly understood. Proposed mechanisms for weight changes before and after diagnosis of dementia include medial temporal cortex atrophy, biological disturbances and changes in eating behaviors; however, none has been proven [10, 11]. Recent evidence indicates that several hormones such as insulin, leptin, and adiponectin regulate brain functions responsible for feeding behavior, energy stores, and various aspects of memory and cognition [12–14].
Animal models of obesity and dementia play an important role in understanding pathogenesis of these disorders and development of an effective therapy. Behavioral and cognitive dysfunctions were found in several genetic or non-genetic models of obesity in mice and rats [15, 16]. In most genetic models of AD, obesity was induced by various high-calorie diets. These studies show a correlation between weight reduction, decreased cerebral amyloid/tau pathology and improved cognition [13, 18]. However, genetic AD models resemble the rare familial forms of the disease, while the majority (more than 95%) of the AD dementia cases belong to the sporadic form in which both genetic predisposition and environmental factors contribute to the genesis of the disease [19]. A commonly used animal model of sporadic AD was generated using intracerebroventricular administration of streptozotocin (icv-STZ) in Wistar and Sprague-Dawley rat strains [20, 21]. STZ, a glucosamine-nitrosourea compound, is selectively toxic toward insulin-producing pancreatic beta cells due to its cellular uptake by the low-affinity glucose transporter 2 (GLUT2) protein densely expressed in beta cell membranes [22]. It has been hypothesized that chronic human exposure to nitrosamine compounds, which are widely present in processed foods, contributes to the pathogenesis of diabetes mellitus and AD-type neurodegeneration [23]. A high dose of peripherally injected STZ (70–200 mg/kg) induces diabetes in rodents due to death of pancreatic islet beta cells [24], while icv-STZ in low subdiabetogenic doses (1–3 mg/kg) does not alter peripheral glucose homeostasis, but gradually decreases cerebral glucose uptake, impairs brain insulin signaling, and produces multiple other effects that resemble molecular, pathological, and behavioral features of AD. STZ cytotoxicity to the brain may be explained as the result of its uptake by GLUT2-expressing cells widely distributed in the brain, in particular, in the circumventricular organs and hypothalamus [25–27]. Nevertheless, the precise mechanisms of icv-STZ activity in the brain are still poorly understood.
The icv-STZ induces AD-like cognitive dysfunctions in rats [21, 27], mice [28], and non-human primates [29]. However, obesity-related peripheral metabolic dysfunctions associated with cognitive decline were not previously described in any experimental animals exposed to icv-STZ. In the present study, we aimed to estimate the effects of single icv-STZ on cognitive functions and peripheral metabolism in inbred Lewis rats, which are rarely used for the induction of cognitive abnormalities.
MATERIALS AND METHODS
Animals and experimental groups
The experiments were approved by the Institutional Animal Care and Use Committee. 10–12-week-old male Lewis (LEW/SsNHsd) inbred rats (Envigo RMS Ltd, Rehovot, Israel) weighing 260–300 g were used in all experiments. The rats were maintained on a regular 12-h dark/light cycle with access to food and water ad libitum. Standard chow diet was from Envigo RMS Ltd. The animals were divided into three groups, as follows: 1) rats exposed to icv-STZ (icv-STZ rats); 2) sham-operated rats exposed to icv administration of vehicle (artificial cerebrospinal fluid, aCSF); and 3) intact rats. The total number of animals per group and time points are presented in the figure legends. An overview of the study design is illustrated in Fig. 1.

Study design. Icv-STZ, intracerebroventricular administration of streptozotocin; MWM, Morris water maze test; OF, Open Field test.
Icv administration
Rats were anesthetized by an intraperitoneal injection of ketamine/xylazine mixture (100 mg and 15 mg per kg body weight, respectively) 10 min before surgery. STZ (3 mg/kg body weight) dissolved in 8 μl of aCSF (120 mM NaCl, 3 mM KCl, 1.15 mM CaCl2, 0.8 mM MgCl2, 0.33 mM NaH2PO4, 27 mM NaHCO3, pH 7.2) or 8 μl of aCSF alone was bilaterally icv-injected (4 μl/2 min in each side) using stereotactic apparatus (Stoelting Co., Wood Dale, IL) and microdrill (RWD Life Science, San Diego, CA). To prevent decomposition, the STZ solution was freshly prepared for each rat just before the injection. The stereotactic coordinates for icv injection were: 0.9 mm posterior, 1.8 mm lateral, and 3.8 mm ventral from the bregma level [30].
Body weight, blood glucose level, and glucose tolerance
Body weight changes were recorded for the duration of the experiment (0, 3, 9, 12, 14, 16, 22, 30, 60, and 90 days) as indicated in Fig. 2A. Non-fasting blood glucose concentrations were measured with a portable glucometer (Accu-Check; Hoffmann La Roche, Basel, Switzerland) in whole blood samples collected from the tail between 9 a.m. and 10 a.m. An intraperitoneal test tolerance to glucose (1 g glucose/kg) was performed in all experimental groups. Glucose solution was injected intraperitoneally after 6 h fasting. Blood glucose was measured before and at 15, 30, 60, and 120 min after glucose injection.
