
Abstract
Alzheimer’s disease (AD), the most frequent type of dementia, is a prototypical neurodegenerative disease, but shares with stroke certain common risk factors. Consequently, how vascular pathology may modulate AD pathogenesis has gained scientific attention. Therefore, aside from typical features of AD (e.g., amyloid-β, tau hyperphosphorylation, and cholinergic dysfunction), changes within the ‘neurovascular unit’ (NVU) are of particular interest. This study focused on cholinergic, choline acetyltransferase (ChAT)-immunopositive, and tyrosine hydroxylase (TH)-containing neurons in association with the vasculature to explore the neurovascular complex of the AD brain affected by stroke. Wild-type and triple-transgenic (3xTg) mice of different ages underwent unilateral permanent focal cerebral ischemia. Histochemical analyses comprised diverse neuronal and vascular NVU components, and markers of AD. Immunofluorescence labeling confirmed the existence of Aβ deposits and phospho-tau together with glial reactions and morphologically altered endothelia, visualized by Solanum tuberosum lectin. Twenty-four hours after ischemia induction, immunoreactivities for ChAT and TH declined in the ischemia-affected striatum and, at least in part, in the ischemic border zone and ipsilateral neocortex. Correlation analyses indicated simultaneous degeneration of neuronal and vascular components. A trend for more severe affection of ChAT was observed in younger as compared with older mice. The present findings suggest complex interactions within the NVU of the AD-like brain affected by ischemia, comprising alterations of the cholinergic system in conjunction with vascular pathology. Hence, it may be worthwhile to explore the impact of a cellular stabilization approach on vascular and glial elements in AD in terms of a potential disease-alleviating strategy.
Keywords
INTRODUCTION
Alzheimer’s disease (AD) accounts for the leading cause of dementia and is characterized by a progressive cognitive impairment, associated with an enormous socioeconomic burden due to wide-ranging consequences for patients themselves, their relatives, and the health system [1]. Extensive research during the last decades identified (i) the accumulation of amyloid-β (Aβ) forming cerebral plaques, (ii) the hyperphosphorylation of tau resulting in neurofibrillary tangles, and (iii) synaptic dysfunction as pathophysiological hallmarks causing neuronal degeneration [2, 3]. Despite accumulating knowledge on the cellular alterations and advances concerning biomarker-based disease identification, a targeted treatment at the cause of AD is still lacking.
Although ischemic stroke is a separate disease, characterized by neuronal cell death that is typically traced back to an acute vessels occlusion in the brain [4, 5], the socio-economic burden and the situation of still limited treatment strategies appear comparable with AD. Driven by accumulating evidence from experimental and epidemiological studies, parallels of AD and ischemic stroke became recently evident [6, 7], which might allow new insights into disease-related processes and novel specific treatment approaches [8, 9]. Notably, arterial hypertension, hypercholesterolemia, and diabetes were identified as risk factors for both ischemic stroke and AD [10 –13]. Age appears as a crucial factor that could explain some of these overlaps from a statistical perspective, since the frequency of cardiovascular risk factors is known to increase over a life time, simultaneously to an increasing rate of stroke and AD [1, 4]. However, growing evidence emerged from numerous experimental studies and clinical observations, suggesting a causal relationship between cerebral ischemia and AD [14, 15]. In animal models, both chronic hypertension and experimental ischemic stroke were robustly demonstrated to cause AD-like changes such as, for instance, increased levels of amyloid-β protein precursor and tau hyperphosphorylation [16 –18]. On the other hand, vascular involvement including cerebral amyloid angiopathy as well as the pre-existing accumulation of Aβ deposits were found to be associated with an increased susceptibility to ischemic events in terms of an aggravated neuronal injury [19 –22].
Consequently, the variety of well-established factors associated with AD – for instance chronic hypoperfusion due to a cerebrovascular dysfunction [23 –25] – may provide connecting points for elaborated treatment approaches. However, for stroke as well as AD, several attempts to replicate successful animal research in the clinical situation have failed, resulting in a so-called ‘translational roadblock’ [26, 27]. In ischemia research, the view of tissue affection was widened toward a more complex perspective now including neurons with the associated vasculature as well as micro- and astroglia, summarized as the ‘neurovascular unit’ (NVU) by del Zoppo [28, 29]. Remarkably, age-related processes like the deposition of Aβ were already considered to critically affect the integrity within in the NVU, even with consequences on ischemia susceptibility, and thus influencing the outcome of therapeutic approaches like recanalization strategies in acute ischemic stroke [30, 31].
During the last decade, the perspective of a complex neurovascular pathology has also been considered for AD, emphasizing the potential role of the vasculature and their interplay with neurons and associated glial cells during disease progression [32 –35]. While previous studies in the field of experimental AD research in conjunction with ischemic stimuli mostly addressed the effect of ischemia on AD characteristics like tau hyperphosphorylation [36, 37] and the amount of Aβ [17, 38], only little is known about ischemia-related changes within the NVU. Our group has recently shown that focal cerebral ischemia subjected to genetically altered mice and respective wild-type controls leads to considerable alterations of the vasculature in terms of a pronounced degeneration of endothelial cells and astrocytic endfeet in animals with an AD-like background [39]. However, the relationships between the vasculature and neuronal subpopulations with relevance to AD were not yet addressed systematically under ischemic conditions. The expected data might help to understand the complex neurovascular pathology in the AD-ischemia context in more detail and might also help to establish more specific treatment strategies.
This study aimed to explore ischemic consequences on two neuronal subpopulations in close regional relationship to the vasculature within brain areas affected by experimental stroke, while considering potential age-related effects. Emphasis was put on choline acetyltransferase (ChAT) as an enzyme whose decline in activity was previously found to be associated with an age-dependent degeneration of cholinergic neurons in the basal forebrain [40, 41]. In addition to a pivotal role regarding impaired cognitive function [42], the degeneration of the cholinergic system is widely accepted to be involved in the progression of AD, i.e., by mediating Aβ deposition and tau phosphorylation [43 –45]. As a counterpart to the cholinergic system, analyses also addressed catecholaminergic neurons by immunolabeling of their marker enzyme tyrosine hydroxylase (TH).
MATERIALS AND METHODS
Study design and content
A total of n = 24 adult mice from both sexes, affected by 24 h of unilateral permanent focal cerebral ischemia, were investigated concerning histopathological alterations in the forebrain, while Sv129/B6 wild-type mice (3 and 12 months of age: n = 6 each) were compared with age-matched triple-transgenic (3xTg) mice (3 and 12 months of age: n = 6 each). The transgenic animals harbored two mutant human transgenes (APPSwedish mutation + tauP301L)-driven by neuron-specific Thy1-regulatory elements-and the homozygous knock-in construct presenilin-1M146V [46, 47]. In addition, three 2-year-old 3xTg mice were included to verify age-dependent Alzheimer-like alterations in the used transgenic animal model [47 –49]. All animals were bred in the Medizinisch-Experimentelles Zentrum at the University of Leipzig, based on breeding pairs obtained from Drs. Frank M. LaFerla and Salvatore Oddo (University of California, Irvine, CA, USA). Animal experiments were carried out according to the European Communities Council Directive (86/609/EEC) after protocol approval by the Regierungspräsidium Leipzig (reference number TVV 24/10).
Experimental stroke induction
Permanent focal cerebral ischemia was induced according to the filament model as originally described by Longa et al. [50] with minor modifications, causing a right-sided middle cerebral artery occlusion. In detail, a standardized silicon-coated 6-0 monofilament (Doccol Corporation, Redlands, CA, USA) was inserted into the right external carotid artery and moved forward into the internal carotid artery until bending was observed or resistance was felt, indicating correct position of the filament tail at the origin of the middle cerebral artery. For surgical procedures, animals were anesthesized with etomidate (33 mg/kg body weight i.p.; Hypnomidate, Janssen-Cilag, Neuss, Germany). Additionally, local anesthesia of the ventral neck was ensured by s.c. injection of lidocaine (Xylocitin 1%, mibe, Brehna, Germany). Anesthesia-associated cooling was prevented by keeping the body temperature constantly around 37.0°C with a thermostatically controlled warming pad including a rectal probe (Fine Science Tools, Heidelberg, Germany). Following surgery, mice were placed on a commercially available warming pad until recovery. Neurobehavioral deficits were assessed on the following day after ischemia onset using a score introduced by Menzies and co-workers, ranging from 0 (no deficit) to 4 (spontaneous contralateral circling) [51]. To enter the study, animals had to present a score of at least 2, indicating sufficient unilateral focal cerebralischemia.
Tissue preparation for histochemistry
Twenty-four hours after ischemia induction, mice were deeply anesthetized by a mixture of ketamine (150 mg/kg body weight i.p.; Ketamin-ratiopharm, ratiopharm, Ulm, Germany) and xylazine (15 mg/kg body weight i.p.; Rompun, Bayer, Leverkusen, Germany) and transcardially perfused with saline and 4% phosphate-buffered paraformaldehyde. Next, the brains were carefully removed from the skulls, post-fixed in the same fixative overnight and then equilibrated in 30% phosphate-buffered sucrose. Forebrains from all animals were serially sectioned in a standardized manner with a freezing microtome (SM2000R; Leica, Nussloch, Germany). Thereby, resulting series comprised each tenth of the 30 μm-thick coronal sections, which were collected in 0.1 M Tris-buffered saline with pH 7.4 (TBS) containing sodium azide.
