
Abstract
Alzheimer’s disease (AD) is the most common form of dementia, characterized by progressive decline in cognitive abilities of the affected individuals. Biological markers are essential to identify individuals at early stages of the disease for timely therapeutic intervention. Currently, pathological biomarkers are detected either through cerebrospinal fluid analysis or brain imaging, or postmortem, all of which are expensive, invasive, or time consuming. Recently, some studies have shown that circulating miR-125b, miR-181c, miR-9, miR-191-5p, miR-26b-3p, and miR-28-3p may be biomarkers of AD. However, those potential biomarkers are not validated in an AD mouse model. In the current study, we found that circulating miR-125b, miR-9, and miR-191-5p are downregulated, and miR-28-3p is upregulated in an APP/PS1 transgenic mouse model of AD. Furthermore, the correlation analysis shows a positive correlation between the expression of miR-125b and cognitive function of the APP/PS1 transgenic mouse. Moreover, we also determined that the level of serum miR-125b, miR-9, and miR-191-5p were reversed in EGCG-treated APP/PS1 transgenic mouse models. Finally, the expression of miR-125b was significantly downregulated in EGCG-treated SH-SY5Y cells.
INTRODUCTION
Alzheimer’s disease (AD) is a chronic, progressive neurodegenerative disease, which leads to severe cognitive loss and eventual death. Of the dementias, AD is the most common, accounting for between 60% and 70% of all dementias [1]. The characteristic features of AD are the appearance of extracellular amyloid plaques and neurofibrillary tangles in the intracellular environment, neuronal death, and the loss of synapses, all of which contribute to progressive cognitive decline [2, 3]. These pathological changes in the brains of patients with AD represent important targets for diagnosis and treatment [4]. The current methods that are used in AD diagnosis are magnetic resonance imaging, positron emission tomography (PET), and biomarkers in cerebrospinal fluid via lumbar puncture [5, 6]. The validity of these methods is very limited, because they are expensive, invasive, or time consuming. Thus, there is an urgent need for less invasive and affordable blood-based biomarkers that can aid in large-scale screening of patients with AD.
The minimally invasive and potentially inexpensive nature of tests using blood-based biomarkers make these approaches practical to implement, allowing for repeated sampling in large cohorts, and, therefore, might have significant advantages over other biomarker modalities [7, 8]. However, the use of blood as a matrix for measurement of biomarkers has the inherent disadvantage of its complex composition and subsequently poses technical difficulties for biomarker detection. Micro ribonucleic acids (miRNAs) are an abundant class of small non-coding RNAs (approximately 22 nucleotides) that negatively regulate gene expression at the post-transcriptional level [9]. Recent studies have shown that miRNAs are not only active in cells but also present in serum or plasma [10]. Circulating miRNAs may act as potential diagnostic biomarkers for many types of diseases, including AD [11–13]. Recently, some studies have shown that circulating miR-125b [14], miR-181c [14], miR-9 [14], and miR-191-5p [15] were downregulated; miR-26b-3p [16] and miR-28-3p [16] were upregulated in AD and may be potential biomarkers of AD. However, those potential biomarkers of AD were only detected in the blood plasma or serum of patients with AD, and not validated in AD model mouse-APP/PS1 transgenic mouse models. In the current study, we first aimed to validate the level of circulating miR-125b, miR-181c, miR-9, and miR-191-5p in APP/PS1 transgenic mouse models. Furthermore, we also evaluated the correlation of those potential biomarkers of AD with cognitive function in APP/PS1 transgenic mouse models.
In the past decade, green tea polyphenols, particularly its active component epigallocatechin gallate (EGCG), has gained a lot of attention as a potential therapeutic agent for preventing neurodegenerative [17], inflammatory diseases [18], and cancer [19]. Extensive research on EGCG has brought to light its potential to ameliorate AD by improving the morphologic and functional alterations that occur in a natural aging brain, its ability to suppress cognitive dysfunction, increase learning ability, and reduce oxidative damage [20, 21]. To further demonstrate the changes in serum miR-125b, miR-9, miR-191-5p, and miR-28-3p levels associated with AD, we next treated APP/PS1 transgenic mice with EGCG to see whether the level of miRNAs was reversed.
In the current study, we first aimed to validate the level of circulating miR-125b, miR-181c, miR-9, and miR-191-5p in APP/PS1 transgenic mouse models. Furthermore, we also evaluated the correlation of those potential biomarkers of AD with cognitive function in APP/PS1 transgenic mouse models. Moreover, we also evaluated the level of those miRNAs in APP/PS1 transgenic mouse models treated with EGCG. Finally, we detected the level of miR-125b in EGCG-treated SH-SY5Y cells.
