
Abstract
Epidemiological studies state that dementia has multiple etiologies including genetic mutation, genetic variation, and environmental factors. Accumulating evidence suggests that dysregulation of cholesterol homeostasis is the major etiological factor in initiating neurodegeneration. Apolipoprotein E (APOE) polymorphic alleles and associated polymorphism of lipoprotein lipase (LPL) and cholesteryl ester transfer protein (CETP) that are important components in regulating cholesterol metabolism are implicated in neurodegenerative diseases. Therefore, the current study focused on identifying the association between several common polymorphism (viz., APOE, CETP, and LPL) to that of change in serum lipid levels and memory symptoms. Volunteer subjects aged 50 and above from rural and tribal areas of the Dharmapuri district, Tamilnadu, India were chosen for the current study and polymorphism was analyzed using PCR-RFLP. Fasting lipid profile and memory function using simplified version of Global Clinical Dementia rating were assessed. Significant difference in the major lipid profile parameters were observed (TC, TGL, LDL, VLDL) among rural and tribal populations that were associated with significant genotypic variation of APOE, CETP, and LPL. Regression analysis revealed significant risk for memory loss that are dependent on age and genetic variants like CETP. These data predict positive correlation between cholesterol-associated genes and their relationship to altered lipid profile and memory symptoms, which possibly link gene-polymorphism and susceptibility ratio for aging and dementia.
Keywords
INTRODUCTION
Dementia is characterized by a progressive decline in cognitive function due to damage to the brain and is different from the slowing of cognitive function that is expected with normal aging [1]. Aging has been considered as the major risk factor for dementia and Alzheimer’s disease (AD) [2]. With the exception of age and apolipoprotein E (APOE) ɛ4 allele, the only disputed genetic component identified [1, 2], environmental risk profiles like smoking, alcohol, obesity, dyslipidemia, diabetes, and cardiovascular diseases may also be the candidates for a dementia condition [3–5]. In developing countries like India, the elderly population is expected to increase 12% of the total population by 2025, making them prone to age-associated inabilities like dementia disorders. As the average life expectancy increases, the number of AD patients are projected to increase 27% by 2020, 30% by 2030, and nearly 70% by 2050, unless there are better treatments for the reduction in the progress of the disease [4].
Accumulated evidence has shown that dysfunction of cholesterol metabolism may contribute to cognitive decline among the aged [6]. Studies on animal and cellular studies have also suggested that abnormalities in cholesterol metabolism are important in the pathogenesis of AD, potentially by increasing neuronal content of cholesterol, promoting cleavage of amyloid-β protein precursor, and impaired working memory [7–10]. Defects in genes regulating the metabolism of cholesterol or lipoprotein have been implicated in the pathogenesis of many age-related diseases including dementia.
The APOE gene polymorphism has been proposed as an important risk factor for neurological disabilities associated with aging [5] and is a key genetic susceptibility factor for AD in numerous ethnic populations [11]. APOE gene exhibits three polymorphic alleles—ɛ2, ɛ3, and ɛ4—which have a worldwide frequency of 8.4%, 77.9%, and 13.7%, respectively [12, 13]. The APOE ɛ4 gene is an only confirmed genetic risk factor for the development of late-onset Alzheimer’s disease (LOAD) [14] and is typically present in >50% of AD patients. It can also be found in about 15% of healthy older controls [13, 16]. It is a high-density lipoprotein that is primarily responsible for lipid transport and cholesterol homeostasis in the central nervous system (CNS). In addition, it is involved in redistributing cholesterol and phospholipids for membrane repair and remodeling [17], and therefore suggested that the alleles of APOE could modulate AD risk through their effects on cholesterol homeostasis.
Apolipoprotein and associated genes of cholesterol metabolism, viz., lipoprotein lipase (LPL) and cholesteryl ester transfer protein (CETP), are implicated in a wide range of neurodegenerative diseases. APOE and LPL polymorphism alter AD risk and indeed LPL is found to present in amyloid plaques [18]. LPL mRNA levels and the highest activity of this enzyme is found in the hippocampus, the region most affected in AD [7, 8]. Corroborating the above statements, polymorphisms in the LPL gene might modify the risk of AD; however, more research is required to understand its relevance to memory function.
CETP regulates cholesterol homeostasis via the transfer of cholesteryl esters from high-density lipoprotein (HDL) to low-density lipoprotein (LDL) in exchange for triacylglycerol (TG) [9, 19]. Clinical and experimental evidence on CETP over the impact of HDL on cognition during aging and in neurodegenerative disorders has been well described and also reported the potential of HDL enhancing approaches to improve cognitive function [20]. CETP gene is a candidate susceptibility gene for LOAD [21] and its polymorphism has been associated with decreased CETP mass and an increase in HDL-cholesterol. The TaqI polymorphism of CETP has been associated with lower cardiovascular risks, but its association with memory decline and dementia risk are unclear. Accordingly, the epistatic effect of these gene variants on the risk profile associated with dementia symptoms must be elucidated.