Multiple fluorescence labeling
In general, staining procedures were started by extensive washes of free-floating sections with TBS. Afterwards, non-specific binding sites of tissues (for subsequently used immunoreagents) were blocked with 5% normal donkey serum (Dianova, Hamburg, Germany, as supplier for Jackson ImmunoResearch, West Grove, PA, USA) in TBS containing 0.3% Triton X-100 (NDS-TBS-T) for at least 1 h. Serial staining experiments included about 20 sections from each forebrain that were applied to double fluorescence labeling of the endothelial marker Solanum tuberosum (potato) lectin (STL) and antibodies directed against either ChAT or TH. Thereby, sections were incubated with mixtures of biotinylated STL (Vector, Burlingame, CA, USA; 20 μg/ml TBS-NDS-T) and either affinity-purified goat-anti-ChAT (AB144P; Merck Millipore, Billerica, MA, USA; 1 : 50) or rabbit-anti-TH (AB152; Millipore; 1 : 200) overnight. The sections were then rinsed with TBS and reacted for 1 h with mixtures of carbocyanine (Cy)2-conjugated streptavidin and Cy3-coupled donkey antibodies recognizing goat IgG and rabbit IgG, respectively. All fluorochromated immunoreagents were obtained from Dianova and used at 20 μg/ml TBS containing 2% bovine serum albumin (TBS-BSA). Selected sections from 3-month- and 2-year-old animals were used for triple fluorescence labeling as listed in Table 1. Thereby, all primary antibodies were applied overnight at room temperature. Finally, all sections were extensively rinsed with TBS, briefly washed with distilled water, mounted onto slides, air-dried and coverslipped with Entellan in toluene (Merck). Control experiments were performed by omitting the primary antibodies, resulting in the absence of cellular staining.
Triple fluorescence labeling
*All fluorescent immunoreagents were obtained from Dianova (Hamburg, Germany) as supplier for Jackson ImmunoResearch (West Grove, PA, USA) and used at 20 μg/ml for 1 h. **Thioflavin S staining of sections mounted onto Superfrost slides after Cy3-immunolabeling of Aβ and Cy5-immunostaining with AT8. ***Anti-digoxin is highly cross-reactive with the aglycon digoxigenin. ****Carbocyanine-tagged anti-goat IgG antibodies are cross-reactive with sheep IgG and appropriate for the visualization of primary sheep antibodies. Aβ, amyloid-β; Iba, ionized calcium binding adapter molecule 1; GFAP, glial fibrillary acidic protein; STL, Solanum tuberosum lectin (potato lectin); ChAT, choline acetyltransferase; TH, tyrosine hydroxylase.
Fluorescence microscopy und quantification of histopathological findings
All stained sections were screened with an Axioplan fluorescence microscope (Zeiss, Oberkochen, Germany). Images for qualitative analyses were captured with the confocal laser-scanning microscope LSM 510 Meta (Zeiss) as well as the phase contrast microscope Biorevo BZ-9000 (Keyence, Neu-Isenburg, Germany). To create panels, images were further processed applying Microsoft PowerPoint (version 2015; Microsoft Corp., Redmond, WA, USA). If necessary, brightness and contrast of micrographs were slightly modified, but without deletion or creation of signals.
For quantification, micrographs captured with the Biorevo BZ-9000 (Keyence) microscope were further processed. By using 3 consecutive brain sections with a distance of 300 μm, and given the fact that the middle section is exhibiting the most pronounced ischemic lesion, subsequent analyses focused on a forebrain area of about 600 μm in the rostral-occipital direction. In each of the 3 brain sections, 6 regions of interest were inserted, while 3 regions were located on the ischemia-affected hemisphere capturing (a) the ischemic zone in subcortical regions (i.e., the striatum), (b) the border zone toward the adjacent neocortex, and (c) the parieto-temporal neocortex, and the same 3 regions were mirrored to the contralateral, non-affected hemisphere. Each of the regions of interest represented the full field of an image taken with a 60x objective and a homogeneous exposure time of 130/s, containing both Cy2-stained STL-binding sites and Cy3-immunolabeling of ChAT or TH. Applying the control software BZ II viewer in addition to the software BZ II analyzer (both Keyence), in each of the 6 regions of interest per brain section, staining patterns were analyzed concerning (a) intensity, (b) density, and (c) integrity. While staining intensity and density represented automatically calculated values, integrity was depicted by a manual scale driven by the visual impression, ranging from 1 (high grade of discontinuity) to 3 (nearly no interruption of the fibers). Data were further processed by calculating differences between the ischemia- and the non-affected hemisphere, which resulted in Δ values reflecting quantitative changes on the affected hemisphere in terms of an increase or decrease of assessed parameters. Further, means between the 3 sections analyzed per animal were calculated. Overall, mean values for Δ intensity, Δ density, and Δ integrity resulted for the ischemic zone, the ischemic border zone, and the neocortex for each of the addressed alterations of ChAT, TH, and STL.
Statistical analyses
All calculations were performed with the IBM SPSS Statistics package version 24.0 (IBM Corp., New York, NY, USA). In addition to descriptive statistics, the Wilcoxon test and the Mann-Whitney U test were used to check for statistical significance between groups. Further, Pearson correlations were applied to explore interrelations between different parameters. Data are given as means ± standard deviation, unless otherwise indicated. Generally, a p-value < 0.050 was considered as statistically significant. If applicable, trends were discussed, if the p-value ranged between 0.05 and 0.400.
RESULTS
Alzheimer-like alterations in close association to the neurovascular unit
To verify the existence of Alzheimer-like alterations in the used 3xTg mouse model, two variants of triple fluorescence labeling were applied. First, hippocampi of 2-year-old mice displayed strong immunoreactivity for total Aβ (Fig. 1A), even without formic acid pre-treatment, and subsequently performed thioflavin S (ThS) staining (Fig. 1A’), indicating numerous β-sheet structures. As expected, Aβ immunolabeling and ThS staining revealed largely the same structures predominantly in the core of plaques (Fig. 1A”), whereas their rims were devoid of β-sheet structures, and phospho-tau as detected by biotinylated AT8 and Cy5-streptavidin (Fig. 1A”, color-coded in blue) was found without evidence of direct co-localization. In a second set of experiments also focusing on hippocampi in aged 3xTg mice, staining was performed to address the regional pattern of N-terminally truncated pyroglutamyl-Aβ3–40/42 and AT8-immunopositive phospho-tau in relation to the astroglial component of the NVU. In good accordance to the first approach, phospho-tau and N-terminally truncated Aβ as features of the AD-like background were allocated and surrounded by reactive astrocytes as shown by a strong glial fibrillary acidic protein (GFAP)-immunoreactivity (Fig. 1B).