MATERIALS AND METHODS
SH-SY5Y cells culture and treatment
SH-SY5Y cells [22, 23] were cultured in Dulbecco modified Eagle medium supplemented with 10% fetal bovine serum and 100 units/mL penicillin and streptomycin. SH-SY5Y cells were cultured in a humidified incubator in 5% CO2 at 37°C. SH-SY5Y cells were seeded in six-well plates at a density of 60% confluence. The culture medium was replaced with serum-free Dulbecco modified Eagle medium and then administered with 32 μmol/L EGCG (Sigma-Aldrich, St. Louis, MO, USA) for indicated times.
APP/PS1 transgenic mouse and treatment
APP/PS1 (APPswe/PSEN1dE9) double Tg mice were obtained from the Model Research Center of Nanjing University, and maintained with a standard diet and sterile water. All animal experiments were carried out in accordance with the guidelines for the care and use of medical animals established by the Ministry of Health, Peoples Republic of China and the ethical standards for laboratory animals of Third Affiliated Hospital of Guangzhou Medical University. Male APP/PS1 mice 4.5 months old were randomly divided into two groups (n = 8 in each group): model groups and EGCG treatment groups (2 mg/kg/day) via intragastric administration for 45 days.
Morris water maze test
The valuation of cognitive function was performed by Morris water maze test [24], which was comprised of a circular 100 cm diameter, 32 cm deep, water-filled pool (23°C±1°C). Different geometric shapes were positioned around the pool to provide spatial orientation. The objective was for the mice to discover a 10 cm diameter translucent acrylic plastic platform located 1 cm below the water surface in the middle of a quadrant of the pool 20 cm away from the wall. Mice were tested three times daily at 15 min intervals for 5 days. The learning and memory of the mice were assessed on the last day of the trial using a probe trial test. The platform was located in a different quadrant to determine whether the mice were able to generalize the task by learning a new position. At the start of each trial, a mouse was carefully placed in the water in any of the three quadrants without the platform and with its head toward the pool wall. The time required for the mouse to find the platform hidden below the water surface was used to assess dynamic changes of learning and memory. If a mouse found the platform within 60 s, it was able to stay on the platform for 10 s. Mice that did not find the platform were carefully assisted to the platform by the experimenter. Between trials, mice were returned to their home cages using a spoon-net to avoid direct contact with the experimenter. Escape latency, indicated as the time to arrive at the platform and the number of times that mice passed by the platform, was monitored by an automated system that recorded and analyzed movements.
miRNA analyses
Isolation of serum RNA
Total serum RNA was harvested extracted from 400 μL volumes of serum using TRIzol (Invitrogen, Carlsbad, CA, USA) and a miRNeasy Mini Kit (Qiagen, Valencia, CA, USA) according to manufacturer’s instructions.
Determination of circulating miRNAs by quantitative real-time polymerase chain reaction
Total RNA (4 μg) was used for reverse transcription using PrimeScript RT reagent Kit Perfect Real Time (Takara Bio Inc., Japan) according to the manufacturer’s protocol. Briefly, the 50 μL reactions were incubated for 30 min at 37°C and 15 s at 85°C and were then stored at 4°C or –20°C until use. The complementary DNA was used for quantitative real-time polymerase chain reaction analysis (qRT-PCR) using SYBR Premix Ex Taq™ (Takara Bio Inc., Japan) and a Roche capillary-based Light Cycler 2.0 Systems (Roche Diagnostics Corporation, Indianapolis, IN, USA) according to the manufacturer’s protocol. In short, the reactions were incubated at 98°C for 30 s, followed by 40 cycles of 95°C for 10 s and 60°C for 30 s. A dissociation stage was performed at the end of the amplification procedure to determine potential nonspecific amplifications. To adjust for variations in RNA extraction, synthetic Caenorhabditis elegans miRNAs, cel-miR-39 [25, 26] (C. elegans miR-39 miRNA mimic: 5’-UCACCGGGUGUAAAUCAGCUUG-3’) were added to the samples after the initial denaturation. The relative expression level of miRNAs was calculated using the relative threshold cycle and the comparative threshold cycle method. Briefly, miRNAs expression was calculated after normalization by cel-miR-39 and was then expressed as 2–ΔCq, in which ΔCqmiRNAs = CqmiRNAs – Cqcel-miR-39. For the primer design, we first found out the sequence of miRNAs from NCBI, and then used Primer 5.0 to design primer. For the quality control, ddH2O was used for negative control. The specificity of resulting PCR products was confirmed by melting curves (data not show). The miRNAs data are presented as the mean±standard error. Primer sequences are as follows:
The detection of secretive miR-125b level in culture supernatant
MicroRNA levels were measured in the whole cell culture supernatant as described previously [27, 28]. To collect conditioned media for RNA extraction, cells were cultured in full media at a 60% confluence and subsequently treated EGCG in FBS-deprived media for further 48 h. After collection, media was spun at 2,000 g for 10 min at 4°C and subsequently filtered (0.22 μm) to remove cellular debris and apoptotic bodies. 500 μL of cell supernatant were using for RNA extraction and performed quantitative real-time polymerase chain reaction.