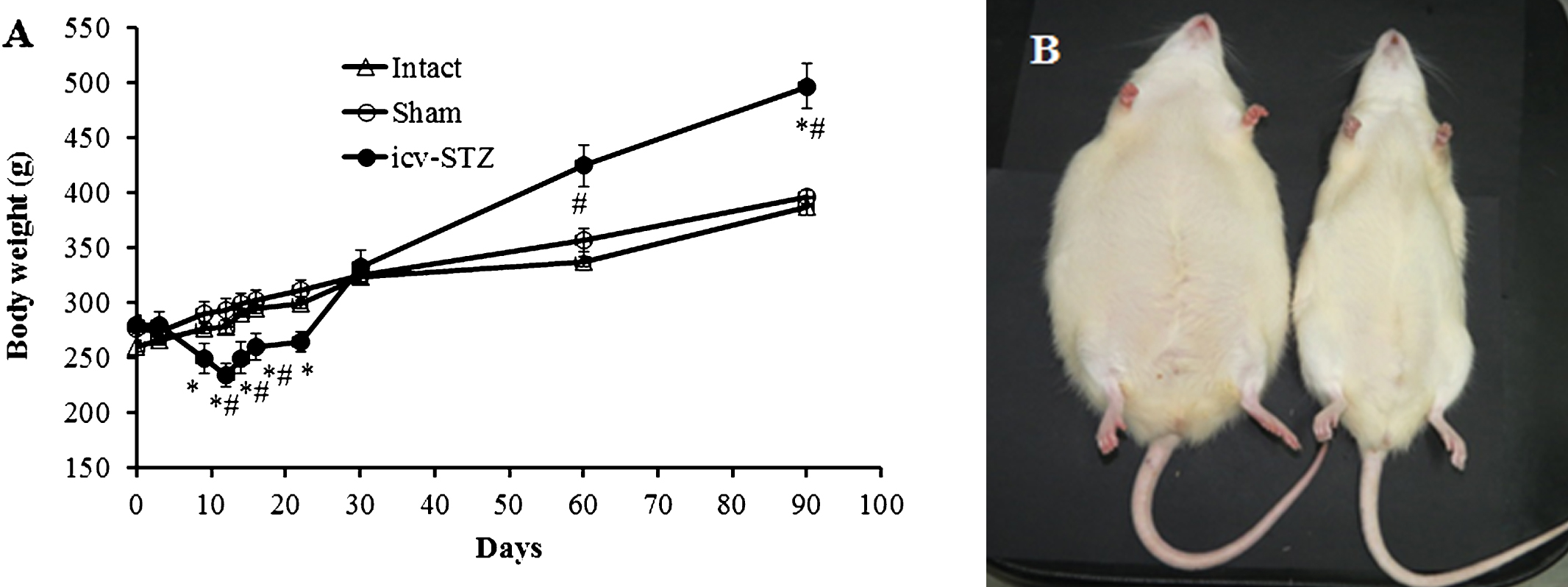
Body weight changes. A) Icv-STZ induced accelerated body weight decline lasting about two weeks and then sharply shifted to intensive weight gain resulting in development of visual symptoms of abdominal obesity in the Lewis rats (n = 12). Sham-operated (n = 13) and intact rats (n = 12) demonstrated moderate body weight gain without signs of abdominal obesity. *p < 0.001 compared to sham group, #p < 0.05 compared to intact group. Data presented as average±SE. B) Representative image of obese icv-STZ rat and lean sham-operated rat three months after icv-administration.
Blood level of insulin, leptin, and adiponectin
Three months after the start of the experiments, the concentrations of insulin, leptin, and adiponectin were measured in the blood serum of intact animals and rats exposed to icv-STZ or icv-vehicle. The serum level of leptin and total adiponectin were measured using ELISA kits (R&D systems, Minneapolis, USA). The serum insulin level was measured using an ELISA kit for rat insulin according to the manufacturer’s protocol (Mercodia AB, Uppsala, Sweden). Blood samples, collected from the orbital sinus of the anesthetized non-fasting rats, were allowed to clot for 2 h at room temperature before centrifuging for 20 min at 1500×g. Aliquots of serum were stored at –35°C.
Visual observations
Visual observations of animal behavior were carried out between 9 a.m. and 10 a.m., three times per week by the same investigator for the duration of the study.
Open field (OF) test
The OF test is one of the most established procedures to evaluate animal mobility and anxiety [31]. 10 weeks after starting the experiments, the animals were exposed to OF test. Briefly, an individual rat was placed in an unfamiliar square open field (65×65 cm) with 40 cm high walls, the floor of which had been divided into two areas: the peripheral zone (10 cm from each wall of the arena) and the center zone. The animal’s behavior was videotaped for 20 min and off-line analyzed using an automated tracking system (Ethovision 3.1 Noldus Information Technology B.V., Wageningen, The Netherlands).
Morris water maze (MWM) test
Spatial learning and memory were measured using MWM as described [32]. This test assesses the ability of the animals to locate an underwater hidden platform, using surrounding visual cues. The water maze is a circular pool (diameter –1.8 m; height –60 cm) filled with water (21±1°C). A circular platform (10 cm diameter) was placed 1 cm under the water surface. To find the platform (the acquisition phase), rats performed four trials/day on three consecutive days (up to 120 s for each trial). The MWM tests were performed 6 weeks (first MWM test) and 12 weeks (second MWM test) after starting the experiments. After 12 weeks in addition to the acquisition phase, on the 4th and 5th days the platform was moved to another quarter of the maze (the reversal phase). The time taken to reach the platform was recorded. If a rat did not find the platform within 120 s, it was manually placed on it for 20 s. In addition, motility parameters including velocity and motility time were analyzed. Data were recorded using an automated tracking system (Ethovision 3.1 Noldus Information Technology B.V., Wageningen, The Netherlands).
Histology and immunohistochemistry of pancreas, liver and brain
Three months after icv injections, the abdominal cavity of the rats was macroscopically examined. Whole rat fixation procedure was made using 4% paraformaldehyde perfused via the vascular system. Liver, pancreas, and brain were excised, processed routinely for paraffin sectioning (4 μm thick) and stained with hematoxylin-eosin (H&E). In addition, pancreas was stained with Masson’s Trichrome.
Immunohistochemical analysis (IHC) of pancreatic islets for insulin and glucagon was performed as described previously [33]. Briefly, sections were blocked with 20% normal serum and incubated with primary polyclonal guinea pig anti-insulin antibodies (1 : 1000) and monoclonal mouse anti-glucagon antibodies (1 : 2000) overnight at 4°C. After washing, sections were incubated with secondary antibodies: Cytm3- or Cytm2-conjugated affinipure anti-guinea pig Ig (1 : 400) and Cytm2-anti-mouse Ig (1 : 400) for 1 h at room temperature. All dilutions were prepared with Dako antibody diluent. Nuclei were counterstained with DAPI (1 : 200 in PBS) for 5 min. The microscopic images were captured by a digital camera connected to a fluorescent microscope (BX-52, Olympus Ltd, Japan). Immunostaining of insulin and glucagon was negative when the primary antibodies were replaced with Dako antibody diluent or normal serum. Sections of pancreas stained with Masson’s trichrome were evaluated under a light microscope.
Sagittal sections of the brain were used for IHC analysis. Briefly, tissue pre-treatment to unmask immunoreactive sites for amyloid-β (Aβ) antigens was conducted using incubation with 98% formic acid for 7 min at room temperature. Sagittal brain sections of 5XFAD transgenic mice (males, eight months old) were used as a positive control for Aβ deposition in the brain. Heat-induced epitope retrieval was used for detection of GFAP. PeroxFree Block (Cell Marque, Rocklin, USA) was used to block the endogenous peroxidase enzymes. The brain sections were incubated overnight at 4°C with mouse monoclonal Aβ antibodies (1 : 300) from Novusbio, polyclonal rabbit Iba1 antibodies (1 : 500) from Wako or GFAP antibodies (1 : 1000) from Dako. Staining was detected using mouse/rabbit PolyScan HRP/DAB detection system (Cell Marque, Rocklin, USA), according to the manufacturer’s protocol.
Quantification of positive staining was completed using ImageJ/Fiji software. Briefly, color images were split into single color channels and each channel was converted to grayscale. The grayscale image was then converted to a binary image by adjusting the image threshold to highlight only the area occupied by the positive immunostaining. Once the image was in binary, percent of positive immunostaining over total image size was analyzed. Three rats (seven sections per each rat) were analyzed for each experimental group.