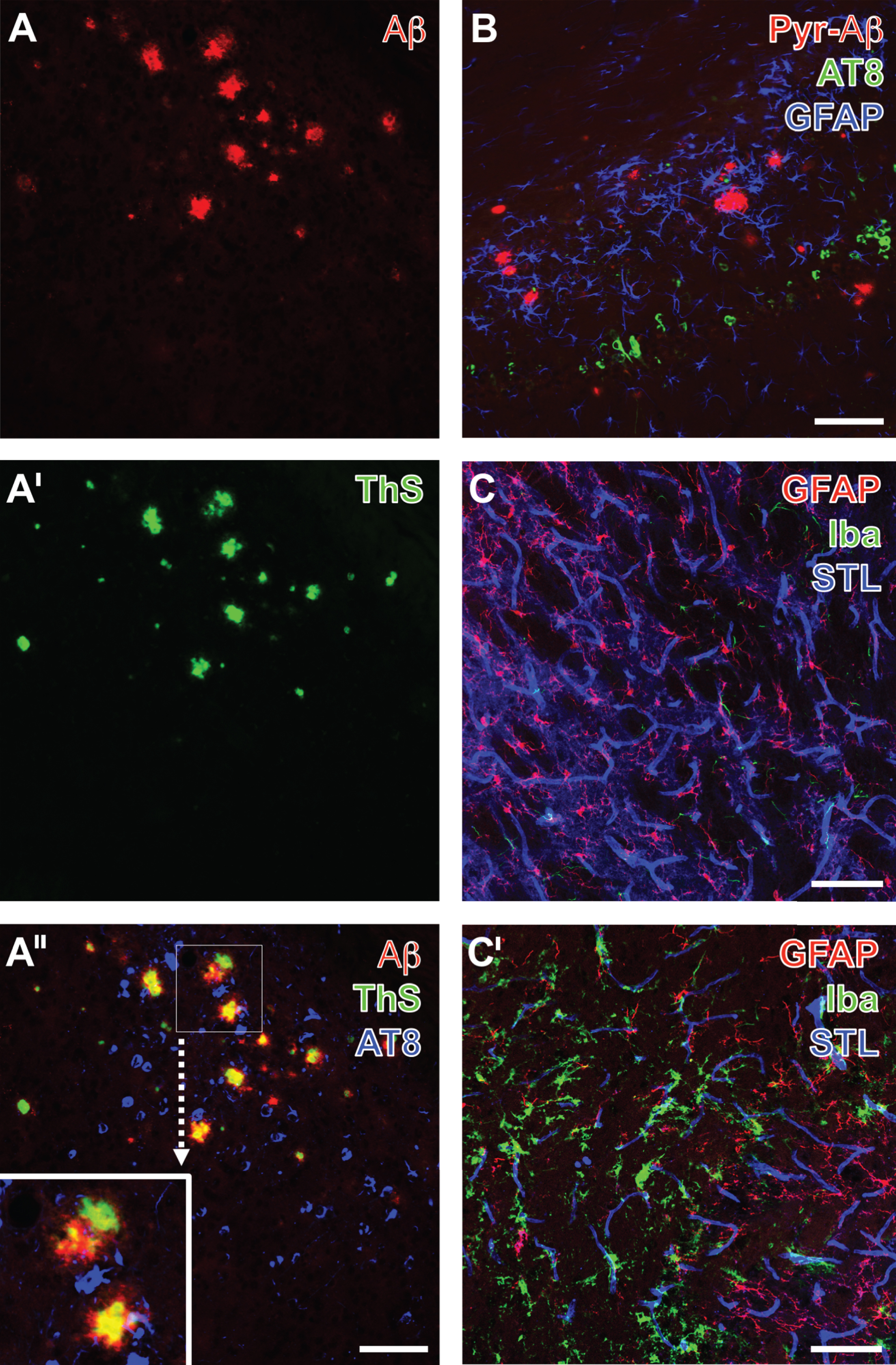
Triple fluorescence labeling of hippocampal age-dependent Alzheimer-like alterations in 2-year-old 3xTg mice (A-A”, B) as well as of striatal vascular and glial alterations one day after focal cerebral ischemia (C,C’), visualized by Keyence and confocal-laser scanning microscopy, respectively. Senile Plaques are revealed by Cy3-immunolabeling of total Aβ (A) and in their cores by thioflavin S (ThS) staining (A’), indicating β-sheet structures. In the merge of staining patterns (A”), additional Cy5-labeling with biotinylated AT8 (color-coded in blue) is found in close vicinity to plaques, but devoid of co-occurring Aβ-immunoreactivity and ThS-binding, which appears yellow in the core of plaques, whereas the red appearing rim of deposits is frequently devoid of β-sheet structures. Further neuropathological alterations are shown in (B) demonstrating the allocation of N-terminally truncated pyroglutamyl-Aβ3–40/42 (Pyr-Aβ, Cy3, red), AT8-immunoreactive phospho-tau (Cy2, green) and GFAP-expressing reactive astrocytes (Cy5, color-coded in blue). Ischemia-induced glial and vascular alterations (C, C’) are representatively captured by concomitant labeling of astroglial GFAP (Cy2, green), microglial Iba (Cy3, red) and endothelial binding sites for Solanum tuberosum lectin (STL, Cy5, color-coded in blue). The contralateral, non-altered side (C) is thereby characterized by a weak GFAP immunosignal, solely ramified microglia, while focal cerebral ischemia causes activation of astrocytes, ameboid microglia and morphological changes of the vasculature visualized by thinned STL-staining (C’). Scale bars: A” (also valid for A,A’), B, C, C’ = 75 μm.
With respect to the concept of the NVU, in 3-month-old wild-type mice a first screening approach for the identification of the ischemia-affected tissue included simultaneous staining of astroglial GFAP and the microglial ionized calcium-binding adapter molecule 1 (Iba) combined with the visualization of STL-binding sites known to be predominantly located in endothelial cells. The striatum on the non-affected hemisphere exhibited astrocytes with fine processes that were embedded in a network of endothelial cells as part of the vasculature, while microglia were only weakly labeled (Fig. 1C). In contrast, regions affected after 24 h of ischemia (on the ipsilateral hemisphere) were characterized by a strong microglial reaction (Fig. 1C’), whereas immunolabeling of the astroglial GFAP became weakened and the endothelial structures as visualized by STL clearly thinned out.
Neuronal subpopulations and fiber characteristics after ischemia
As morphological alterations of the endothelium visualized by STL were found to represent a robust feature of focal cerebral ischemia, double fluorescence labeling of STL and ChAT or TH was conducted to reveal changes of neuronal subpopulations as well as their fibers, representing the striatal catecholaminergic and cholinergic system in wild-type (Fig. 2) and genetically altered animals (Fig. 3).
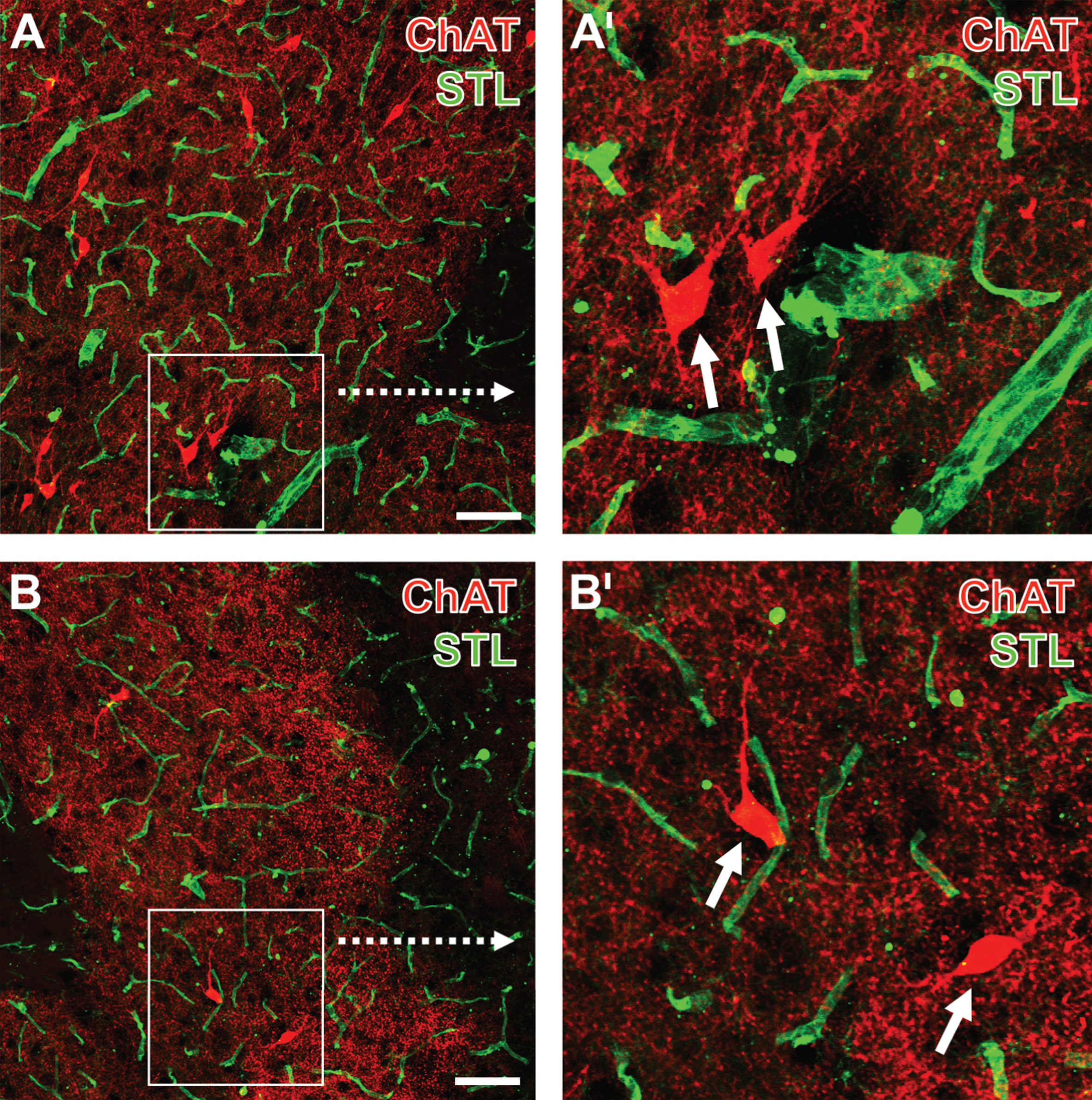
Concomitant fluorescence staining of choline acetyltransferase (ChAT, Cy3, red) and endothelial binding sites for Solanum tuberosum agglutinin (STL, Cy2, green), exemplified in the striatum of a 12-month-old wild-type mouse one day after focal cerebral ischemia, comparing the non-affected contralateral side (A,A’) with the ischemia-affected striatum (B,B’). Lower magnified pictures (A, B) show in addition to perikarya of cholinergic interneurons a dense cholinergic fiber network, frequently in close vicinity to STL-stained endothelial cells. Fibers in the ischemic tissue are found considerably coarser than those in the non-affected hemisphere. Additionally, the STL-stained vessels appear thinned in stroke-affected regions. Higher magnified frames (A’,B’) demonstrate the ischemia-caused alterations of fibers even clearer and reveal details of perikarya (arrows). Scale bars: A and B = 25 μm.