Statistical analyses
All data were expressed as mean±standard deviation. Statistical analyses were performed with SSPS 17.0. Following normality and equality of variance tests, miRNAs were compared using an independent sample t test, with a threshold for significance of 0.05. Relative expression levels of miRNAs were calculated by the 2–ΔCq method. The miRNeasy serum spike-in C. elegans miR-39 (cel-miR-39) was used as the normalized external control. Difference in circulating miR-125b, miR-181c, miR-9, miR-191-5p, miR-26b-3p, and miR-28-3p expression in APP/PS1 mice were assessed using an independent sample t test. Intergroup comparisons were assessed by ANOVA using SPSS statistical software. We corrected for multiple testing using Bonferroni method. p < 0.05 was considered statistically significant. The levels of miRNAs were plotted using GraphPad Prism. Pearson correlation coefficient was calculated to estimate the correlations between miRNA levels and cognitive function, and values of p < 0.05 were considered statistically significant.
RESULTS
The level of circulating miR-125b, miR-181c, miR-9, miR-191-5p, miR-26b-3p, and miR-28-3p in APP/PS1 transgenic mouse
To verify whether the expression of circulating miR-125b, miR-181c, miR-9, miR-191-5p, miR-26b-3p, and miR-28-3p were similar to that of patients with AD, we selected AD animal models – APP/PS1 transgenic mouse models as research objects. The data showed that serum miR-125b, miR-9, and miR-191-5p were significantly downregulated in APP/PS1 mouse models compared with control groups (p < 0.001, n = 8); serum miR-28-3p level was distinctly upregulated in APP/PS1 transgenic mouse models, but serum miR-181c and miR-26b-3p level did not change (Fig. 1), which were slightly different from those of patients with AD.
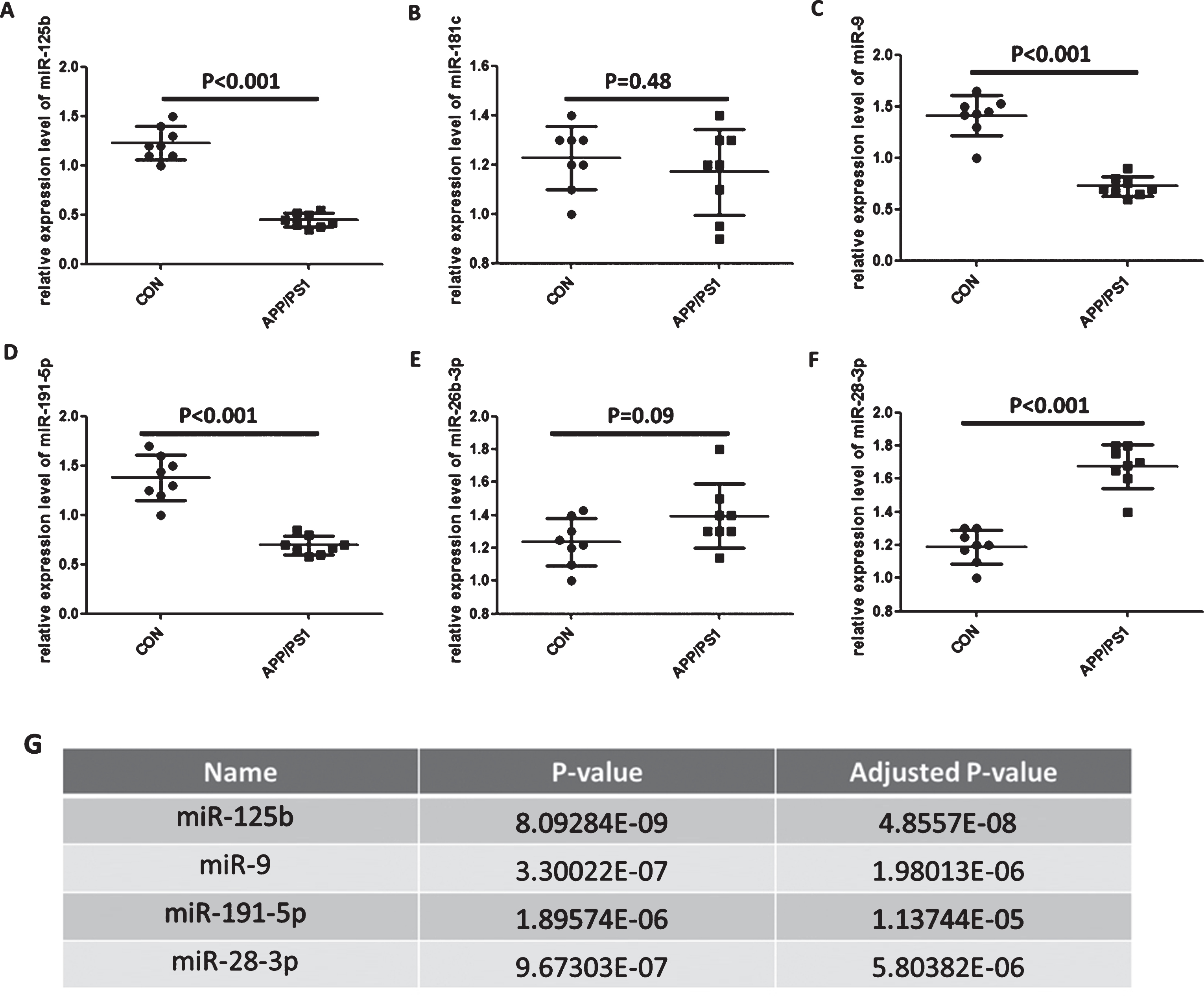