Statistical analysis
Data were analyzed using Statistical Package for the Social Sciences (SPSS) 17.0 software (SPSS Inc., Chicago, IL). Comparisons between experimental groups for body weight and glucose tolerance were performed using two-way repeated measure analysis of variance (RM-ANOVA) and the Bonferroni post hoc test. Alterations in the serum levels of hormones and histological assessment of neuroinflammation were analyzed by one way analysis of variance (ANOVA) and the Tukey post hoc test. Comparisons between experimental groups in the OF test were performed using one way ANOVA and the Tukey post hoc test. The results were expressed as mean±SE and considered significant at p < 0.05. The MWM experiments (escape latency) were evaluated using a Multivariable Cox regression time to event approach in SAS® software. A RM-ANOVA was not considered appropriate for these data as they were not normally distributed and the data were censored at 0 and 120 s [34]. The event was defined as finding the platform and the time to event was defined as the time in seconds from placing the animal in the tank to the animal finding the platform. This approach allowed us to investigate the interactions between different treatments and the days of the test. The acquisition phase and the reversal phase were analyzed separately. MWM motility parameters were analyzed using two ways RM-ANOVA and the Bonferroni post hoc test.
RESULTS
Weight changes
Data presented in Fig. 2A show body weight changes in icv-STZ rats, sham-operated, and intact rats. A two ways RM-ANOVA test for body weight found significant differences between the groups (F (2,19) = 9.83; p < 0.0001), the time (days) (F (9,19) = 99.12; p < 0.0001), and an interaction (F (2,29) = 17.11; p < 0.0001). Bonferroni post hoc test showed that icv-STZ group differ from sham group at days 9, 12, 14, 16, 22, and 90 (p < 0.001) and from the intact group at days 12, 14, 16, 60, and 90 (p < 0.05). No significant differences were found between sham and intact groups. During the first two postoperative weeks, the body weight of icv-STZ rats was significantly lower compared to intact and sham-operated rats. Thereafter, the accelerated weight decline was sharply shifted to intensive body weight gain, resulting in development of visual symptoms of abdominal obesity in icv-STZ rats (Fig. 2B). Three months after icv-STZ, some animals weighted more than 620 g. Sham-operated and intact rats did not show significant weight reduction and demonstrated only moderate body weight gain without signs of abdominal obesity. Maximal body weight of sham-operated and intact rats did not exceed 440 g.
Blood glucose
Non-fasting blood glucose levels in rats from all groups were in physiologically normal range (90–120 mg/dl) during all the experimental period. Intraperitoneal test tolerance to glucose (IPTTG) was performed at the end of experiment. Two-way RM-ANOVA for IPTTG showed no significant differences between the groups (F (2,34) = 1.22; p = 0.30), but significant interaction (F (8,34) = 6.35 ; p < 0.0001) was revealed (Fig. 3A). Bonferroni post hoc test showed differences in level of glucose (15 min after glucose injection) between icv-STZ rats and rats from both intact and sham groups (p < 0.001). At this time point, the level of glucose was decreased in icv-STZ rats. No significant differences were found between the sham and intact groups.
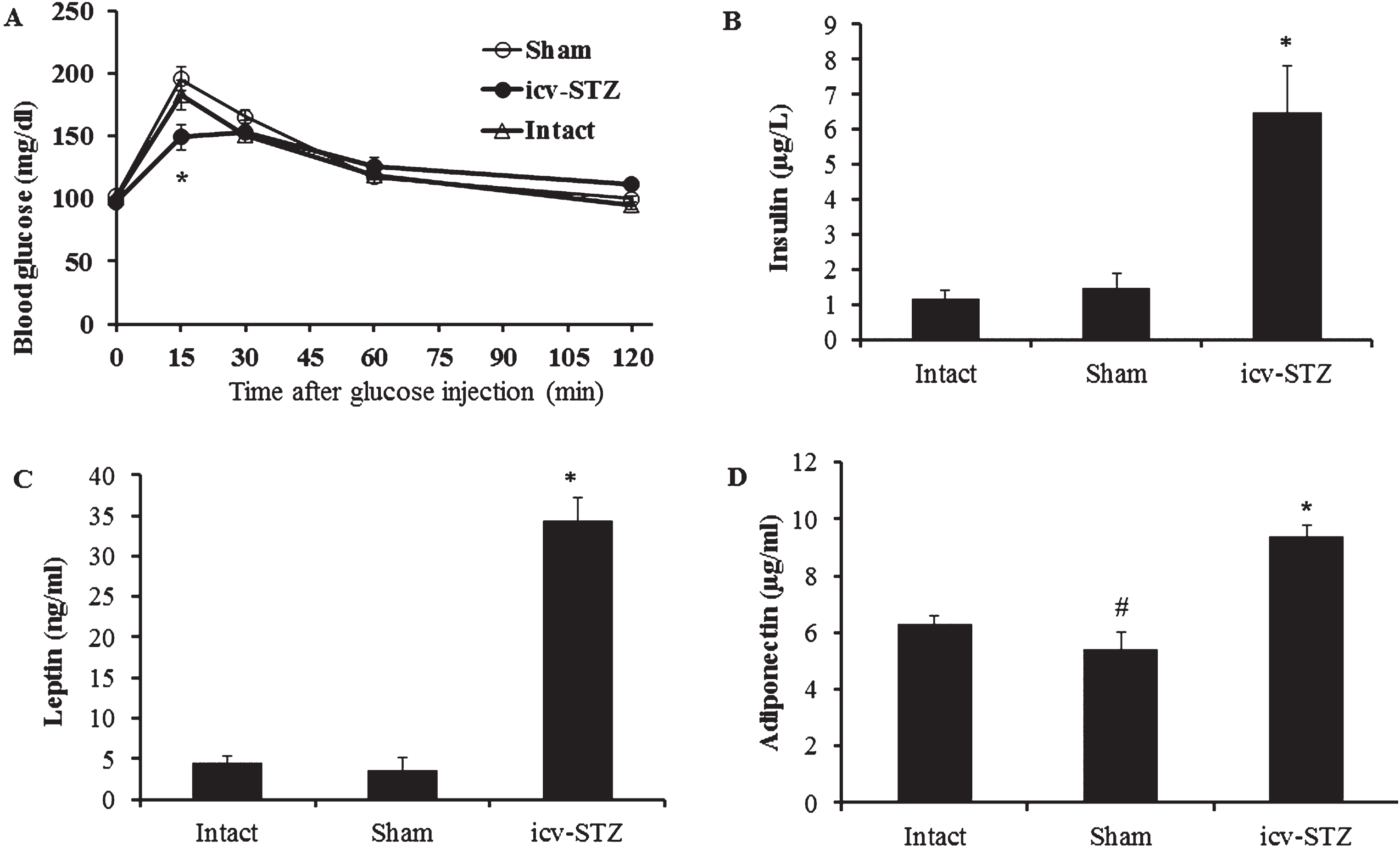
Blood glucose, insulin, leptin and adiponectin. A) IPTTG (1 g glucose/kg): 25% dextrose solution was injected intraperitoneally after 6-h fasting at a dose of 4 μl/g body weight. Blood glucose was measured before and after glucose injection. No significant differences in IPTTG were found between intact (n = 12), sham-operated (n = 13) and icv-STZ Lewis rats (n = 12). Blood levels of insulin (B), leptin (C), and adiponectin (D) detected using ELISA kits were statistically significantly increased in icv-STZ rats (n = 12) compared to sham-operated (n = 13) and intact (n = 13) rats. IPTTG and collection of blood samples for measurements of hormones were carried out three months after beginning the experiments. *p < 0.001 compared to sham and intact group, #p < 0.05 compared to intact group. Data presented as average±SE.