Our present study attempted to focus on the polymorphic distribution of two varied populations, Palacode (rural) and Pappireddipatti (tribal) from the Dharmapuri district (India) that was chosen based on their lifestyle. Both populations are exposed to varied culture and influenced by different environmental factors. One set of the population is from rural area, where the likely awareness on globalization is prevalent and people around in this area are exposed to wide range of food products, education, communication across the globe, and advancements to some extent. Whereas the other population from tribal group do not have any exposure to globalization nor their lifestyle does not have any variation from their ancestors. Dharmapuri district is one of the reserved places for tribal living and also urbanization. These two subsets of Dharmapuri district differ based on their community and cultural activities. It was observed that the tribal population lives longer with lower incidence of memory symptoms.
Since lifestyle is considered as one of the major associated factors on dementia related complications, and further lack of report on gene polymorphism data among Tamilnadu population, it provoked us to make an attempt to record genetic variations among the population with varied culture and food habits. Also, the present study focus on to determine the relationship with common cholesterol related gene polymorphisms e.g., APOE, LPL, and CETP, to that of serum lipid levels and risk of memory loss.
MATERIALS AND METHODS
This survey study is based on different populations from the Palacode (rural) and Pappireddipatti (tribal) districts that involved 52 subjects with an average age of 57.59±6.64 y and 135 subjects with an average age of 60.91±11.46 y, respectively. Subjects were identified randomly through door-to-door census and invited to participate in the study, with written informed consent, an extensive face-to-face interview, assessments, and tests performed by a trained research group to collect baseline information on a wide range of demographic, biological, clinical, psychosocial, and behavioral variables. Everyone was interviewed for a detailed inquiry including medical history, family history, education, smoking and alcohol use, hyperlipidemia, hypertension, diabetes and coronary heart disease, etc. The questionnaire was adopted from the standard formats available from Indian Council of Medical research (ICMR) with slight modifications (Supplementary Material).
Recruiting inclusion criteria was set with respect to the age that was defined to collect samples from an aged population of above 50 years old. Particularly, the criteria identifies population with memory symptoms based on their response to questionnaires in the random population study. Exclusion criteria were set to exclude subjects with well-known major complications that are diagnosed and under treatment for cardiovascular diseases, cancer, etc.; otherwise, diabetes, hypertension, hyperlipidemia, obesity are considered in inclusion criteria, as these are the contributing factors for dementia and AD among aged.
Memory symptoms were scored and applied in the Global Clinical Dementia Rating chart to evaluate the status of the individual. Subjects scoring between 25–30 was categorized as normal; 20–25 under mild cognitive decline; 10–20 as moderate memory performance; 0–10 as a severely impaired group. An average of 25–30 questions were asked among the volunteers and the scores obtained were converted to 100% and categorized accordingly. Since the present study with rural and tribal subjects exhibited only mild to moderate symptoms of memory loss, it has been described as the presence or absence of memory symptoms among the subjects chosen for the study (Supplementary Material).
Blood samples were collected from 187 subjects of rural and tribal population of Dharmapuri District, Tamilnadu (State), India. The simplified version of Global Clinical Dementia Rating (CDR) chart was used to assess the memory performance among the volunteers aged between 50 to 80. The community survey was conducted by a group of NGO (Non-Government Organization) social workers (AAROGIYAM CHARITABLE TRUST, Regd. No. 184/2005, Dharmapuri) who were trained by the Bharathidhasan University (BDU) research group. Subjects responding to printed pamphlets, awareness, and public advertisements that were carried out by the research group of Bharathidasan University. Suitable subjects who met these initial criteria were scheduled for screening visits to determine their eligibility for participation. The study was approved by the Institutional Ethics committee for Human research (IEC: DM/2011/101/30;DM/2016/101/65) Bharathidhasan University, Tiruchirappalli, Tamilnadu. These 187 individuals constitute a random sample from two different populations who differ in their lifestyle and are used to determine the frequency change in gene polymorphism and their susceptibility for dementia condition during aging.
Biochemical analysis
A venous blood sample of 3.5 ml was collected from individuals. Serum separated from 1.5 ml of blood sample was used to determine lipids, and the remaining 2.0 ml blood was transferred to EDTA tubes to extract DNA. The levels of TC (total cholesterol), TG, HDL-C, and LDL-C in serum samples were determined by enzymatic methods using commercially available kits (ERBA TransAsia kit). To evaluate the effect of APOE genotypes on lipid values, the subjects were classified as hypercholesterolemia (total plasma cholesterol >300 mg/dl), normolipidemic (total cholesterol <200 mg/dl and serum triglycerides <182 mg/dl), and hypertriglyceridemia (plasma triglyceride levels >350 mg/dl). All determinations were performed with an ERBA Chem– 5X Clinical Chemistry auto analyzer.