Identification of the level of circulating miR-125b, miR-181c, miR-9, miR-191-5p, miR-26b-3p, and miR-28-3p in APP/PS1 transgenic mouse model. A-F) Measurement of circulating miR-125b, miR-181c, miR-9, miR-191-5p, miR-26b-3p, and miR-28-3p level in APP/PS1 transgenic mouse. The level of circulating miR-125b, miR-181c, miR-9, miR-191-5p, miR-26b-3p, and miR-28-3p were evaluated by quantitative real-time polymerase chain reaction. Bar corresponds to mean±standard deviation. ***p < 0.001 as compared with control group. G) The levels of circulating miR-125b, miR-9, miR-191-5p, and miR-28-3p were corrected using Bonferroni method. p < 0.05 was regarded as statistically significant. ***p < 0.001 as compared with control group.
Correlation between the cognitive function and the level of serum miR-125b, miR-181c, miR-9, miR-191-5p, miR-26b-3p, and miR-28-3p in APP/PS1 transgenic mouse
We used the Morris water maze test (Escape latency, Number of crossing of platform, and Time spent in platforms quadrant) to evaluate the cognitive function of APP/PS1 transgenic mouse models. As shown in Fig. 2A-C, Morris water maze test (Escape latency, Number of crossing of platform and Time spent in platforms quadrant) showed that there were significant differences in cognitive function of APP/PS1 transgenic mouse models compared with control groups. We next analyzed the potential correlation between the cognitive function and the levels of serum miR-125b, miR-181c, miR-9, miR-191-5p, miR-26b-3p, and miR-28-3p. Pearson correlation analysis results showed that a positive correlation between the relative expression of miR-125b and cognitive function (Escape latency, r = 0.8719, p = 0.0048; Number of crossing of platform, r = 0.9453, p = 0.0004; Time spent in platform quadrant, r = 0.9330, p = 0.0007 respectively), not statistically significant in other miRNAs (Fig. 2D-G).