Hormones (insulin, leptin, and adiponectin)
Concentrations of insulin, leptin and adiponectin in blood serum of rats were tested three months after starting the experiments. One-way ANOVA for serum insulin and leptin levels demonstrated significant differences between the groups (F (2,33) = 17.23; p < 0.0001 and F (2,34) = 114.68; p < 0.0001, respectively). Tukey post hoc tests showed no significant differences in serum levels of insulin and leptin between the sham and intact groups. However, icv-STZ rats differed significantly from the other groups (p < 0.001), expressing increased level of these hormones compared to both intact and sham-operated rats (Fig. 3B, C). A one-way ANOVA for serum adiponectin found a significant difference between the groups (F (2,34) = 38.15; p < 0.0001). Tukey post hoc test showed differences between all tested groups (icv-STZ versus sham and intact, p < 0.001; sham versus intact, p < 0.05). The level of serum adiponectin in icv-STZ rats was higher compared to sham-operated and intact animals (Fig. 3D). A slightly decreased level of serum adiponectin in sham-operated rats compared to intact rats may reflect craniotomy-induced effect.
Macroscopic examination, histological analysis of liver, pancreas and brain
Macroscopic examination of the abdominal cavity revealed symptoms of intra-abdominal and subcutaneous fat accumulation (Fig. 4A) and fatty liver (yellowish and greasy) in obese icv-STZ rats (Fig. 4B), but not in sham-operated rats (Fig. 4C, D). The abdominal cavities of intact rats were not different from sham-operated rats. Histological analysis of H&E stained liver sections of obese icv-STZ rats showed typical symptoms of steatosis such as intracytoplasmic fat vacuoles and hepatocyte damage (Fig. 5A). No histological signs of the liver steatosis were observed in sham-operated (Fig. 5B) and in intact rats (data not shown). H&E staining revealed pancreatic islet hypertrophy in obese icv-STZ (Fig. 5C). The islets displayed irregular outlines and increased intra-islet deposition of fibrous tissue stained with Masson’s Trichrome (Fig. 5D). IHC analysis of islets showed peripheral localization of glucagon and central insulin staining of beta cells separated into groups by fibrous tissue in icv-STZ rats (Fig. 5E). Intact morphology of islets stained with H&E (Fig. 5F) and with Masson’s Trichrome (Fig. 5G) as well as intensive staining for insulin and glucagon were found in pancreatic islets of sham-operated (Fig. 5H) and intact rats (data not shown).
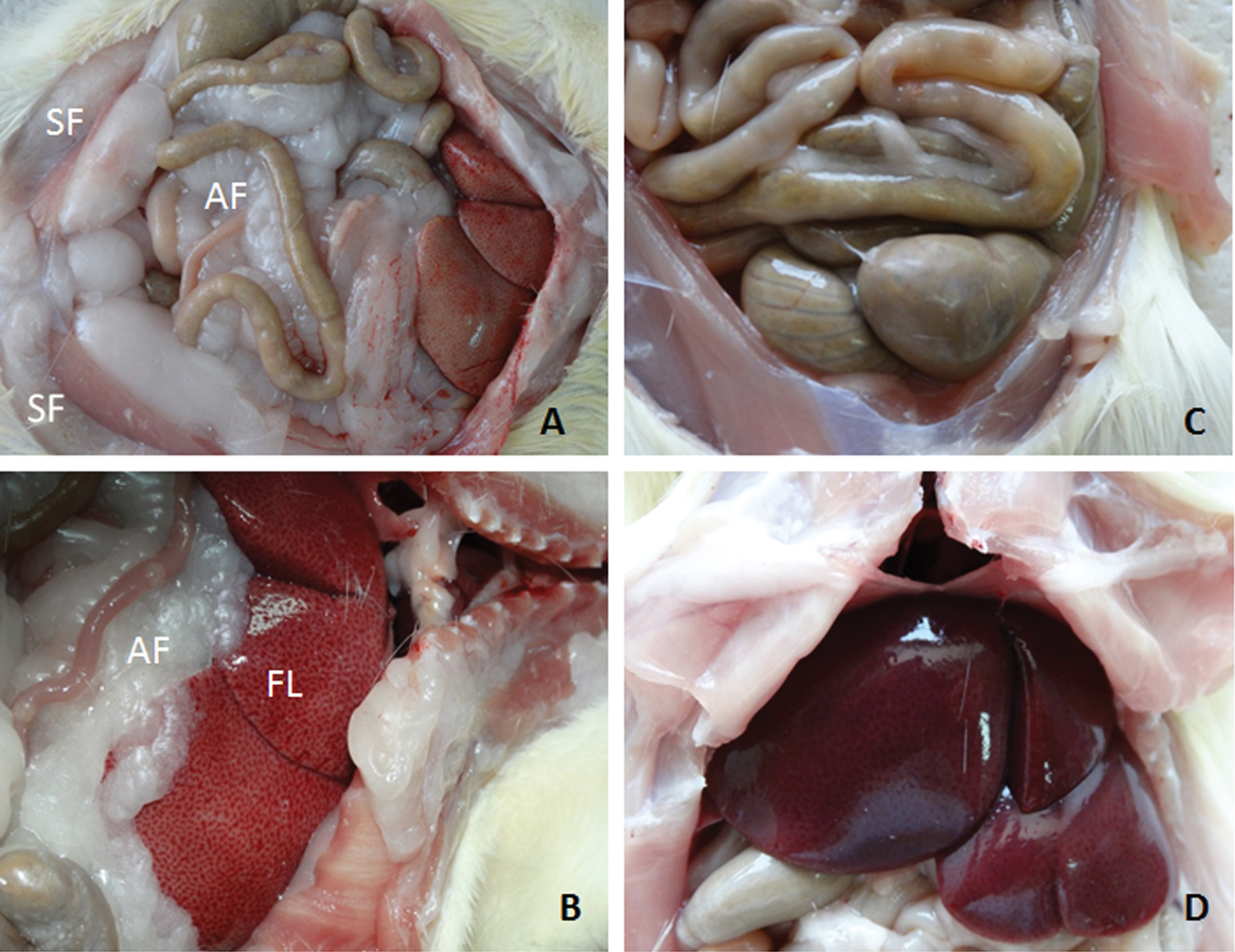
Macroscopic images of abdominal cavity. Three months after icv-administrations, the abdominal cavities of rats were opened. Macroscopic examination revealed symptoms of intra-abdominal and subcutaneous fat accumulation and fatty liver (yellowish and greasy) in obese icv-STZ Lewis rats (Fig. 4A, B), but not in sham-operated lean Lewis rats (Fig. 4C, D). The abdominal cavities of intact rats were not different from sham-operated rats. SF, subcutaneous fat; AF, intra-abdominal fat; FL, fatty liver.