Genotyping
DNA was extracted from the peripheral blood leukocytes by the phenol-chloroform method as described previously [22]. The extracted DNA was stored at– 20°C until analysis. All genotypes were examined using restriction fragment length polymorphism (RFLP) amplified by polymerase chain reaction (PCR). Information on polymorphic sites of APOE, LPL, and CETP were from the database of single nucleotide polymorphisms (SNPs) established by the National Center for Biotechnology Information. A 244-bp fragment of the APOE gene covering the codons for amino acids 112 and 158 was amplified by polymerase chain reaction using the primer pairForward (5’-ACAGAATTCGCCCCGGCCTGGTACAC-3’) and Reverse 5’ TAAGCTTGGCACGGCTGTCCAAGGA-3’) according to Yin et al. [23]. After amplification, 8 μl of the PCR product were directly digested with 12 units of the restriction enzyme HhaI (Thermoscientific) for 4 h at 37°C. Gene fragments were separated using 3% Agarose gel electrophoresis (1 : 30 h, 150 V). After digestion with HhaI, six different fragments viz., 91, 83, 72, 48, 38, and 35-bp were analyzed for polymorphic status of APOE.
A fragment of 535 bp from intron 1 of the CETP gene was amplified by PCR12 using these primers: Forward (5’– CACTAGCCC AGAGAGAGGAGTGCC-3’) and Reverse (5’-CTGAGCCCAGCCGCACACTAA-3’) [24–26] and the primer set for LPL was: Forward (5’-GATGTCTACCTGGATAATCAAAG-3’) and Reverse (5’-CTTCAGCTAGACATTGCTAGTGT-3’) respectively [27, 28]. The