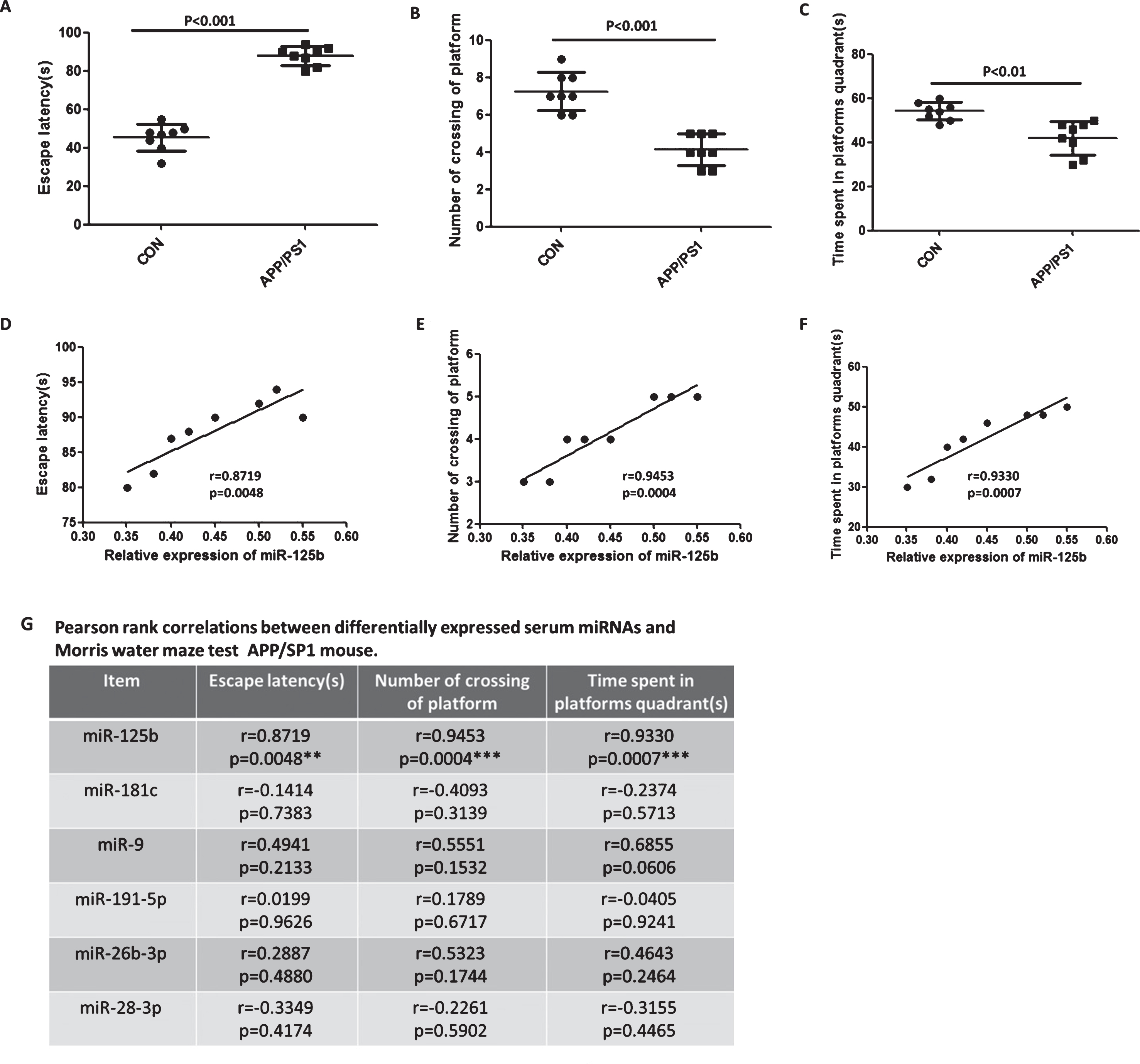
Correlation between the cognitive function and the level of serum miR-125b, miR-181c, miR-9, miR-191-5p, miR-26b-3p, and miR-28-3p in APP/PS1 transgenic mouse model. Cognitive function of APP/PS1 transgenic mouse was tested by Morris water maze. Escape latency (A), Number of crossing of platform (B), and Time spent in platform quadrant (C) were validated by Morris water maze test. Bar correspond to mean±standard deviation. p < 0.05 was considered statistically significant. D-F) The levels of serum miR-125b correlate with cognitive function. A positive correlation between the relative expression of serum miR-125b and cognitive function (Escape latency, r = 0.8719, p = 0.0048; Number of crossing of platform, r = 0.9453, p = 0.0004; Time spent in platform quadrant, r = 0.9330, p = 0.0007, respectively). G) Pearson rank correlation between differential expression of serum miRNAs and Morris water maze test in APP/PS1 transgenic mouse model. p < 0.05 was considered statistically significant. *p < 0.05, **p < 0.01, ***p < 0.001 as compared with control group.
The level of circulating miR-125b, miR-9 and miR-191-5p were reversed in EGCG-treated APP/PS1 transgenic mouse
To further demonstrate the changes in serum miR-125b, miR-9, miR-191-5p, and miR-28-3p levels associated with AD, we treated APP/PS1 transgenic mouse models with EGCG, which improved APP/PS1 transgenic mouse disease status, and then detected serum miR-125b, miR-9, miR-191-5p, and miR-28-3p levels. As shown in Fig. 3, the level of serum miR-125b, miR-9, and miR-191-5p were reversed in EGCG-treated APP/PS1 transgenic mouse models, but there was no change in the miR-28-3p level, indicating that the changes of serum miR-125b, miR-9, and miR-191-5p levels were related to AD, rather than miR-28-3p.
