
Abstract
Herpes simplex virus type 1 (HSV-1) has long been suspected to play a role in Alzheimer’s disease (AD), the most common form of dementia. IFN-lambda (IFN-λ) is one of the key cytokine in innate antiviral defenses and, in particular, has an appreciable antiviral activity against HSV-1 infection. IFN-λ expression is regulated by the interaction between two different proteins: Mediator Complex 23 (MED23) and Interferon-Responsive Transcription Factor 7 (IRF7); single nucleotide polymorphisms (SNPs) in these genes as well as in IFNL3 were shown to be differently distributed in AD patients. In this study, allelic discrimination analysis for IFNL3 rs12979860, MED23 rs3756784, and IRF7 rs6598008, as well as IFN-λ serum concentration and anti-HSV-1 antibody (Ab) titers were performed in 79 AD patients, 57 mild cognitive impairment (MCI) individuals, and 81 healthy controls (HC) who were HSV-1-seropositive. Results showed that INF-λ serum concentration was increased in AD and MCI carrying the IFNL3 T allele compared to HC (AD versus HC: p = 0.014; MCI versus HC: p = 0.029), with the highest anti-HSV-1 Ab titers seen in AD patients carrying the IFNL3 CC genotype (p = 0.012 versus HC). Notably, anti-HSV-1 Ab titers were higher in AD and MCI individuals carrying the IRF7 AA genotype compared to HC (p = 0.018 for both). MED23 polymorphisms did not show any statistical association either with serum IFN-λ or with anti-HSV-1 Ab. Data herein suggest that the IFNL3 rs12979860 and IRF7 rs6598008 polymorphisms modulate immune responses against HSV-1 via their effect on the IFN-λ pathway. These results help to clarify the possible role of HSV-1 infection in AD pathogenesis.
Keywords
INTRODUCTION
Alzheimer’s disease (AD), the most common age-related dementia, is a progressive neurodegenerative disorder characterized by impairment of memory and other cognitive functions that cause interference with activities of daily life [1]. The main pathological features that characterize the AD brain are the extensive deposition of senile plaques resulting from the extracellular aggregation of amyloid-β peptide (Aβ), and the presence of neurofibrillary tangles as a consequence of the intracellular aggregation of hyperphosphorylated tau protein. Mild cognitive impairment (MCI) is an intermediate stage between the physiological cognitive decline due to aging and the pathological failure typical of AD that over time can convert to AD. Individuals with a diagnosis of MCI report memory deficits that, nevertheless, do not impair either global cognitive function or the ability to perform daily life activities [2].
The etiopathogenesis of AD is unclear, but genetic and environmental factors are both involved in disease onset. The only known confirmed genetic risk factor for AD is the ɛ4 allele of the Apolipoprotein E gene (APOE4) [3]; polymorphisms of several other genes have been recently being identified and could possibly become predictive biomarkers of disease development and progression [4 –7].
Environmental factors playing a role in the etiology of AD have been extensively debated. The AD pathogen hypothesis asserts that pathogens act as triggers interacting with genetic and/or immunologic factors to initiate Aβ accumulation, tau hyperphosphorylation and inflammation in AD brain [8]. Several infectious agents are suspected to be involved in the disease [9, 10]; herpes simplex virus type 1 (HSV-1), in particular, has long been suspected to play a crucial role in AD [11 –14]. HSV-1 is a double stranded DNA neurotropic virus that attains 90% prevalence of infection by the sixth decade of life. HSV-1 primary infection usually occurs during childhood or adolescence, initially involves epithelial cells of the mucosal membranes, and eventually reaches the sensory nerve terminals. From these sites, HSV-1 can spread in the nervous system, and viral genome is present in the trigeminal ganglia of the peripheral nervous system. Secondary HSV-1 reactivation is relatively infrequent, and in rare cases can cause dangerous neurological complications, including encephalitis [15].