presence of a restriction site for the enzyme Taq I (Thermoscientific) in intron 1 was referred to as B1 and its absence as B2 for CETP polymorphism. Digestion of the 535-bp PCR products containing Taq I restriction site (B1 allele) with Taq1B yielded two fragments (361 and 174 bp), whereas one fragment (535 bp) was generated upon digestion of the PCR products exhibiting B2 allele [29]. Gene fragments were separated using 2% Agarose gel electrophoresis (1 : 30 h, 150 V). The PCR product was digested overnight with Hind III (Thermoscientific) enzyme and visualized on 2% agarose gel. The identified genotypes were named according to the results obtained based on enzymatic cleavage at respective sites. Figures 1–3 depicts the gel-separated products of APOE amplification by HhaI digestion, LPL polymorphism using Hind-III digestion, CETP gene polymorphism of Taq1 genotypes from the subjects interpreting each homozygotic and heterozygotic combination of common alleles.
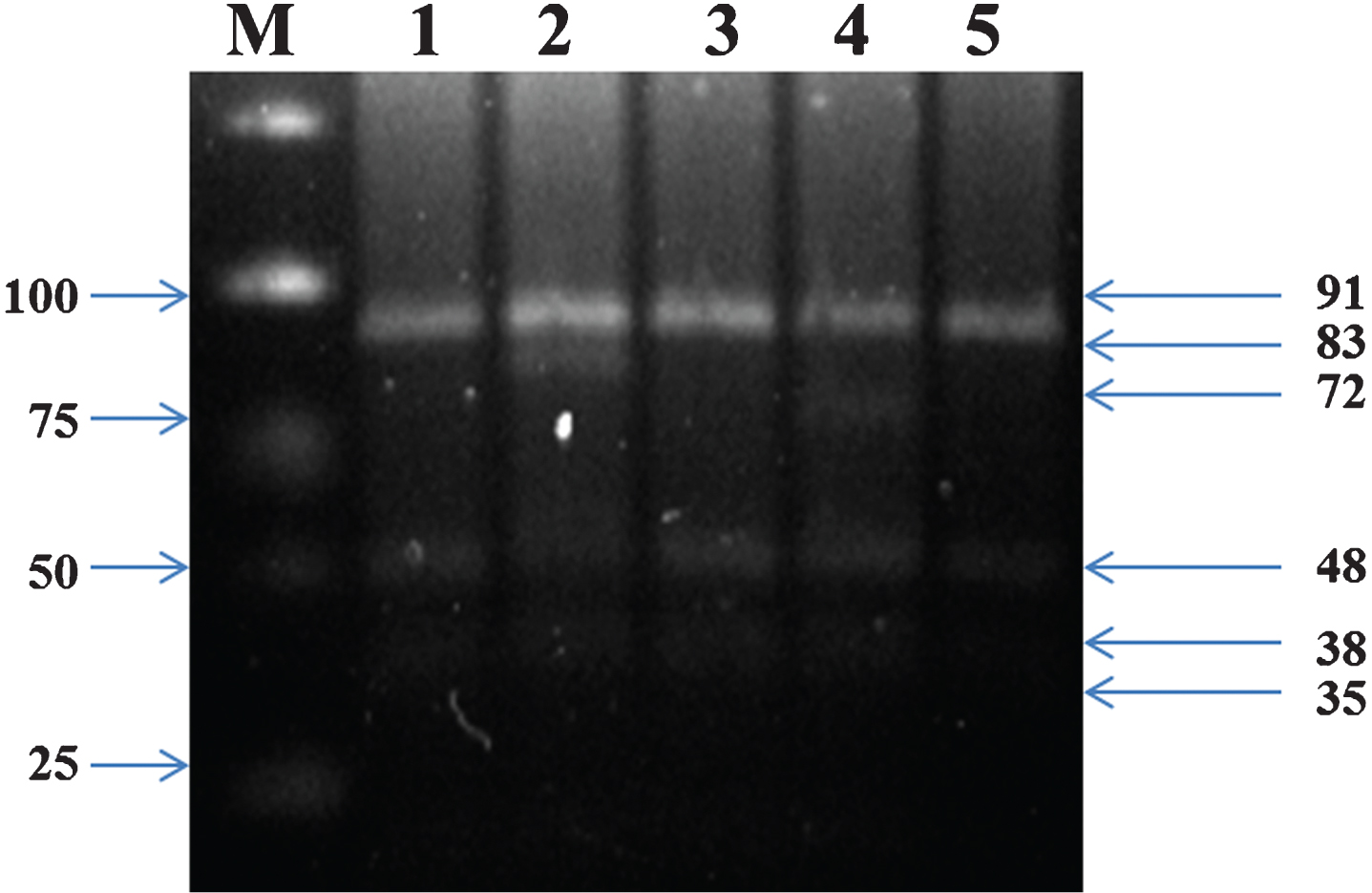
Genotyping of APOE using polymerase chain reaction and restriction fragment length polymorphism (PCR-RFLP). Lane M, marker ladder; lane 1 to 5, ɛ3/ɛ3, ɛ2/ɛ3, ɛ3/ɛ3, ɛ3/ɛ4 and ɛ3/ɛ3 genotypes, respectively.
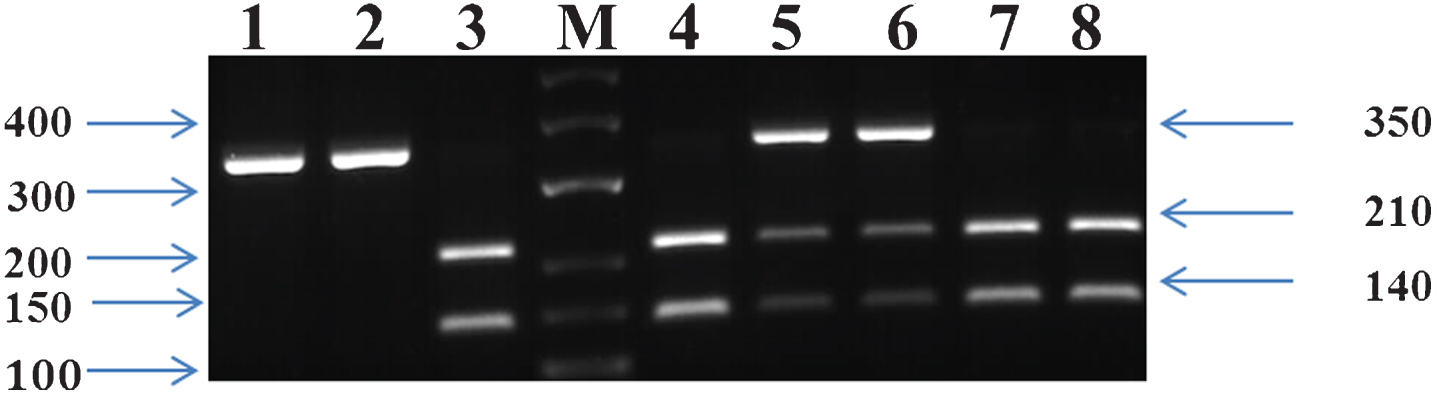
Electrophoresis pattern of HindIII polymorphism. M: Marker Lanes- 3, 4, 7, 8: H+H+ genotype (140 and 210 bp); Lanes– 5,6: H+H–genotype (140, 210, and 350 bp); and Lanes– 1, 2: H-H- genotype (350 bp).