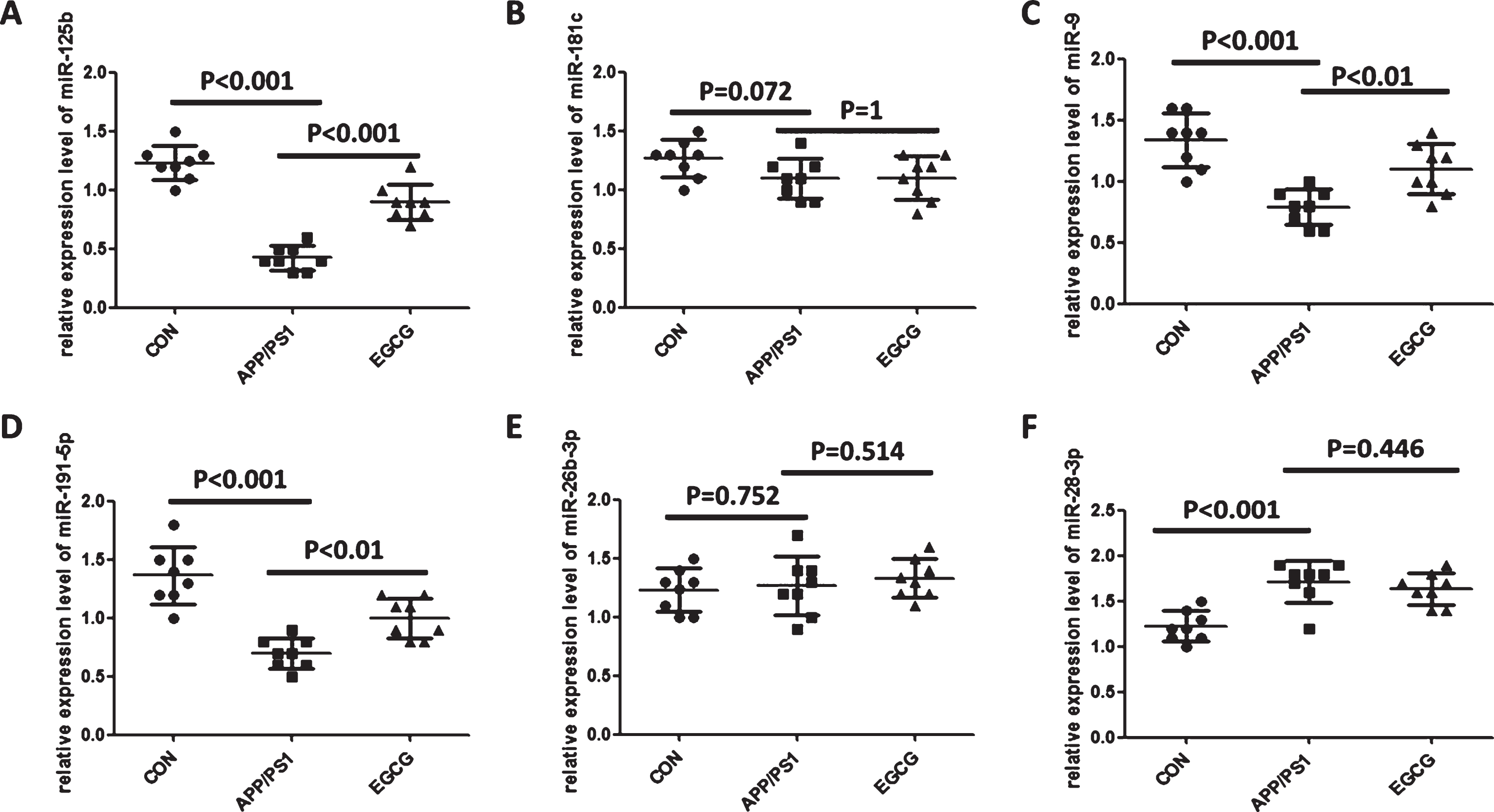
The levels of circulating miR-125b, miR-9 and miR-191-5p were reversed in EGCG-treated APP/PS1 transgenic mouse model. A-F) circulating miR-125b, miR-181c, miR-9, miR-191-5p, miR-26b-3p, and miR-28-3p level in APP/PS1 transgenic mouse model treated with EGCG. Bar corresponds to mean±standard deviation. p < 0.05 was considered statistically significant.
EGCG significantly increased the level of miR-125b in SH-SY5Y cells
To further identify the relationship between miR-125b and AD, in addition to animal experiments, we also treated SH-SY5Y cells with EGCG and detected secretive miR-125b level in culture supernatant. Unfortunately, the level of miR-125b was not detectable in culture supernatant (data not shown). However, the level of miR-125b was significantly increased in EGCG-treated SH-SY5Y cells (Fig. 4), which was in accordance with that of EGCG-treated APP/PS1 transgenic mouse models.