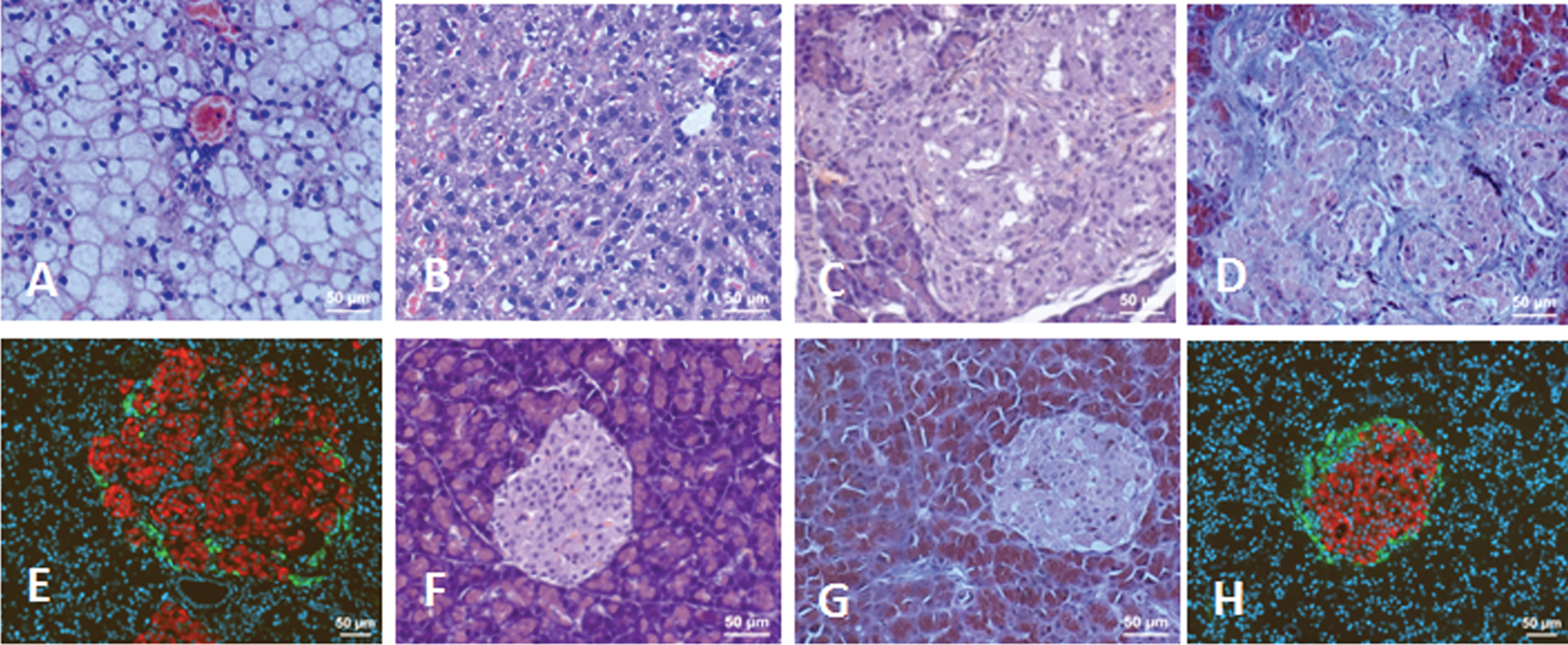
Histology of liver and pancreatic islets of Lewis rats. A) Liver of icv-STZ rat, H&E staining. B) Liver of sham-operated rat, H&E staining. C) Pancreatic islet of icv-STZ rat, H&E staining. D) Pancreatic islet of icv-STZ rat, Masson’s Trichrome staining. E) Pancreatic islet of icv-STZ rat, IHC staining for insulin (red) and glucagon (green). F) Pancreatic islet of sham-operated rat, H&E staining. G) Pancreatic islet of sham-operated rat, Masson’s Trichrome staining. H) Pancreatic islet of sham-operated rat, IHC staining for insulin (red) and glucagon (green). Scale bars = 50 μm.
Aβ expression in the brain
To determine whether obese icv-STZ rats displayed Aβ deposits in the brain, a major hallmark of AD, we used monoclonal antibodies for IHC detection of the antigen. IHC analysis showed vascular but not diffuse Aβ deposition in the brain of obese icv-STZ rats. The Aβ deposits were only found in the blood vessels located in the leptomeningeal cavity of icv-STZ rats (Fig. 6A, B). No signs of Aβ depositions were detected in the brains and leptomeningeal blood vessels of sham-operated (Fig. 6C, D) and intact rats (Fig. 6E, F). In contrast, aged 5xFAD transgenic mice, used as a positive control, demonstrated vascular and diffuse deposition of Aβ in the brain(Fig. 6G, H).

Aβ deposition in icv-STZ Lewis rats and 5XFAD mice. The sagittal sections of the brain were incubated with mouse monoclonal Aβ antibodies. The staining was detected using PolyScan HRP/DAB detection system. A, B) icv-STZ rats. C, D) sham-operated rats. E, F) intact rats. G, H) 5xFAD mice. No positive staining was found in sham and intact groups. Arrows show Aβ deposits in icv-STZ rat and 5XFAD mouse. Scale bars for low magnification (x100) images (A, C, E, G) = 200 μm and scale bars for high magnification (x400) images (B, D, F, H) = 100 μm.
Neuroinflammation
To test whether icv-STZ induces inflammation in the brain, we evaluated activation of astrocytes (astrogliosis) and microglia (microgliosis) in the fimbria-fornix area of hippocampus. Increased level of astrogliosis estimated by GFAP staining (Fig. 7) and microgliosis as evident by increased expression of Iba1 (Fig. 8) were found in icv-STZ rats compared to sham-operated and intact rats. Quantitative analyses for GFAP and Iba1 expression were performed using ANOVA and the Tukey post hoc test. A one way ANOVA for GFAP positive staining showed significant difference between the groups (F (2,6) = 47.43; p < 0.0001). Tukey post hoc tests revealed differences between the icv-STZ rats and animals from both the sham and intact groups (p < 0.001). A one-way ANOVA for Iba1 positive staining showed significant difference between the groups (F (2,6) = 102.9; p < 0.0001). Tukey post hoc tests showed differences between the icv-STZ group and other tested groups (p < 0.001). The expression of GFAP and Iba1in the brain of intact rats was not differ to that of the sham group (Figs. 7 and 8, respectively). In addition, icv-STZ induced spongiform vacuolation (spongiosis) in the fimbria-fornix of the hippocampus while the spongiosis was not found in sham-operated and intact rats.