Simultaneous fluorescence labeling of tyrosine hydroxylase (TH, Cy3, red) and endothelial binding sites for Solanum tuberosum agglutinin (STL, Cy2, green), exemplarily captured in the striatum of an aged 3xTg mouse one day after focal cerebral ischemia at lower (A) and higher magnification (B). Notice the close spatial relationships between catecholaminergic (TH-positive) fiber endings and the vasculature. Scale bars: A = 100 μm, B = 25 μm.
Twenty-four hours after ischemia onset, ChAT-immunopositive neuronal structures were identified in terms of a dense network of well delineated fibers and nearly completely visualized perikarya, both in close regional relationship to STL-positive vascular elements in subcortical regions on the non-affected (Fig. 2A,A’) as well as the ischemia-affected hemisphere (Fig. 2B,B’). ChAT-positive elements appeared, at least concerning their processes, slightly rarefied and also decreased regarding the intensity of Cy3-staining. Additional quantitative analyses of ChAT immunolabeling in the overall study sample confirmed this observation by providing a significant decrease of the staining intensity and density in the ischemia-affected striatum compared to the contralateral non-affected side (intensity: 1.63±1.34 versus 2.25±1.64, p = 0.002; density: 3.04±7.39 versus 12.13±13.69, p < 0.001; Wilcoxon test for each). Although to a much lesser degree, a significant decrease of the staining intensity and density was also observed in the border zone of the ischemia-affected hemisphere when compared to the non-affected side (intensity: 2.22±1.95 versus 2.63±2.26, p = 0.008; density: 10.78±12.25 versus 18.00±18.72, p = 0.036; Wilcoxon test for each). However, also in the neocortex, a region not primarily affected by the ischemic model, which causes proximal middle cerebral artery occlusion and thus an ischemic lesion on subcortical areas, a significant decrease of the staining intensity and density was noted when comparing the ischemia-affected hemisphere with the contralateral side (intensity: 2.25±2.19 versus 2.71±2.38, p = 0.008; density: 13.71±16.62 versus 22.71±18.77, p = 0.003; Wilcoxon test for each).
Serial double fluorescence labeling of TH and STL-binding sites revealed strongly stained and clearly delineated catecholaminergic fibers in the ischemia-affected striatum (Fig. 3A). The fibers appeared mostly arranged in a parallel fashion, whereas their endings were found in close vicinity to the endothelium as stained by STL, which became visible at higher magnification (Fig. 3B). Subsequent quantitative analyses addressing inter-hemispheric differences of TH-immunoreactivity in the overall study sample revealed a significant decrease of staining density in the striatum as the primarily affected zone (7.78 ± 10.45 versus 13.41 ± 16.11, p = 0.014; Wilcoxon test), while staining intensity did not differ markedly (5.22 ± 3.81 versus 5.50 ± 4.03, p = 0.271; Wilcoxon test). In the border zone of ischemia, staining intensity was not altered by the ischemic stimulus when compared to the contralateral side (5.45 ± 3.97 versus 5.59 ± 3.97, p = 0.512; Wilcoxon test), whereas staining density decreased due to ischemia (12.78 ± 19.02 versus 18.95 ± 22.57, p = 0.002; Wilcoxon test). However, in the neocortex neither staining intensity nor density differed significantly between the ischemia-affected and the non-affected hemisphere (intensity: 4.54 ± 3.61 versus 4.73 ± 3.32, p = 0.417; density: 7.64 ± 15.50 versus 11.09 ± 21.57, p = 0.549; Wilcoxon test for each).
Neurovascular changes after ischemia
To further explore ischemia-related alterations of cholinergic and catecholaminergic neuronal structures and the vasculature, we performed triple fluorescence labeling of ChAT, TH, and STL-binding sites. Thereby, the striatum on the non-affected hemisphere exhibited ChAT-immunopositive cellular bodies with adjacent processes in addition to a well-arranged network of fine ChAT- and TH-positive fibers (Fig. 4A,A’). These structures were found to be embedded in a network of STL-positive endothelia (Fig. 4A”,A”’), characterized by frequently apparent lumina and a continuous surface. In contrast, the ischemia-affected striatum exhibited a numerous decrease of ChAT-immunopositive cellular bodies and their processes as well as a degraded appearance of the ChAT-positive fiber network (Fig. 4B). However, while the TH-positive network appeared seriously diminished in the directly ischemia-affected region (right part of the figure), the neighboring parts displayed a slightly increased density of coarser fibers (Fig. 4B’). The merge of staining patterns clearly indicated concomitant changes of the two addressed neuronal components (predominantly characterized by signs of degradation) and the STL-positive endothelium, which appeared significantly decreased concerning the number of endothelial cells and the diameter of definable vessels (Fig. 4B”’).
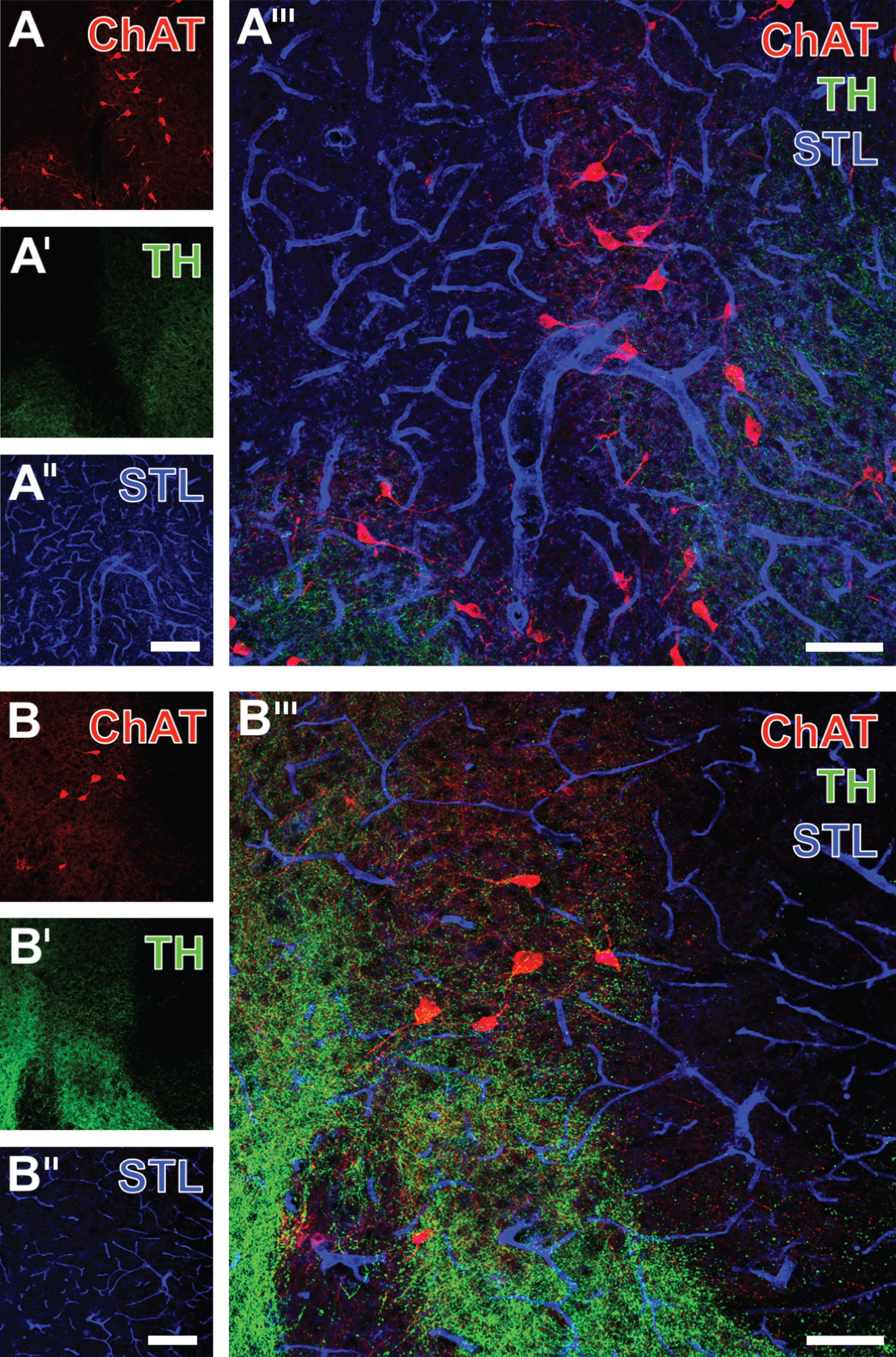
Triple fluorescence labeling of ChAT, TH and STL-binding sites one day after focal cerebral ischemia, exemplarily shown in the striatum of a 3-month-old wild-type mouse, comparing the non-affected (A-A”’) with the ischemia-affected side (B-B”’). Thereby, Cy3-immunolabeled cholinergic cells appear less numerous and display a reduced staining intensity due to ischemia (A,B). Concomitantly, Cy2-immunostaining reveals diminished TH-positive fibers in the severely ischemia-affected region (right part of the figure), while in the neighboring region both a degradation and at least partially increased fiber density is noted (A’,B’). Additionally visualized STL-binding sites of endothelia appear somewhat thicker in the non-affected (A”) versus the ischemic tissue comprising the same neuroanatomical structures ipsilaterally (B”). Notably, the comparison between merged staining patterns (A”’, B”’) clearly demonstrates altered TH-stained fiber morphology and numerical reduction of ChAT-positive cell bodies due to focal cerebral ischemia. Scale bars: A” and B” (also applicable to A and A’ as well as B and B’) = 150 μm, A”’ and B”’ = 75 μm.