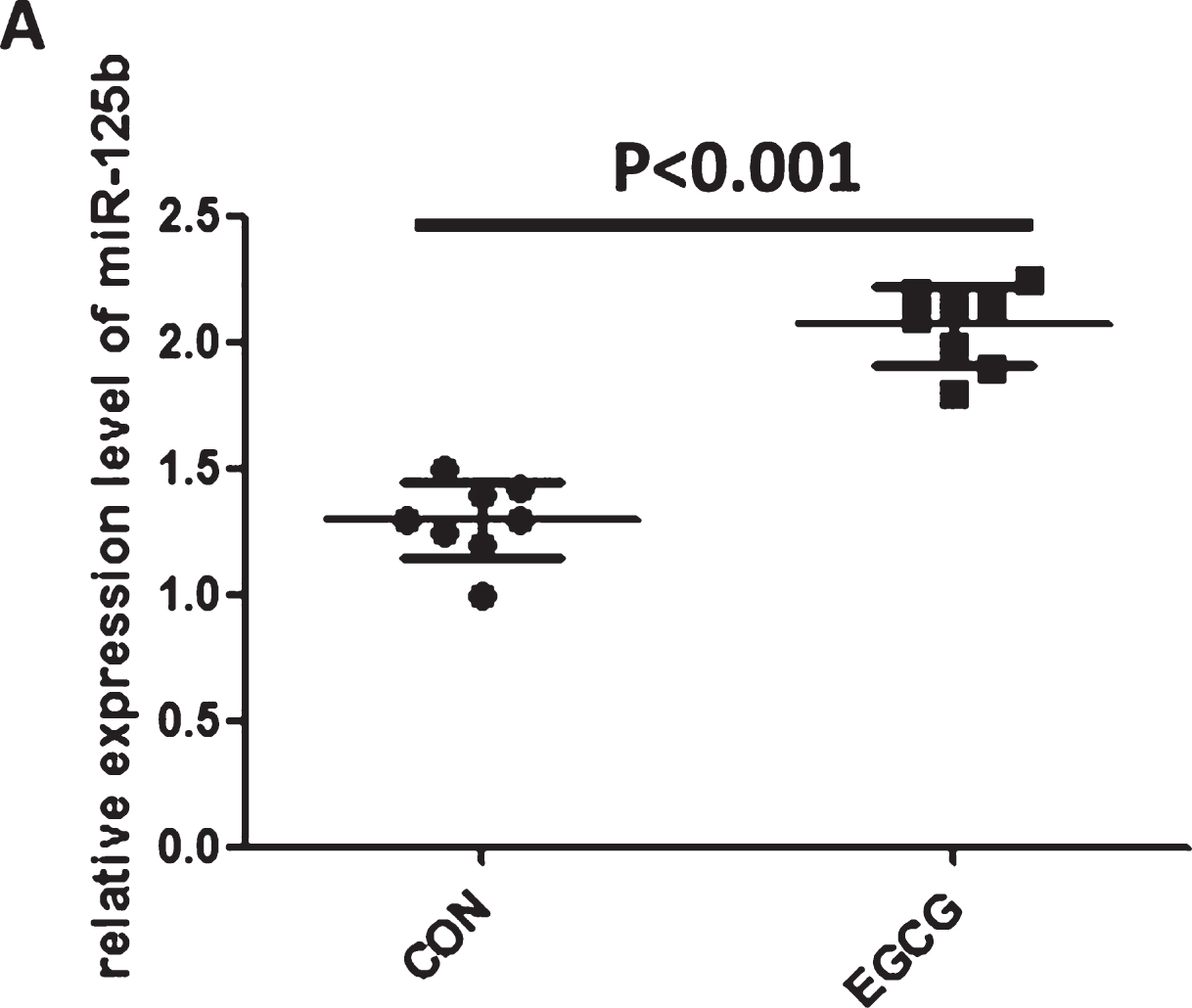
EGCG significantly increased the level of miR-125b in SH-SY5Y cells. The level of miR-125b was upregulated in EGCG-treated SH-SY5Y cells. The level of miR-125b was evaluated by quantitative real-time polymerase chain reaction. Bar corresponds to mean±standard deviation. ***p < 0.001 as compared with control group.
DISCUSSION
Dementia in the elderly population is most commonly caused by AD. The characteristic features of AD are the appearance of extracellular amyloid plaques and neurofibrillary tangles in the intracellular environment, neuronal death, and the loss of synapses, all of which contribute to cognitive decline in a progressive manner [2, 29]. A number of hypotheses have been advanced to explain the mechanism of AD. Abnormal tau phosphorylation and amyloid protein deposition are the most pathological changes and contribute to the formation of abnormal neurofibrillary and amyloid plaque structures [30]. AD is a terminal and incurable disease [31]. In AD, the early stages of neural degeneration appear in the transentorhinal cortex and extend to the entorhinal cortex and hippocampus. Later, the degeneration spreads toward the temporal, frontal, and parietal lobes, leading to the loss of neurons and synaptic contacts [32]. Impairment in neurogenesis is relevant to AD progression as the disease affects the two neurogenic niches in the brain [33, 34]. There is a growing need for a biomarker in AD as recent clinical findings suggest that successful treatment needs to start in the prodromal stages of the disease [35]. Currently, the biomarkers used most extensively in clinical trials for AD diagnosis are structural magnetic resonance imaging, molecular imaging of amyloid deposition using PET, and cerebrospinal fluid measures of Aβ and tau [5, 36]. However, the invasive examination of lumbar puncture and expensive tests of amyloid PET have restricted their use in AD diagnosis and risk assessment. The minimally invasive and potentially inexpensive nature of tests using blood-based biomarkers make these approaches practical to implement, allowing for repeated sampling in large cohorts, and, therefore, might have significant advantages over other biomarker modalities [7, 8].