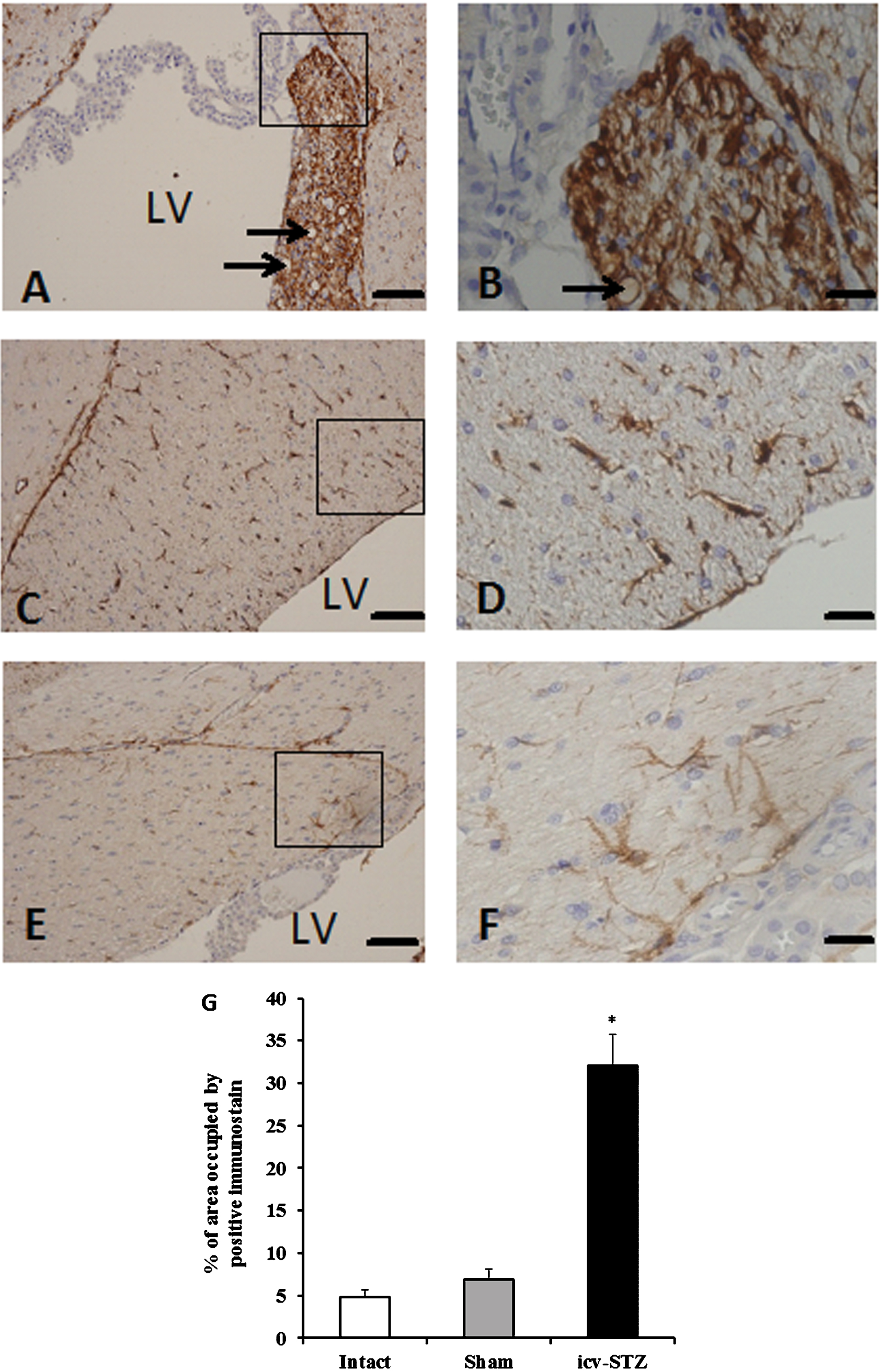
Evaluation of GFAP staining in the fimbria-fornix of hippocampus of icv-STZ (A, B), sham-operated (C, D), and intact rats (E, F). Arrows show spongiform vacuolation (spongiosis). LV, lateral ventricles. Scale bars for low magnification (x100) images A, C, E) = 100 μm and scale bars for high magnification (x400) images (B, D, F) = 20 μm. G) The percentage of binary area occupied by the positive immunostaining. Three rats (seven sections per each rat) were analyzed per each experimental group. *p < 0.001 versus intact and sham groups.

Evaluation of Iba1 staining in the fimbria-fornix of hippocampus of icv-STZ (A, B), sham-operated (C, D), and intact rats (E, F). Arrows show spongiform vacuolation (spongiosis). LV, lateral ventricles. Scale bars for low magnification (x100) images (A, C, E) = 100 μm and scale bars for high magnification (x400) images (B, D, F) = 20 μm. G) The percentage of binary area occupied by the positive immunostaining. Three rats (seven sections per each rat) were analyzed per each experimental group. *p < 0.001 versus intact and sham groups.
Behavioral characteristics
To evaluate effect of icv-STZ on behavioral and cognitive characteristics, the rats from all studied groups were exposed to weekly observations of behavior, as well as to OF and MWM tests. Visual observations of animal behavior showed that during the period of weight decline and initial weight gain (from second to fifth week after icv-STZ), the icv-STZ rats demonstrated symptoms of hyperactive and impulsive behavior such as restlessness, noise anxiety, and tail-tremor. Thereafter, the neurological symptoms in icv-STZ treated rats gradually disappeared. No such neurological abnormalities were found in intact and sham-operated rats.
In the OF test performed 10 weeks after starting the experiments, we measured motility parameters (velocity and distance moved) as well as the time spent in the center of the field as a measurement of anxiety and danger assessment. One way ANOVA tests for the velocity and distance moved showed a significant differences between the groups (F (2,14) = 5.591; p = 0.0164).
Post-hoc test showed that icv-STZ rats were not different from intact and sham-operated animals in terms of velocity and distance moved. Sham-operated rats showed statistically significant decreased motility compared to intact rats (Fig. 9A, B). One way ANOVA test for center field activity found no significant difference between the groups (F (2,14) = 0.6067; p = 0.5589), data not shown.