With the intention to perform a numerical investigation of apparently simultaneously occurring changes in the neuronal and the vascular component of the NVU, correlations between histochemical findings were calculated in the overall study sample, regardless of age and genetic background. Thereby, inter-hemispheric differences (Δ) of ChAT, which were larger than those of TH (see previous paragraph), and STL staining intensity (Fig. 5A) and density (Fig. 5B) were used. Notably, most correlation coefficients were characterized by a consistently positive sign, indicating the same direction of changes, which means that the diminished ChAT-immunoreactivity due to ischemia corresponds with the affection of endothelia. However, considerable interactions were limited to the staining intensity of ChAT in the ischemic zone (p = 0.018; Pearson correlation) and in the neocortex (p = 0.001; Pearson correlation). Further, changes of staining intensity in the border zone (p = 0.187; Pearson correlation) as well as staining density in the bordering zone (p = 0.340; Pearson correlation) and the neocortex (p = 0.114; Pearson correlation) remained as a non-significant trend, while staining density in the ischemic zone (p = 0.956; Pearson correlation) clearly failed statistical significance and thus an interpretable relationship between both markers.
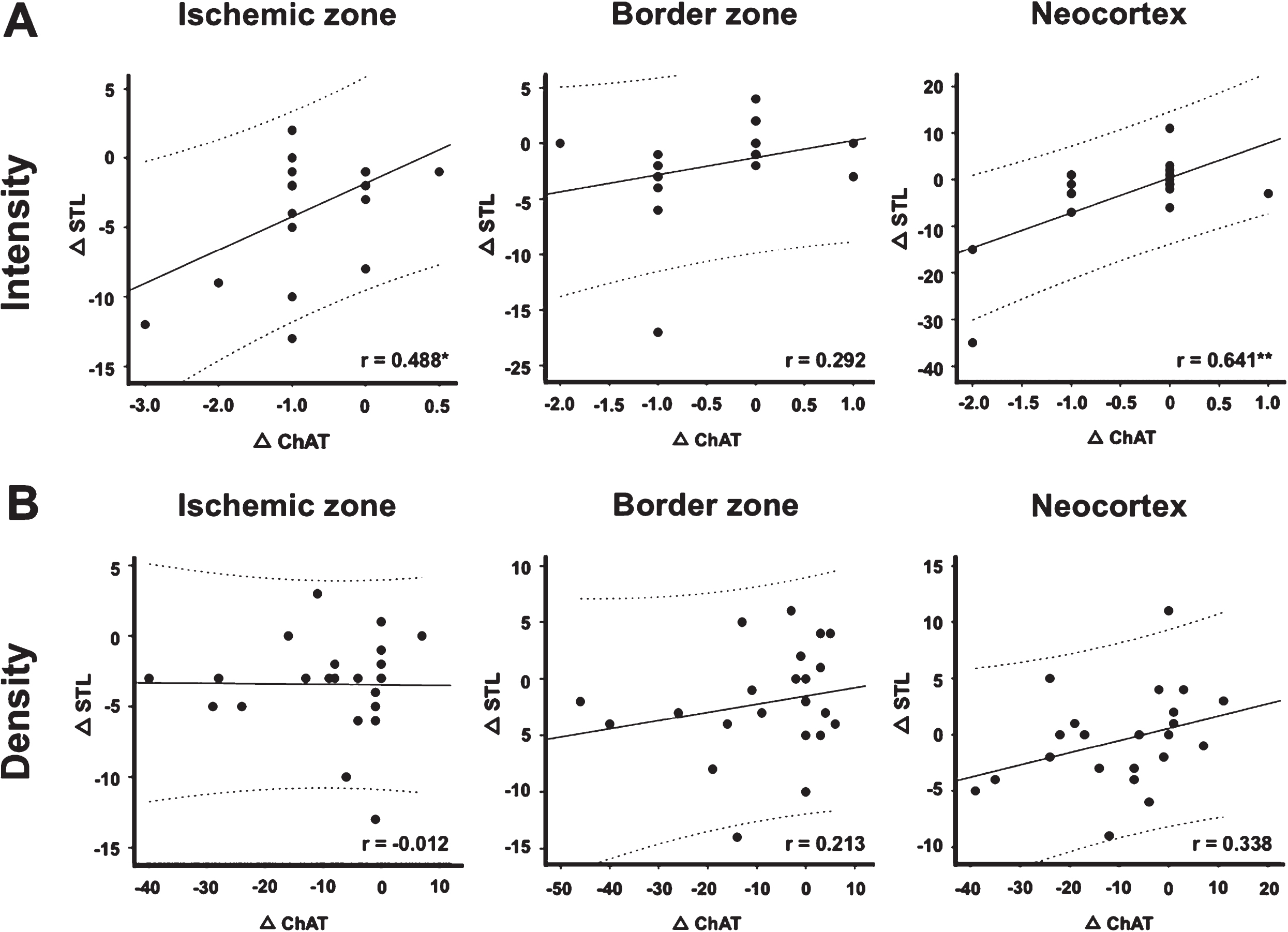
Scatter plots from the overall study sample, visualizing the relationship between inter-hemispheric changes of Solanum tuberosum agglutinin-binding sites (Δ STL) and choline acetyltransferase immunostaining (Δ ChAT) in different brain regions 24 hours after unilateral focal cerebral ischemia. A significant statistical relationship is found between the vascular and the neuronal component in the ischemic zone and the neocortex. Note the positive sign of the coefficient, indicating alterations of the same direction, which means that a decrease in ChAT-immunoreactivity corresponds with a decrease in STL-staining and vice versa. r, Pearson correlation coefficient; * p < 0.05, ** p < 0.01. Scatter plot and respective calculation based on n = 22 to n = 23 animals.
Since ChAT- and TH-immunopositive fibers were shown to form, at least in part, parallel tracts in the vicinity of the vasculature and appear to build contacts with the endothelium (see Figs. 2 and 3), a subset of experiments focused on the neurovascular coupling, i.e., the relation of fiber endings to the surface of vessels. Therefore, triple fluorescence labeling was performed to visualize ChAT (Fig. 6A) and TH (Fig. 6B) in the ischemia-affected striatum together with STL (Fig. 6A”,B”) and collagen IV (Fig. 6A’,B’), known as a main component of basal membranes. This approach showed the close regional association of addressed neuronal elements and the vasculature. Although the merge of staining patterns (Fig. 6A”’, B”’) elucidates the association of basal membrane constituents and the endothelium, a certain coupling of ChAT- or TH-positive fibers could be detected.