A key role in innate antiviral defenses is played by cytokines belonging to the interferon (IFN) family. IFN-lambda (IFN-λ), in particular, consists of four structurally related IFN-λ molecules (λ1, λ2, λ3, and λ4) [16] endowed with potent antiviral activities [17 –19]. Recent results showed a relation between a specific polymorphism of IFNL3 (rs12979860) and HSV-1 reactivation in individuals with herpes labialis [20]. Notably, this polymorphism was also shown to be positively associated with IFN-λ3 serum levels and treatment outcome in hepatitis C virus infection [21, 22]. IFN-λ expression is regulated by the interaction of two proteins: Mediator Complex 23 (MED23) and Interferon-Responsive Transcription Factor 7 (IRF7) [20]. The MED23 gene is located in chromosome 6q23.2 and produces the subunit 23 of the mediator complex, a co-activator involved in the transcription of RNA polymerase II-dependent genes [23] that can directly inhibit HSV-1 replication [20]. The IRF7 gene, located in chromosome 11p15.5, is a fundamental transcriptional regulator of type I interferon (IFN)-dependent immune responses and is involved in innate immune responses against DNA and RNA viruses [24]. Recent results showed that a MED23 polymorphism is more frequent in AD patients compared to controls [25], suggesting that polymorphisms in genes involved in antiviral responses might modulate the risk of developing AD.
We evaluated the presence of possible correlations between anti-HSV-1 antibody (Ab) titers, IFN-λ concentration and polymorphisms of MED23, IRF7, and IFNL3 in individuals with a diagnosis of either AD or MCI comparing the results to those obtained in a group of age- and gender-matched healthy controls.
MATERIALS AND METHODS
Patients and controls
Eighty-two patients with a diagnosis of AD and 60 individuals with MCI were recruited in the Department of Neurorehabilitation of the Don Carlo Gnocchi Foundation. Patients were diagnosed either with probable AD according to the NINCDS-ADRDA Work Group criteria [26] or with MCI according to Petersen criteria [27]. MCI individuals were required to meet the Grundman operational criteria [28] including the presence of: 1) memory complaint, confirmed by an informant; 2) abnormal memory function, documented by previous extensive neuropsychological evaluation; 3) general cognitive status within a normal range as confirmed by both the Clinical Dementia Rating scale (CDR [29] with at least a 0.5 in the memory domain) and the Italian version of the Mini-Mental State Examination (MMSE [30] with score ≥24); 4) no impairment in functional activities of daily living; 5) no substantial cerebral vascular disease (Hachinski score ≤4; [31]); 6) no major psychiatric disease or history of depression (Hamilton Depression Rating Scale score ≤12; [32]). In addition, all MCI participants performed the Montreal Cognitive Assessment (MoCA) test [33]. To exclude reversible cases of dementia, all patients underwent complete medical and neurological exam, laboratory analyses, and MRI or CT scan. None of the patients suffered from malnutrition or vitamin deficiency syndromes. Blood count, urine analysis, blood chemistry screen, serum folate, B12 levels, and thyroid function tests were normal in all the subjects. Eighty-five healthy controls (HC) matched with patients for gender and age were also enrolled in the study.
The study conformed to the ethical principles of the Helsinki Declaration. All the individuals enrolled in the study, and their relatives when appropriate, provided written informed consent according to a protocol approved by a local ethics committee of the Don Carlo Gnocchi Foundation before admission to the study. For each participant, whole blood and serum samples were collected.
Participants with negative anti-HSV-1 IgG antibodies were excluded (3 AD patients, 3 MCI and 4 HC subjects). This selection step allowed us to define a main study sample of 217 subjects (Table 1).
Demographic and clinical characteristics of the main study population
Data are reported as mean±standard deviation or median and interquartile range. AD, Alzheimer’s disease; aMCI, amnestic mild cognitive impairment; M, male; F, female; MMSE, Mini-Mental State Evaluation; MoCA, Montreal Cognitive Assessment; APOE, Apolipoprotein E. * p < 0.0001 between AD and MCI subjects, Mann-Whitney test. # p = 0.0001 between AD and HC subjects, Chi-square test; ¶ p = 0.03 between MCI and HC subjects, Chi-square test.
Immunological analyses
Anti-HSV-1 IgG titers were measured in diluted serum (1 : 81) using commercial enzyme immunoassays (BEIA HSV-1 IgG, Technogenetics, Milano, Italy), according to the manufacturer’s instructions. Titers were evaluated using an absorbance microplate reader (SunriseTM, Tecan Group Ltd, Mannedorf, Switzerland); optical densities (OD) were determined at 450/620 nm. HSV-1 Ab titers were expressed as antibody index (AI), calculated by dividing OD measurement generated from the assay by OD cut-off calibrator. Subjects with AI >1.1 were considered seropositive, whereas subjects with AI <0.09 were considered seronegative.