Open field (OF) test. A) Velocity. B) Distance moved. Lewis rats (n = 9 for icv-STZ group, n = 4 for intact and sham group) exposed to OF and monitored for 20 min. Motility and track exploration were videotaped and analyzed with a Noldus digital system. Data presented as mean±SE. No statistically significant differences were found between icv-STZ group and both sham and intact groups. *p < 0.05 versus intact group.
In the MWM tests, carried out 6 weeks (first MWM test; Fig. 10A) and 12 weeks (second MWM test; Fig. 10B) from the start of the experiments, a multivariable Cox regression time to event approach found that sham-operated animals showed a slight impairment of spatial memory compared to intact animals in the acquisition phase on day 2 (in both tests) and day 3 (first MWM test; Supplementary Table 1), whereas the icv-STZ rats demonstrated severe impairment in spatial memory and learning as expressed by a significantly increased latency to the platform during all tested days compared to intact animals in both tests (Supplementary Table 1). In addition, the performance of icv-STZ rats was significantly compromised compared to sham-operated animals on day 3 of acquisition in the first MWM test (Supplementary Table 1; Fig. 10A). Icv-STZ rats appeared cognitively impaired compared to both intact and sham groups during all days of the second MWM test (Supplementary Table 1; Fig. 10B). Comparison between the first and the second MWM tests revealed a long-term retention of spatial memory in both sham-operated and intact rats: in the second test, the time taken to reach the hidden platform during all acquisition and reversal days was about the same as in the last day of acquisition and reversal phases. Icv-STZ rats also showed long-term retention of spatial memory, but these animals manifested a profound deficit in learning capacity compared to sham-operated and intact rats (Fig. 10A, B).
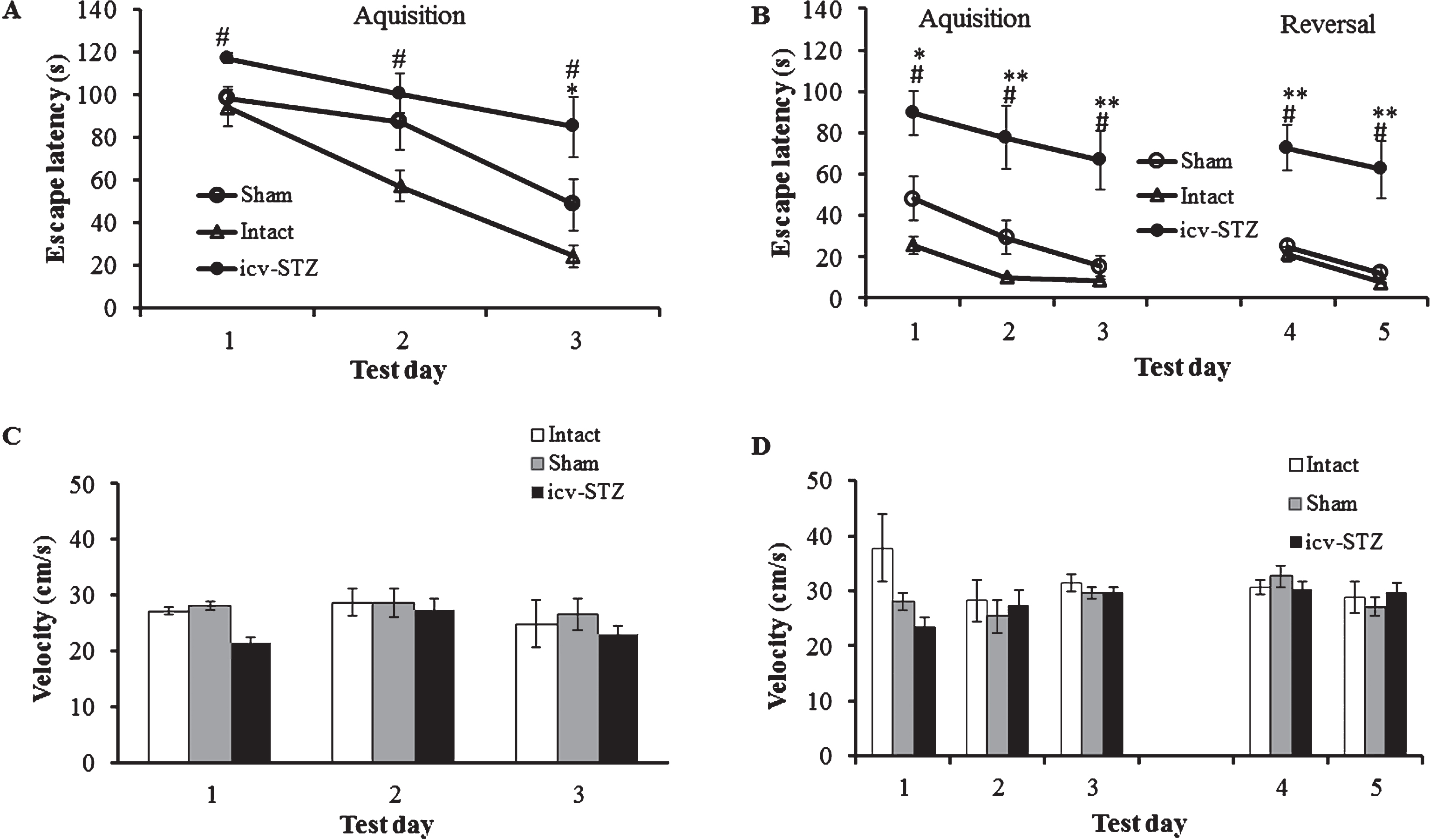
Morris water maze (MWM) task. A, B) Escape latency, 1.5 month and 3 months after the start of the experiments, respectively. C, D) Velocity, 1.5 and 3 months after the start of the experiments, respectively. Each session consisted of four trials (120 s each trail). Data are expressed as means±SE per experimental condition. n = 9 for intact and sham group, n = 13 for icv-STZ group. *p < 0.01 versus intact, #p < 0.05 versus sham, # #p < 0.01 versus sham, **p < 0.05 versus intact and sham groups.

A speculative scheme of the possible mechanism of action of icv administrated STZ in Lewis rats. STZ-induces damage in GLUT2 expressing areas of hippocampus (HC-GLUT2) and hypothalamus (HT-GLUT2) probably leading to development of dementia and peripheral metabolic dysfunctions. In turn, the peripheral metabolic abnormalities might further enhance cognitive dysfunctions leading to development of severe dementia in the obese Lewis rats.