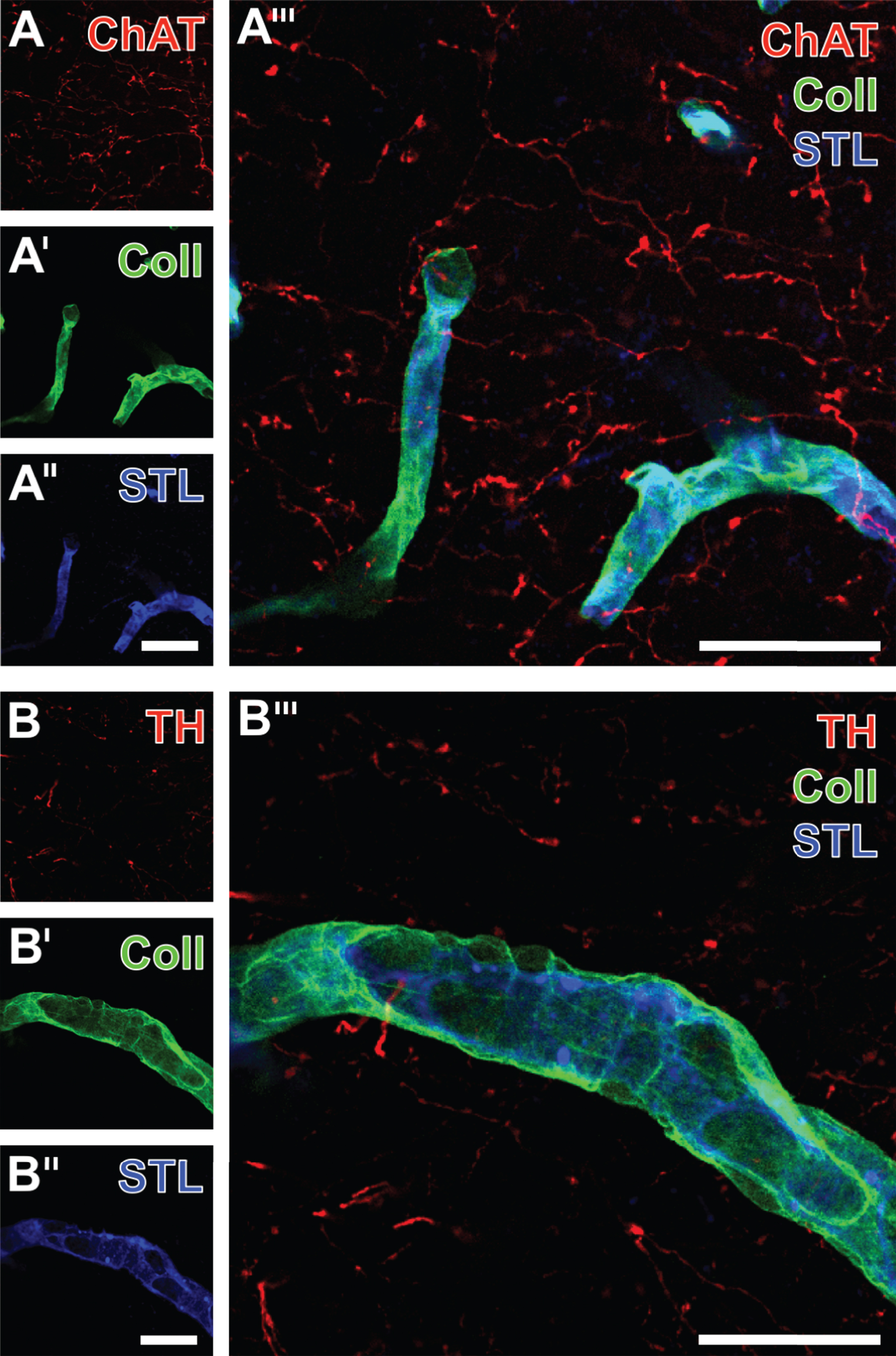
Vascular double staining of collagen IV (Coll) in basal membranes and associated STL-labeled endothelial cells combined with the detection of fibers expressing ChAT (A) or TH (B). Confocal laser scanning micrographs, exemplarily taken from an aged 3xTg mouse, visualize at lower magnification (A-A”’) and higher magnified (B-B”’) the neurovascular arrangement in the ischemia-affected striatum. The overlay at higher magnification (B”’) clearly elucidates how the endothelia are enwrapped by Coll-marked basal membranes toward the neuropil. Scale bars: A” and B” (also applicable to A and A’ as well as B and B’) = 25 μm, A”’ and B”’ = 25 μm.
Ischemia-induced neurovascular alterations depending on age and genetic background
As immunoreactivities for ChAT and TH were found to decrease due to the ischemic stimulus, subsequent analyses focused on age and the genetic background as potential influencing factors. For this purpose, differences between the ischemia-affected and non-affected hemisphere (Δ intensity, Δ density, and Δ integrity) were compared between WT and 3xTg mice at an age of 3 as well as 12 months (Fig. 7).
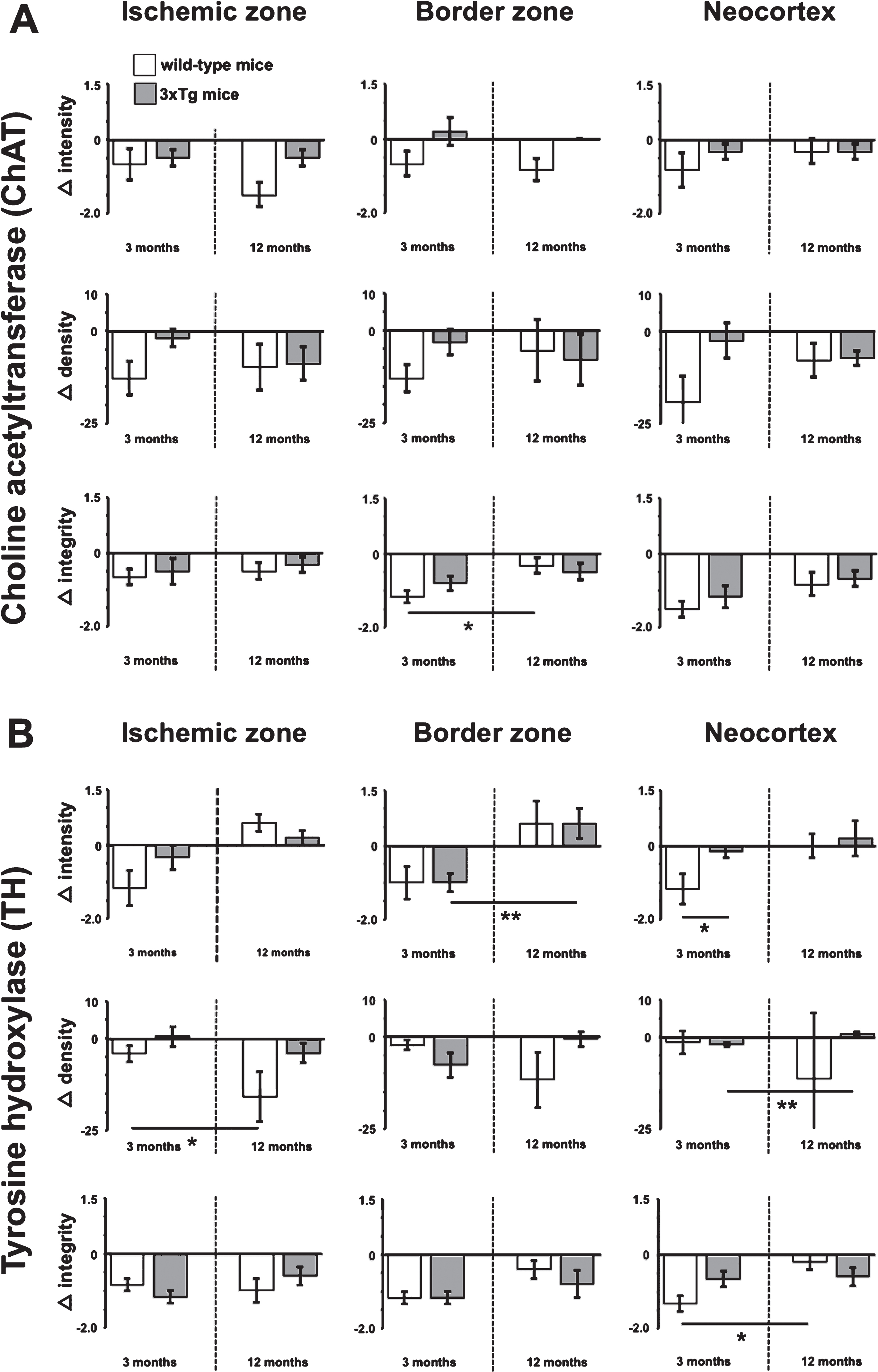
Immunofluorescence-based quantified inter-hemispheric changes of choline acetyltransferase (Δ ChAT) and tyrosine hydroxylase (Δ TH) in diverse brain regions, captured by the staining intensity and density as well as fiber integrity (ranging from 1 [high grade of discontinuity] to 3 [no fiber interruption]), and with reference to age (3 versus 12 months) and the genetic background (wild-type versus 3xTg) one day after unilateral focal cerebral ischemia. The Mann-Whitney U test is used to check for statistical significance. * p < 0.05, ** p < 0.01. Descriptive analyses and statistical calculations are based on n = 5 to n = 6 animals per group. White and grey bars represent mean values, while added lines represent respective standard errors.
In 3-month-old animals, ChAT-immunolabeling appeared was less dense and weaker in wild-type mice when compared to animals exhibiting the transgenic background. Such data were obtained for the ischemic zone, the border zone, and the neocortex (Fig. 7A), but, however, failed statistical significance. In detail, considerable trends were noticed for staining intensity in the bordering zone (p = 0.126) and the neocortex (p = 0.394) as well as staining density in the ischemic zone (p = 0.093), the bordering zone (p = 0.126), and the neocortex (p = 0.132; Mann-Whitney U test each). However, 12-month-old animals displayed remarkable differences depending from the genetic background with regard to staining intensity in the ischemic zone and the border zone, which closely failed statistical significance (p = 0.065 each, other p-values ranging between 0.485 and 1.000; Mann-Whitney U test each). Analyses concerning the integrity of ChAT-immunopositive fibers revealed a homogeneous pattern between the genetic background in both 3- and 12-month-old mice. When focusing on potential age-related effects, a significant difference in fiber integrity toward a higher grade of discontinuity was seen in 3-month-old wild-type mice as compared with the older animals (p = 0.041; Mann-Whitney U test).