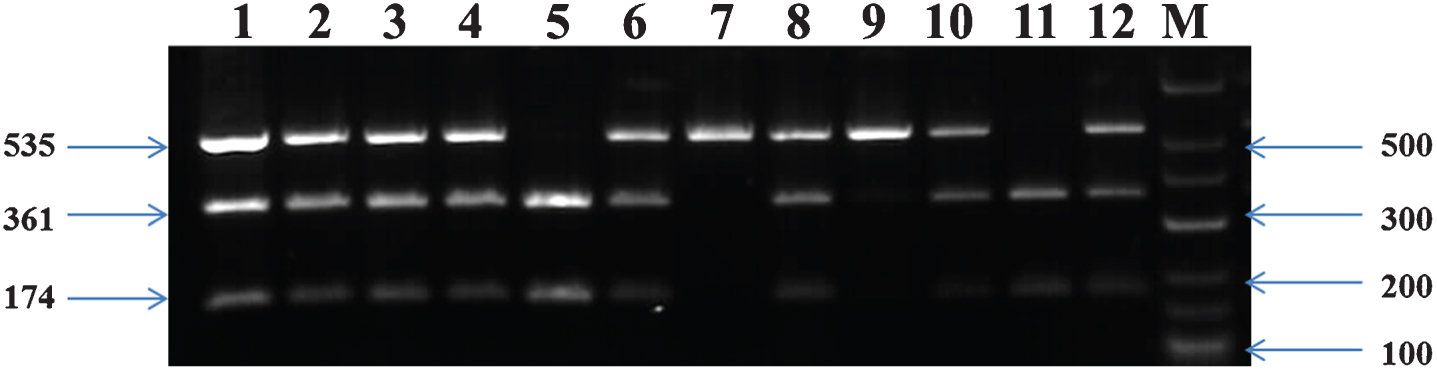
Genotyping of PCR products of CETPTaq1B. Lane M- Marker Ladder; Lanes 1,2,3,4,6,8,9,10,12, B1B2 genotype (535 bp, 361 bp and 174 bp); Lane 7- B1B1genotype (535 bp); and Lanes 5,11,B2B2 genotype (361 bp and 174 bp).
Statistical analysis
Quantitative variables were expressed as mean±standard deviation, and qualitative variables as percentages. The comparison of quantitative traits, including biochemical index and the memory scores (CDR) between rural and tribal was performed using Independent-Samples t-Test. Frequency distribution analysis of the various gene variance and the Chi-square test was studied using descriptive statistics. Univariate analysis of variance was applied to correlate the interactive effects of these gene variants on lipid profiles. Standard Goodness-of-fit was used to test the Hardy-Weinberg equilibrium. Logistic regression analysis was performed to evaluate the simultaneous influence of these genetic variations and some environmental factors on the risk of developing memory loss and Dementia symptoms. All statistical tests were two-sided. p < 0.05 was considered statistically significant. All Statistical tests were carried out using SPSS 16.0 Software.
RESULTS
General characteristics
There was no significant difference between rural and tribal groups on body mass index (BMI), fasting blood glucose, HDL, and the percentage of smokers and pan chewing (p > 0.05). However, there exists a significant variation in the major lipid profile (TC, TGL, LDL, VLDL) between rural and tribal subjects (p < 0.05). The percentage of alcohol consumption and incidence of memory symptoms were significantly higher in rural population than in tribal population, reflecting the association of lifestyle changes to that of biochemical parameters. The anthropometric and demographic data were summarized along with biochemical parameters as shown in Table 1.
Anthropometric, demographic, and biochemical characteristics among rural and tribal subjects
TC, total cholesterol; TGL, triglycerides; HDL, high-density lipoprotein; LDL, low-density lipoprotein; VLDL, very low-density lipoprotein.
Comparison of genotypes and allelic frequencies of APOE, LPL, and CETP between rural and tribal groups
Previously, genetic studies on several common polymorphisms have been carried out to interpret the relationship between lipid metabolism and AD complications [26]. However, polymorphic studies on random population to study the correlation between lipid levels and memory function is limited. The present study between rural and tribal population depicted that the overall distribution of APOE gene polymorphism was significantly different between these two populations (p = 0.044). Among them, the genotype analysis depicted a higher frequency of APOE E3/E4 genotype in tribal groups when compared to rural (p < 0.05). Also, the percentage of E2/E3/E4 allele frequency displayed significance at p = 0.056 between rural and tribal subjects(Table 2).
Genotype distribution and allele frequencies of the three polymorphism (APOE, LPL, CETP) between rural and tribal population of Dharmapuri District
The associated gene polymorphism, viz., LPL and CETP genotypes, did not exhibit a significant difference between rural and tribal groups (χ2 = 5.065, p = 0.079 and χ2 = 1.169, p = 0.428, respectively). However, the frequency of H+/H+ genotype of LPL was significantly lower in rural when compared to the tribal population (χ2 = 35.636, p = 0.000). Also, the percentage allelic distribution of both the genotype did not exhibit significant differences among the two populations (Table 2).