To exclude the possibility that the difference in the latency to the platform is linked to impaired motility due to overweight, we assessed the locomotor activity of each group of animals. A two ways RM-ANOVA test for the velocity (acquisition phase) found no significant difference between the groups in both of the MWM tests (F (2,29) = 2.91; p = 0.07 in the first MWM test and F (2,29) = 2.855; p = 0.074 in the second MWM test). In both first and second MWM tests, no significant differences between the days (F (2,29) = 2.02; p = 0.14 and F (2,29) = 1.32; p = 0.274, respectively) nor interactions (F (4,29) = 0.56; p = 0.68 and F (4,29) = 2.24; p = 0.075, respectively) were found. A two ways RM-ANOVA test for the velocity (reversal phase of the second MWM test) found no significant difference between the groups (F (2,29) = 0.01; p = 0.9891). The test found no significant difference between the days (F (1,29) = 3.49; p = 0.07) nor interaction (F (2,29) = 1.1; p = 0.34; Fig. 10C, D). Thus, dramatic impairment of spatial learning and memory of obese icv-STZ rats cannot be explained by changes in locomotor activity compared to lean sham-operated and intact rats.
DISCUSSION
Previous studies showed that icv-STZ induces AD-like cognitive dysfunctions without alterations in peripheral glucose homeostasis and body weight changes in Wistar or Sprague Dawley rat strains [20, 21], mice and monkeys [27–29]. Of note, genetic differences between rodent strains are known to play an important role in regulation of behavioral and metabolic responses to various environmental stimuli [35, 36]. In the present study, we found that in inbred Lewis rats the icv-STZ induces not only severe cognitive dysfunctions, but also significant changes in body weight associated with peripheral metabolic abnormalities. During the first two-week post-operation period, most of icv-STZ rats lost 15–30% of their initial weight. Thereafter, the STZ treated rats showed accelerated weight gain associated with obesity-related metabolic dysfunctions and cognitive functions decline. Such changes in body weight after icv-STZ resemble the changes in BMI of a subset of AD patients which show body weight decline after clinical AD onset and weight gain in advanced stage of the disease [9]. Importantly, early visual symptoms of hyperactive and impulsive behavior in icv-STZ rats were observed during the period of weight decline and initial weight gain. None of these cognitive dysfunctions and peripheral metabolic abnormalities were found in intact and sham-operated rats.
In OF test, the sham-operated animals showed decreased motility compared to intact rats, but not to icv-STZ rats. Such reduced locomotion might be explained by mild damaging effect of sham surgery. It should be noted that the sham-operated rats were exposed to craniotomy performed by electric drill and icv-administration of vehicle. Recently, it was shown that such type of “sham” surgery (craniotomy alone) causes a mild brain injury leading to impaired motor responses compared to naive rats [37]. In case of icv-STZ rats, the craniotomy-induced reduction of motility in OF test might be neutralized by restless behavior observed in these animals during weight gain period.
The cytotoxic effect of STZ depends on its uptake by GLUT2-expressing cells that are widespread in the brain [25]. In order to induce brain injury, STZ was injected directly into the lateral ventricles [20]. We found that icv-STZ induced Aβ deposition in blood vessels located in the leptomeningeal area of Lewis rats. Moreover, the icv-STZ rats demonstrated an elevated level of activated microglia, astrogliosis, and spongiosis in the fimbria-fornix area of hippocampus, which is one of the most important anatomical structures related to memory and learning [38]. Of note, neuroinflammation (activated microglia and astrogliosis) is considered as a primary driver of brain insulin resistance in AD [39].
In addition to the brain abnormalities, icv-STZ treated Lewis rats also showed severe dysfunctions of peripheral metabolism. 12 weeks after icv-STZ, the rats expressed an obese phenotype associated with subcutaneous and visceral fat deposition, fatty liver, and significantly increased blood levels of insulin, leptin, and adiponectin. Histological analysis of pancreas showed islet hypertrophy, intensive insulin staining, and intra-islet fibrosis. In spite of obese-related metabolic dysfunctions, the icv-STZ rats exhibited uncompromised glucose homeostasis. It can be speculated that in the absence of genetic predisposition to diabetes type 2, a compensatory hyperinsulinemia is able to maintain intact glucose homeostasis in the obese rats. Although mechanisms of brain-centered glucoregulatory systems are poorly understood, growing evidence suggests that the brain can improve or even normalize glycaemia in rodent models of obesity and diabetes [40]. A recent study suggests that the relationships linking peripheral metabolic abnormalities and brain dysfunctions are bidirectional [41]. Early research demonstrated that systemic metabolic dysfunctions, such as diabetes, obesity, and metabolic syndrome may be risk factors for development of neurocognitive disorders. More recent research indicate that cognitive dysfunctions increase the risk of metabolic disorders [42–44]. The data presented in this paper demonstrate that sporadic AD dementia in the Lewis rats induced by icv-STZ increases the risk of metabolic abnormalities associated with obesity. There are several reasons allowing us to suggest that the peripheral metabolic dysfunctions are secondary to the brain damages induced by icv-STZ. Firstly, we used a very low sub-diabetogenic dose (3 mg/kg) of icv-STZ that is about 30 times less than the intravenous dose of STZ required to induce diabetes-related metabolic dysfunctions in Lewis rats [45, 46]. Secondly, the possibility that a low dose of icv-STZ is able to induce direct damage of peripheral tissues is unlikely because of the extremely short half-life, about 15 min [47] and inability of STZ to cross the blood-brain barrier [27, 48]. Thirdly, visual signs of behavioral dysfunctions such as hyperactivity and impulsiveness were observed shortly after icv-STZ administration and they disappeared with progression of peripheral metabolic dysfunctions.
Based on our results showing peripheral metabolic dysfunctions in the icv-STZ rats, as well as on previous data [49] demonstrating expression of GLUT2 and metabolic hormone (leptin, insulin, adiponectin) receptors [50–52] in the hypothalamus, it can be speculated that the hypothalamus might be the main target for the cytotoxic effect of icv-STZ. A deleterious effect in the brain area may be involved in the peripheral metabolic changes, which in turn might lead to enhancement of cognitive dysfunctions related to hypothalamic abnormalities. It is likely that cytotoxic effects of STZ in the brain of Lewis rats lead to both development of severe cognitive dysfunctions and peripheral metabolic abnormalities as shown in the speculative scheme depicted in Fig. 11. However, the precise molecular mechanisms involved in the relationships between central and peripheral metabolic systems are still poorly understood.
To the best of our knowledge, this study is the first to report that icv administration of STZ to Lewis rats leads to development of severe cognitive dysfunctions and body weight changes associated with peripheral metabolic abnormalities. This novel animal model may be useful for to better understanding the pathophysiological relationship between dementia and obesity and provides a new tool for development of an effective therapy for this complex brain-metabolic pathology. Authors’ disclosures available online (http://j-alz.com/manuscript-disclosures/16-1289r2).
Footnotes
ACKNOWLEDGMENTS
The authors thank Dr. Daniel Lazard for fruitful discussions and critical reading of this paper, Prof. Daniel Offen and Yael Barhum for providing us with 5xFAD mice and assistance with stereotactic surgery, and Dr. Ella Koganovsky and Olga Skokov for assistance with histology. This study was supported in part by grants (0601243032 and 0601523471) from Vice President for Research and Development of Tel Aviv University (KB) and Mayer Foundation for Research (AW).