miRNAs are small, endogenous, nonprotein coding RNAs, originally discovered in the nematode C. elegans and found to be present in all plants and animals [37]. miRNAs can be released into the extracellular environment by binding to RNA binding proteins or transported through body fluids in vesicles such as exosomes and microvesicles [38]. Circulating miRNAs are considered to be reproducible and consistent among individuals of the same species and exhibit altered expression under certain disease states [39]. They are desirable candidate biomarkers for many diseases, including AD, because they remain stable over time, even after repeated freeze-thaw cycles in plasma and serum [40, 41]. Recently, some studies showed that circulating miR-125b [14], miR-181c [14], miR-9 [14], and miR-191-5p [15] were downregulated; miR-26b-3p [16] and miR-28-3p [16] were upregulated in AD and may be potential biomarkers of AD. However, those potential biomarkers of AD were only confirmed in the plasma or serum of patients with AD and with correlation analysis, and not manifested in animal experiments. In this study, we found that serum miR-125b, miR-9, and miR-191-5p were downregulated; miR-28-3p was upregulated in AD animal models APP/PS1 transgenic mice, which were consistent with that of patients with AD [14], but no changes in miR-181c and miR-26b-3p level, indicating that circulating miR-181c and miR-26b-3p were only a random variable, independent of AD diagnosis. In addition, we detected the level of these mRNAs in hippocampus of APP/PS1 mice. We found that the levels of miR-125b, miR-26b-3p, and miR-9 were consistent with those in serum, but there was no change in miR-181c, miR-191-5p, and miR-28-3p (Supplementary Figure 2).
Green tea (Camellia sinensis) is an extremely popular beverage worldwide, and its habitual consumption has been associated with health benefits including chemo-preventive efficacy [42]. In the past decade, green tea polyphenols, particularly its active component EGCG, have gained a lot of attention as a potential therapeutic agent for preventing neurodegenerative [17], inflammatory diseases [18], and cancer [19], mainly because of their beneficial effects on human health. This ability is mostly attributed to their anti-oxidant [17], radical scavenging [17], metal chelating [17, 19], anti-carcinogenic [19], anti-apoptotic [17], and anti-inflammatory properties [18]. Extensive research on EGCG has brought to light its potential to improve AD by improving the morphologic and functional alterations that occur in a natural aging brain, its ability to suppress cognitive dysfunction, increase learning ability, and reduce oxidative damage via the ability to cross the blood-brain barrier [20, 44]. To further demonstrate circulating miR-125b, miR-181c, miR-9, miR-191-5p, miR-26b-3p, and miR-28-3p correlation with AD, we treated APP/PS1 transgenic mouse models with EGCG. As shown in Fig. 3, the levels of serum miR-125b, miR-9, and miR-191-5p were reversed compared to APP/PS1 transgenic mice, but there was no change in miR-28-3p level, indicating that the changes of circulating miR-28-3p level was not correlated with patients with AD. To further valuate circulating miR-125b, miR-181c, miR-9, miR-191-5p, miR-26b-3p, and miR-28-3p correlated with AD, we also performed correlation analysis. The data showed that only serum miR-125b has positive effects with cognitive function of EGCG-treated APP/PS1 mouse models, implying serum miR-125b was the most potential biomarker of patients with AD.
SH-SY5Y cells, a neuroblastoma cell line that closely resembles primary neurons, may be used instead of primary neurons in AD [22, 23]. In vitro assay, we also treated SH-SY5Y cells with EGCG, and detected secretive miR-125b in culture supernatant to further identify miR-125b as a potential biomarker of AD. Unfortunately, the level of miR-125b was not detectable in cell culture supernatant (data not shown), but the level of miR-125b was markedly upregulated in EGCG-treated SH-SY5Y cells. To exclude the possibility that upregulation of miR-125b induced by EGCG is independent from AD-related condition, we also detected the Aβ1–42 level in cell culture supernatant. The data showed that EGCG significantly inhibited secretion of Aβ1–42 (Supplementary Figure 1). The expression of secretive miR-125b in cell culture supernatant was negative, which may be the result of insufficient secretion.
In conclusion, we first evaluated the level of circulating miR-125b, miR-181c, miR-9, miR-191-5p, miR-26b-3p, and miR-28-3p in AD animal models-APP/PS1 transgenic mouse models, and demonstrated the levels of circulating miR-125b, miR-9, miR-191-5p, and miR-28-3p were consistent with that of patients with AD. Furthermore, improvement of cognitive function of APP/PS1 transgenic mouse models occurred during treatment with EGCG, the levels of circulating miR-125b, miR-9, miR-191-5p were reversed, and only miR-125b showed a positive correlation with cognitive function of EGCG-treated APP/PS1 transgenic mice. Our results indicated that circulating miR-125b was the most likely to be a potential biomarker of AD.
Footnotes
ACKNOWLEDGMENTS
This study was supported by Zhengdatianqing Foundation of China, the doctoral startup foundation of Guangzhou Medical University (Grant Number: 2015C24), and Guangzhou City Health Bureau General guidance project (Grant Number: 20171A011312). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript. Honghai Hong is responsible for the experimental operation and study design; Yang Li is responsible for animal experiments, study design and revising the manuscript; Baochang Su is responsible for the study design and drafting the work.