Immunoreactivity for TH in relation to age and the genetic background revealed more inhomogeneous patterns than ChAT (Fig. 7B). Remarkably, animals of 3 months of age exhibited a significant decrease of TH-immunoreactivity in the neocortices from the wild-type group when compared to animals with AD-like background (p = 0.041; Mann-Whitney U test). Additionally, staining intensity in the ischemic zone in 3-month-old mice revealed a trend toward a more severe affection in wild-type animals when compared with 3xTg mice (p = 0.180; Mann-Whitney U test). However, in animals of 12 months of age, TH staining intensity in the ischemic zone appeared only slightly affected and in an inverse manner, while 3xTg mice tended to be more affected than the wild-type group (p = 0.310; Mann-Whitney U test). Analyses focused on age-related effects revealed a stronger decrease of TH staining density in the ischemic zone for 12-month-old mice compared to the younger animals (p = 0.017; Mann-Whitney U test). In contrast, 3-month-old 3xTg mice exhibited a pronounced decrease of TH staining intensity in the border zone and TH staining density in the neocortex (p = 0.009 each; Mann-Whitney U test each). Regarding changes on integrity of TH-immunopositive fibers, a significant difference was only found in the neocortex of wild-type mice, while 3-month-old mice showed more diminished values indicating an increase of discontinuity when compared to older animals (p = 0.017; Mann-Whitney U test each).
DISCUSSION
The present study aims to explore neuronal changes with emphasis on the cholinergic system in conjunction with vascular and glial alterations in the AD-like brain affected by permanent focal cerebral ischemia. As earlier studies mainly addressed ischemic consequences with regard to typical AD features like tau hyperphosphorylation (e.g., [16]), the focus of the present study was changed toward simultaneous histopathological alterations of the NVU complex. Strictly following the concept of the NVU, this study therefore provides novel insights into the relationship between neuronal subpopulations and the vasculature in the AD-like brain. To address potential age-related effects as a typical feature of AD, analyses covered 3- and 12-month-old mice with an either transgenic or wild-typebackground.
Among the variety of animal models in the field of AD research [52, 53], we used a triple-transgenic mouse model with age-dependent deposition of Aβ and tau hyperphosphorylation [46, 47]. To verify the study setup, i.e., the existence of AD characteristics together with ischemia-affected constituents of the NVU, multiple fluorescence labeling was performed visualizing phospho-tau and Aβ in terms of total Aβ and the N-terminally truncated pyroglutamyl-Aβ3–40/42 concomitantly with a morphologically altered vasculature and astro- and microglia.
Vascular and glial changes in the AD-like brain
Unilateral focal cerebral ischemia for 24 h was associated with reactive astrocytes and a strong microglial reaction, characterized by morphological changes in terms of diminished cellular integrity and numerically reduced astrocytic processes and less ramified microglia with swollen cell bodies. These findings are in line with reported cellular changes for both the AD background itself [54, 55] and following ischemia [39]. Albeit in our previous report, STL as a vascular marker did not reveal a hemispheric difference on a statistically significant level when captured as stained area per region of interest [39], the current experiments robustly demonstrated morphological changes on the level of single vessels in terms of a thinned appearance in ischemia-affected regions, which allowed allocation analyses with neuronal components of the here addressed NVU.
In general, changes in vascular morphology and functionality have extensively been studied in the field of stroke, presumably related to the fact that focal cerebral ischemia is caused by acute vessel occlusion, which has very early directed therapeutic efforts towards re-opening approaches [56 –59]. In association with age-related, i.e., neurodegenerative pathologies, the term NVU was first used following the perspective that features of the aged brain like deposition of Aβ and long-term consequences of cardiovascular risk factors (e.g., diabetes and hypertension) influence the vasculature and related glial cells [30 , 60]. For example, in a model of chronic hypoperfusion due to bilateral carotid artery stenosis in mice, a significantly reduced blood flow in deep cortical capillaries and an increased adhesion of leukocytes in pial vessels was observed by using in vivo real-time imaging [61]. Further, a study on 19 patients with cerebral amyloid angiopathy provided evidence for a significant impairment of the blood-brain barrier integrity as addressed by magnetic resonance imaging and serum levels of tight junction proteins and matrixmetalloproteinases [62].
More focused on the setting of AD, changes on the vasculature with associated blood-brain barrier breakdown were consistently described in disease-related human and animal studies, leading to the hypothesis that at least some of the AD-specific pathophysiological findings might be due to alterations of the vascular integrity [34, 63]. A current hypothesis of AD goes one step further and discusses a failure for the drainage of perivascular Aβ as a pivotal element for disease progression [64, 65]. Although this view is still a matter of debate, its existence clearly illustrates the emerging thoughts on a bidirectional causal relationship between neuronal features seen in AD and concomitant changes of the vasculature, in particular of the blood-brain barrier.
Morphological changes of astrocytes in the AD-like brain were described in numerous experimental studies, while astrogliosis is widely accepted as a reactive phenomenon caused by the AD-related pathophysiology [55, 66]. This perspective is largely based on the observation that astroglial changes occur in an age-dependent fashion concomitantly to the AD-like pathology [47]. However, recent reviews [67, 68] clearly support the view that disease-related neuropathological alterations including those during AD might be a consequence of astroglial dysfunction, which would turn the traditional perspective.
Microglia as a population of predominantly resident, immunologically active cells were comprehensively investigated in the healthy brain as well as under various neurodegenerative and ischemic conditions [69]. In the field of AD, very early morphological alterations on microglial cells were described [70], while a causal relationship was postulated in terms of a cellular activation due to AD featured like the deposition of Aβ [71]. However, in a recent study dealing with mouse models of stroke and AD, a subpopulation of microglia was identified to exhibit muscarinic acetylcholine receptors [72]. This finding might be relevant since the activity of ChAT, the marker enzyme for cholinergic neurons, was found to correlate with the amount of cognitive impairment in humans [73]. Further, as the cholinergic system is characterized by a widespread network between subcortical regions such as the basal forebrain, and the neocortex [74 –76], its drastic involvement in AD was also discussed to be causally linked to other AD characteristics, i.e., hyperphosphorylation of tau and Aβ deposition [43, 49]. Summarizing the last two decades, alterations of the vasculature, the micro- and astroglia along with the clinical phenotype and histopathological hallmarks of AD have gained increasing attention, accompanied with a shift from the traditional unilateral view for causal relationships toward a bidirectional model describing complex interactions between diverse cell types within the NVU [32, 35]. The emerging perspective of multiple interplays in the NVU with assumed consequences on disease progression or at least maintenance of AD may also improve treatment strategies [77].
Neurovascular alterations due to ischemia
Cholinergic neuronal structures, i.e., mainly fibers and cell bodies, as identified by ChAT-immunoreactivity were found in close vicinity to the vasculature indicating that both structures are crucial parts of the NVU in the AD-like brain. Qualitative and added quantitative analyses revealed an impairment of ChAT-immunopositive structures as both staining intensity and density were significantly decreased due to focal cerebral ischemia. Remarkably, diminished staining intensity and density were also noted in the ischemic border zone and in the ipsilateral neocortex as a region with some distance to the origin of the middle cerebral artery, which is getting closed in the applied ischemia model. Given the fact that addressing ChAT-immunoreactivity represents a robust technique to visualize cholinergic neurons [49], the current findings entailed two implications: First, the far-reaching affection beyond the striatum as the primary ischemic region strengthened the already established view of a widespread network of cholinergic neurons involving neocortical and subcortical regions [43, 74]. Second, as the performed analyses were corrected for the contralateral, non-affected hemisphere, the found decrease in ChAT-immunoreactivity could be seen as a result of the ischemic stimulus that lasted 24 h prior to tissue fixation. In the synopsis with previously reported vascular changes in terms of a collagen IV upregulation 24 h after focal cerebral ischemia [39], the present data illustrated the manifold reactions within the NVU in a relatively short duration, i.e., one day, of an ischemic stimulus. Thereby, it is likely that in a longer observation period an aggravation of the existing AD-like histological features would also be detectable, since earlier reports from Koike et al. [16] and Majd et al. [78] found altered tau levels up to 3 months after cerebral ischemia. Regarding cognitive consequences, although not assessed in the present study, a considerable impact might be adopted from two recent studies [79, 80]: Combined Aβ toxicity (by intraventricular infusion) and endothelin-1-induced ischemia resulted in impaired memory function, even if the striatal ChAT-staining only tended to decrease. Comparable data were provided from Kemppainen and colleagues [21], who demonstrated robust impairment on spatial memory function during the first 5 days after transient global cerebral ischemia in mice exhibiting an AD-like background.