Single nucleotide polymorphism and serum lipid levels
One-way analysis depicted potential difference in VLDL and HDL levels between APOE, LPL, and CETP genotypes of rural population, whereas in tribal people, remarkable difference in VLDL, TC, and LDL levels can be observed between APOE and CETP genotype (Table 3A-C). APOE E3/E4 being the genetic risk factor for dementia including AD, interestingly depicted p < 0.05 significance on VLDL levels of tribal population. Similarly, LPL variants significantly disturbed VLDL among rural subjects displaying p < 0.05. Interestingly, the major allele B2 of CETP genetic variant exhibited its influence on TC and LDL levels among tribal subjects, while significant variation on HDL levels was found among B1B2 variants. Nevertheless, no significant association was reached for other serum lipids with SNP in both rural and tribal populations.
A) Effect of APOE (A), LPL (B), and CETP (C) genetic variants on serum lipid levels (mg/dl) among rural and tribal subjects
Combined analysis of APOE, CETP, and LPL polymorphism on serum lipid levels
Association studies were executed on the impact of combined effects of these frequent SNPs on the lipid levels. There existed evident combined genotype effect of APOE, CETP, and LPL polymorphism on lipid profile in the overall subjects. APOE combined with LPL/CETP polymorphism displayed significant change in TC and TGL (FAPOE *LPL = 2.817, p = 0.041; FAPOE *LPL = 2.925, p = 0.035; FAPOE *CETP = 2.631, p = 0.036) levels and could not find any statistical significance among other lipid parameters. Interestingly, APOE E3/E4 when combined with homozygous trait of CETP (H+/H+ and H–/H–) displayed significant increase in the TGL levels (Table 4A).
Combined analysis on APOE, CETP, and LPL on serum lipid levels (mg/dl)
Univariate analysis on association between CETP and LPL gene variants projected significant change in the levels of TC and LDL (FCETP *LPL = 2.470, p = 0.047; FCETP *LPL = 2.485, p = 0.046, respectively). In particular, the combined homozygous traits of the major allele of both CETP and LPL gene polymorphism, e.g., H+/H+ and B2B2 displayed a wide variation in the levels of TC and LDL (249.25±45.7 and 166±41.13) when compared to other genotypic variants (Table 4B).
Combined analysis on CETP and LPL on serum lipid levels (mg/dl)
Regression analysis on the effect of explanatory variables over memory function
Unconditional logistic regression analysis was performed to assess the influence of age, BMI, memory symptoms, and gene variants for the risk of developing dementia. The analysis revealed that the regression coefficients or beta weights, which include the influence of explanatory variables (BMI, age, place) on the occurrence of memory symptoms was associated with statistically significant independent effects of the predictors on memory loss (p = 0.048, 95% CI = 1.007±5.383 for BMI and p = 0.038, OR = 1.81, 95% CI = 1.059±7.393 for age and p = 0.000 for place) in individuals irrespective of their location, habitation, and life style. It also exemplifies that there is a significant control of CETP on the occurrence of memory symptoms among tribal population (Table 5A-C).
Logistic regression analysis: Risk factors for age associated memory loss
DISCUSSION
Genetic factors are considered as importantdeterminants of plasma lipid levels in adults [30, 31]. Reports have highlighted the evidences for the association of APOE E4 with cognitive decline and AD [12]. APOE is important in cholesterol transport and several other genes have also been involved in the transport and regulation of cholesterol metabolism in central nervous system. Among those, LPL regulates the body and energy balance [31] while CETP is involved in maintenance of cholesterol metabolism through HDL levels [32]. Complex age associated diseases such as dementia are likely to be influenced not only by genetic factors, but also by environmental components. However, there is no convincing relationship between gene and environment established. Thus, the functional roles and interplay between genes and environment in progressive dementia conditions remains controversial.
In the present study, the demographic data on rural and tribal subjects revealed no significant difference in the BMI, fasting blood glucose, percentage of smoking, and pan chewing. However, their lipid profile varied to a significant range with higher levels among tribal groups (p < 0.05) (Table 1). In spite of this, the tribal population did not exhibit significant memory related symptoms reflecting the association of lifestyle factors to that of genetic and biochemical parameters.
The incidence of memory symptoms with a CDR score more than 20 is considered as a key marker for the prevalence of dementia or AD like complications. In the present study, 40.4% of rural subjects aged above 50 displayed mild to moderate memory symptoms. The frequency of APOE E4 allele among rural group is not as high as compared to that of tribal population; however, they exhibited variation in the other gene polymorphism that are strictly associated with cholesterol metabolism (Table 2). Thus, the overall data indicates that the effect of APOE status on the cognitive function is likely to be associated with regulation of cholesterol metabolism.
Frequency distribution studies displayed significant over representation of APOE E3/E4 and H+/H+ among tribal groups (χ2 = 7.118, p = 0.008 and χ2 = 35.636, p = 0.000) when compared to rural groups. However, CETP variants did not display any significant difference in its frequency among rural and tribal populations (χ2 = 1.169, p = 0.428). Genetic studies revealed the distribution of APOE E4 and H+ allele, higher among tribal than rural subjects, but with no significance in B2 and B1 allele frequency (Table 2). These findings indicate that the frequency distribution of APOE and LPL polymorphism vary among these two population. It has been shown that LPL modulates the risk and pathogenesis of SAD (Sporadic Alzheimer’s disease) [33]. Notably, CETP facilitates cholesteryl ester and triglyceride exchange between lipoproteins [34]. Moreover, CETP is found in the same pathway as APOE, thereby considered as other important genetic risk factor for the development of AD. Even though the overall distribution of CETP polymorphism between rural and tribal populations did not show any significance, the minor alleles (B1B1) and the frequency distribution of common form of heterogeneous traits (B1B2) displayed wide difference between these two subgroups (χ2 = 22.167, p = 0.000; χ2 = 32, p = 0.000, respectively). This observed difference among rural and tribal population provide a clue on role of genetic polymorphism that could influence cholesterol metabolism, and thereby it could possibly contribute to disease progression.
Dyslipidemia is a complex trait caused by multiple environmental and genetic factors and their interactions [26]. This present study investigated variations of cholesterol-related genes correlating serum lipid levels to that of memory symptoms among aged population of two different groups from the Dharmapuri district. One-way ANOVA depicted potential difference in VLDL and HDL levels between APOE, LPL, and CETP genotypes in the rural group, whereas in the tribal population, remarkable difference in VLDL, TC, and LDL levels were observed between the variants of APOE and CETP polymorphism (Table 3A-C). In line with this, studies have reported that APOE genes carrying the E4 allele are responsible for delayed VLDL clearance rates in experimental models [35], which could be a probable reason for the observed change in VLDL levels among rural and tribal population. Hind III polymorphism of LPL has been shown to significantly associated with ischemic stroke risk and elevated levels of plasma triglycerides [36]. The less common B2B2 isoform of CETP was reported to be associated with altered HDL and LDL levels [37], as observed in the present study among rural and tribal subjects (Table 3C). In accordance with this, reports suggest that lower HDL levels, higher LDL levels, or increased TC are associated with AD cases when compared to control [38–40]. Hall et al. [41] had reported an obvious association of plasma TC or LDL levels with increased risk for LOAD and the rate of cognitive decline among elderly.
Interestingly, the current study displayed noteworthy differential effects on the interaction of various alleles (B2B2, B1B2 and B1B1) of CETP polymorphism, (H+/H+, H+/H–, H–/H–) of LPL polymorphism with the different alleles (E3/E4, E3/E3, and E2/E3) of APOE polymorphism (Table 3). Further association studies depicted the impact of combined effects of these frequent SNPs on the lipid levels (Table 4A, B). There existed an evident combined genotype effect of APOE, CETP, and LPL polymorphism on lipid profile in the overall subjects. APOE association with CETP exhibiting significant difference in TGL levels among the total population (FAPOE *CETP = 2.631, p = 0.036), and when combined with LPL variants, displayed significant changes in TC and TGL (FAPOE *LPL = 2.817, p = 0.041; FAPOE *LPL = 2.925, p = 0.035). In particular, populations exhibiting the less common allele APOE E2 when combined with the homozygous traits of LPL (H+/H+) and CETP (B2B2) influence significant increase in the lipid levels (Table 4A) when compared to other polymorphic combination. Similarly, CETP in combination with LPL displayed significant change in TC and LDL levels (FCETP *LPL = 2.470, p = 0.047; and FCETP *LPL = 2.485, p = 0.046 for TC and LDL, respectively). This correlates well with the previous observation on increased TC and LDL levels with B2B2 polymorphism (Table 3C). In this, the major alleles (B2B2 and H+/H+) influenced lipid levels to a greater extent, in particular TC and LDL, while there exists no other significant change in other lipid parameters. This is in accordance to the previous findings on rural subjects exhibiting these polymorphisms in higher percentage (Table 2) when compared to tribal subjects making them prone to lipid changes. These may be due to the reduced consanguinity among rural population and are common among the tribal subjects where gene manipulation is limited and therefore the incidence of disease complications associated to metabolic events are very fade as obtained from the questionnaire information in tribal population.