The present study also provides evidence for a crucial impact of the catecholaminergic system in the ischemia-affected NVU of the AD-like brain, since TH-positive structures were found to build fiber-like structured in close vicinity to the vasculature in ischemia-affected regions. The conducted quantitative analyses revealed a significantly decreased staining density of TH-immunopositive fibers in the ischemic striatum and in the border zone. However, TH-immunoreactivity failed to differ significantly between the ischemia-affected and the non-affected hemisphere, suggesting a regional limitation of ischemic pathways involving the catecholaminergic system, at least when focusing on its marker enzyme TH.
In order to explore concomitant immunohistochemical characteristics of the ChAT-immunopositive neuronal structures and the vasculature, scatter plots capturing the inter-hemispheric differences were used to visualize possible relationships between the neuronal and the vascular compartment, added by calculated correlations. Thereby, a statistically significant relationship was identified regarding the changed staining intensity within the ischemia-affected striatum and the ipsilateral neocortex. Given the positive sign of the respective correlation coefficient, the ischemia-related declined ChAT-immunoreactivity corresponds with diminished STL-labeling, which indicates alterations in the same direction with a considerable explained variance. However, the causal relationships remain to be elucidated, while both AD-specific or age-related features might explain the vascular reaction in the setting of the ischemic stimulus. Concerning the latter one, a study on non-transgenic mice of different ages and with chronic cerebral hypoperfusion revealed a significantly impaired cerebrovascular reactivity and collateral remodeling in animals with 18 months of age when compared to younger groups (12 and 4 to 6 weeks of age) [81]. These data strongly indicate that age is the main factor for the increased susceptibility of the vasculature against ischemic events. However, as bidirectional causal relationships between ischemic events and the histopathological phenotype of AD have been discussed extensively during the last decade [25], various reasons for morphological and functional changes of the vasculature in AD can be assumed.
To further explore potential age-dependent effects, analyses were extended by considering subgroups of wild-type and 3xTg mice of either 3 months or 12 months of age. Throughout the investigated regions, a trend was found toward a more severe, ischemia-caused affection of ChAT-immunoreactivity in wild-type than in 3xTg mice of 3 months of age. However, in 12-month-old animals, this trend remained limited to the ischemia-affected striatum and the border zone. Notably, a significant difference in the integrity of ChAT-positive fibers was observed in the border zone when asking for age-related effects, while wild-type mice of 3 months of age exhibited fibers with more severe discontinuity. A similar effect was noted for the integrity of TH-positive fibers in the neocortex, also pronouncing the younger animals. For TH-immunoreactivity, significant differences between the genetic background were also observed in the neocortex, while wild-type mice here showed a more severe decline of staining intensity compared with the genetically altered group. However, the observed inhomogeneously altered intensity and density of TH-immunoreactivity might be related to the noted TH staining pattern, i.e., its fiber-like appearance and frequent coarse puncta as well as the lacking cell bodies. It remains to be elucidated if these different morphologies and the observed age-related effect of a slightly enhanced immunolabeling of TH in the bordering zone of 12-months-old animals may represent part of a compensatory mechanism due to a decreased activity in the ischemic striatum. In summary, age- and phenotype-related effects were found for both ChAT- and TH-immunoreactivity. As our previous work, dealing with vascular changes after focal cerebral ischemia, identified an age- and phenotype-dependent upregulation of collagen IV-immunoreactivity as part of the vasculature [39], it is likely that several NVU components hold the capacity for age-dependent reactions against ischemic stimuli. This perspective is based on the alterations of the NVU under primary ischemic conditions [30] and the typical age-dependent features of AD[32 , 48].
Conceptional considerations
The present study has some limitations. First, by investigating 3xTg mice, a model of AD was chosen that appears well characterized and translational oriented as it harbors the majority of AD-specific hallmarks [46 , 82]. Studies using other animal models should also involve screening methods to explore neuronal subpopulations and NVU components to either support or revise the findings of the present study. Second, as neuronal constituents within the NVU, exclusively ChAT- and TH-positive subpopulations were chosen as the targeted enzymes critically involved in cholinergic and catecholaminergic pathways. However, from the variety of neuronal subpopulations, future studies should extend the neuronal spectrum, e.g., by addressing GABAergic (inter-)neurons, which are known to have pivotal roles in neostriatal circuits [83]. Third, analyses were limited to a single time point, i.e. 24 h after ischemia onset, and a permanent scenario of focal cerebral ischemia. With respect to translational aspects, future studies are needed to address potential time-depending effects, which are known for post-ischemic reactions [56], and a reperfusion scenario to mimic already existing treatment approaches including intravenous thrombolysis and mechanical recanalization in stroke [58, 59]. Considering the relatively small group size in this study, which is due to the underlying explorative design, future work should analyze more subjects per group to avoid overlooking weak but potentially relevant neurovascular changes. From this perspective, subsequent multicenter studies appear appropriate to achieve sufficient group sizes especially when focusing on pharmacological interventions [84].
Therapeutic implications
Among the changes presented here in association with 24 h of focal cerebral ischemia, the significant decline of ChAT-immunoreactivity together with alterations on vascular and glial components of the NVU appears important for therapeutic considerations. Of note, the traditional perspective of the cholinergic dysfunction in terms of a solely underlying feature for cognitive decline has shifted to a more complex view of bidirectional causal relationships including the deposition of Aβ and hyperphosphorylation of tau [41 , 85]. This view is supported by experimental data demonstrating that an immunolesion-induced impairment of cholinergic projection neurons in the mouse basal forebrain can induce typical AD features, i.e., the accumulation of Aβ peptides [49]. Consequently, Ahmad and colleagues considered the assumed impact of the cholinergic system on AD progression and applied a rat model of transient focal cerebral ischemia, which caused cholinergic dysfunction in the striatum [86]. Experimental treatment with edaravone, known to act as a scavenger for ischemia-caused free radicals, was able to ameliorate the decreasing effect of ischemia on ChAT, and in parallel, provided neuronal survival. Confirming this approach, a recent study by Jiao et al. demonstrated that experimental treatment with edaravone in mice with an AD background leads to a reduction of Aβ deposition and can ameliorate other hallmarks of AD [87]. Although these data are very promising, the next step would be a clinical trial to reevaluate this approachin humans.
Derived from the epidemiologically identified, common risk factors of stroke and AD, i.e., arterial hypertension, hypercholesterolemia, and diabetes, with known detrimental impact on the vasculature, a treatment approach for AD might involve drugs usually applied in cardiovascular medicine [88]. This view is supported by a recent study on transgenic animals harboring an AD-like background that underwent initiation of chronic hypertension by adding Nω-Nitro-L-arginine methyl ester hydrochloride to the drinking water, which resulted in a significant increase in arterial blood pressure [89]. Remarkably, in this study chronic hypertension caused microvascular deposition on Aβ, accompanied with an acceleration of cognitive deficits as well as a decrease in blood-brain barrier integrity and vascular inflammation. Conversely, experimental treatment with telmisartan, a drug known to interact with the angiotensin II receptor, was found to decrease the amount of tau and Aβ after long term intake in spontaneous hypertensive rats [90]. Albeit the use of angiotensin II receptor blockers might represent a promising strategy to counteract the pathophysiological hallmarks of AD [91], clinical trials are still missing at this time and need to be performed before a recommendation can be given.
As a further, not yet addressed approach in the field of AD research, pharmacological substances known to provide cellular stabilization or at least maintenance of cellular integrity might be tested in experimental studies. The underlying hypothesis that stabilizing effects within the NVU by well-balanced interactions between neurons, the vasculature, and glial cells might decrease or at least attenuate pathophysiological hallmarks on the cellular level is strongly supported by the findings of the present study.
Conclusion
This study provides novel evidence for a variety of cellular reactions within the NVU of the AD-like brain in the setting of experimental stroke. Along with morphological changes of the vasculature and a strong glial reaction, ChAT and TH as neuronal constituents of the NVU were found to decrease drastically due to 24 h of experimental focal cerebral ischemia. Additionally, statistical analyses revealed commutated degeneration of the cholinergic system (ChAT) and the endothelium, indicating simultaneous changes of different cell types within the NVU from ischemia-affected brains with an AD-like phenotype. Further, a trend was noted toward a more severe involvement of cholinergic components in younger animals. These findings strengthened current efforts on a pharmacological modulation of neuronal subpopulations such as cholinergic cells, and emphasized the complex interplay of cell types within the NVU. The latter one implicates that novel treatment strategies should consider cellular changes beyond neuronal degradation and tau hyperphosphorylation, e.g., by approaches on stabilizing effects toward an enhanced integrity of the endothelium and glial elements, which might be accompanied by an alleviated disease progression.
Footnotes
ACKNOWLEDGMENTS
The authors would like to thank Petra Fink-Sterba and Sigrid Weisheit (Medizinisch-Experimentelles Zentrum, University of Leipzig) for animal care and breeding. Initial microscopic analyses were kindly supported by Cheryl A. Hawkes and Roxana O. Carare (University of Southampton, UK). Carsten Hobohm (Department of Neurology, University of Leipzig) kindly provided initial financial support. The authors would also like to thank Joseph Claßen (Department of Neurology, University of Leipzig) for critical revision of an earlier manuscript version.