It remained controversial as to whether ethnic or region specificity, carrying APOE4 allele or not, difference of lifestyles such as fat intake and BMI [42], physical activity, and/or oxidative stress likely impaired apolipoprotein on lipid metabolism [43]. Regression analysis of SNPs with age-onset and memory symptoms among aged subjects revealed that there was a significant association between genetic variants of overall gene polymorphism to that of memory performance (Table 5), which is consistent with previous report by Presecki et al. [44].
From this study, we could track that gene polymorphism associated changes in lipid levels were further linked with memory performance in the rural population who were exposed to different kinds of food habits and are found to be more prone to poor memory performance compared to tribal population. It has been proposed that some specific nutrients and physical exercise can potentially contribute to the memory functions acting via the hippocampal plasticity [45]. This could partly support the hypothesis that lifestyle, such as consumption of naturally occurring antioxidants and enhanced physical activity in association with pollution free environment, may contribute to increased memory function as observed in tribal population regardless of APOE genotype and other polymorphisms. However, further studies using appropriate cellular and animal models are required to investigate the precise effect of APOE and other gene polymorphism that are related to lipid metabolism on the better memory function in tribal population.
Logistic regression results also showed significant interaction between CETP and biomarkers on the risk of memory loss. This is in accordance with the observation of a significant increase in TC and LDL levels with CETP variants among the subjects predicting its influence over hippocampal disturbances and memory loss. Convincing evidence by Lesser et al. [46] indicated that serum TC and LDL levels were positively correlated with densities of neuritic plaques in both neocortex and hippocampal regions. In this study, we determined an apparent association between APOE, CETP, and LPL polymorphism and memory performance. Significant positive correlations were obtained between age, memory performance, and lipid levels (TC and LDL levels) in combined subjects. These results support the presumption that lipid profile might be connected with the etiology and progress of dementia among aged. It was reported that statins could slow the rate of cognitive decline and reduced risk of dementia in the elderly [47, 48].
It is further interesting to note that tribal subjects, in spite of increased presence of homozygous traits carrying major allele particularly H+/H+ (χ2 = 35.636; p = 0.000) with significant increase in LDL, had lower incidence of hypercholesterolemia and decreased memory symptoms (Table 1). However, their HDL levels are increased significantly depicting lipid metabolism that are strictly associated with polymorphism and life style. While among rural subjects, the levels of lipids are regulated that may be due to their exposure to medical advancements and prior clinical diagnosis on various other complications that include diabetes, hypertension, etc., as observed through questionnaire information. This is therefore expected to interfere the determination on the association of polymorphism to lipid profile and memory loss symptoms and thereby considered to be the lacunae for the current study in concluding the association between polymorphism and lipid changes. Recent identification has been extensively recognized that the effect of serum TC on dementia risk that occurs in midlife but not late-life, which may be different for cardiovascular risk factor profile compared to dementia and AD [49, 50]. To conclude, lifestyle has significantly contributed to lipid profile changes that are strictly regulated by genetic polymorphism and further has extended its influence on memory performance particularly among rural subjects.
APOE and its associated genes, viz., CETP and LPL, has found to confer a significant increase in the susceptibility to disease indicating potential interaction among genes in the complement system and lipid metabolic pathway. Hence, the combined study of common genetic variants as well as environmental risk factors, may contribute to the etiological understanding of age associated complication in particular neurodegenerative diseases. Also, replication with larger sample size and more detailed individual information using much more loci variants as well as other ethnic population help to perceive the analytical facts in higher dimensions.
Footnotes
ACKNOWLEDGMENTS
The authors gratefully acknowledge Bharathidasan University for the University research fellowship (Ref. No. 05441/URF/K7/2013), CSIR for funding Mr. Karthick with SRF (09/475(0194)/2014-EMR-I) and Department of Biotechnology (DBT, No.102/IFD/SAN/PR-2459/ 2011) New Delhi, India for the financial assistance. The infrastructure of our Department is supported by the Department of Science and Technology – Fund for Infrastructure facilities in Universities and Higher Educational Institutions (DST-FIST). Our Collaborator Dr Mahesh Kandasamy has been supported by the Faculty Recharge Programme, University Grants Commission (UGC-FRP), New Delhi, India. We also thank the Non – Governmental Organization (Aarogiyam Charitable Trust, DHVANI & AWARD, Dharmapuri Dt). The authors are also grateful to Dr. M. Manickavasagam, Department of Biotechnology and Genetic Engineering, Bharathidasan University, Tiruchirappalli for his help with gel documentation studies. The authors would like to sincerely thank Ms. Sunandita Ghosh, Director, Sanya Analytics and Dr. A. Tamilselvan, Associate Professor and Head, Department of Mathematics for their support in statistical analysis and interpretation of data.