Levels of IFN-λ 1/3 cytokine were measured in undiluted serum of all enrolled subjects by ELISA immunoassays (R&D Systems Inc., Minneapolis, MN, USA), according to the manufacturer’s protocol. After OD measurement at 450/620 nm by microplate reader, the data were interplated with a standard curve to obtain IFN-λ 1/3 concentrations in pg/ml; the minimum detectable dose for this kit is 8.0 pg/ml.
Genetic analyses
APOE4 genotyping
Genomic DNA was isolated from whole blood by phenol-chloroform extraction. Customer-designed Taqman probes for the 112 and 158 codons were used. Primers and probes for the 112 codon are: 112 Forward primer: 5’-GGG CGC GGA CAT GGA G-3′, 112 Reverse primer: 3′-TCC TCG GTG CTC TGG CC-5′, 112 Arg Probe: 5′-CGT GCG CGG CCG-3′-FAM, 112 Cys Probe: 5′-ACG TGT GCG GCC GCC TG-3′-VIC. Primers and probes for the 158 codon are: 158 Forward primer: 5′-TCC GCG ATG CCG ATG-3′, 158 Reverse primer: 3′-GCT CGG CGC CCT CG-5′, 158 Arg probe: 5′-CCT GCA GAA GCG CCT GGC A-3′-FAM, 158 Cys probe: 5′-CCT GCA GAA GGG CCT GGG AGT-3′-VIC.
MED23, IRF7, and IFNL3 genotyping
Single nucleotide polymorphisms (SNPs) rs3756784, rs6598008, and rs12979860 of MED23, IRF7, and IFNL3 respectively, were typed using the Taqman SNP Genotyping Assays (Applied Biosystems by Life Technologies, Foster City, CA, USA) on an ABI PRISM 7000 Sequence Detection System. For rs3756784, rs6598008, and rs12979860 respectively, the C_27496685_10, C_26775253_10, and C_7820464_10 Human Pre-Designed Assays (Applied Biosystems by Life Technologies) were used.
Statistical analyses
Parametric data are expressed as mean±standard deviation, whereas non-parametric data as median and interquartile range (IQR). Categorical data were compared using the Chi-square test. This analysis was used to exclude any deviation of SNP genotype distribution from Hardy-Weinberg equilibrium (HWE) and to compare case-control differences of SNPs distributions after gender stratification. Regarding the continuous values, parametric data were analyzed using the Analysis of Variance (ANOVA) and Student’s t-test, whereas non-parametric data were analyzed using the Kruskal-Wallis test and the Mann-Whitney test. We computed correlation coefficients using Spearman’s formula. Three binomial logistic regression models (with forward stepwise selection) were also computed, considering pathology status as response variable (model 1: AD versus HC; model 2: MCI versus HC; and model 3: AD versus MCI) and HSV-1 Ab titers as explanatory variable and inserting age, gender, and APOE4 positivity as covariates. p-values corresponding to <0.05 are described as statistically significant in the text. Corrected p-values (pc) is reported for statistical analysis corrected for covariates. The statistical analyses were accomplished using commercial software (MedCalc Statistical Software version 14.10.2, Ostend, Belgium and IBM SPSS Statistics 23.0, IBM Inc., Chicago, IL, USA).
RESULTS
Clinical characteristics
Demographic and clinical characteristics of the main study population are summarized in Table 1. Gender and age were comparable in the three groups examined; AD and MCI subjects were also matched for educational level, whereas, as expected, MMSE scores were lower in AD than MCI (p < 0.0001). We observed, as expected, a higher frequency of the APOE4 variant in AD and MCI than HC (AD: 48%; MCI: 30%; HC: 15%; AD versus HC: p = 0.0001; MCI versus HC: p = 0.03).
HSV-1 infection
Anti-HSV-1 IgG Ab titers were significantly higher in AD patients (mean±SD: 8.80±2.39 AI) than HC (mean±SD: 7.93±2.43 AI, p = 0.037), whereas MCI individuals were characterized by intermediate titers between AD and HC (mean±SD: 8.55±2.44 AI). Results of a binomial logistic regression between AD and HC (model 1), adjusted for age, sex, and APOE4 positivity/negativity, confirmed a statistically higher anti-HSV-1-specific Ab titers in AD patients (pc = 0.016; df: 1; OR: 1.22; 95% CI: 1.04–1.43). Regarding binomial logistic regression of model 2 (MCI versus HC) and model 3 (AD versus MCI) no statistical differences were observed, confirming our previous findings [34](Fig. 1).
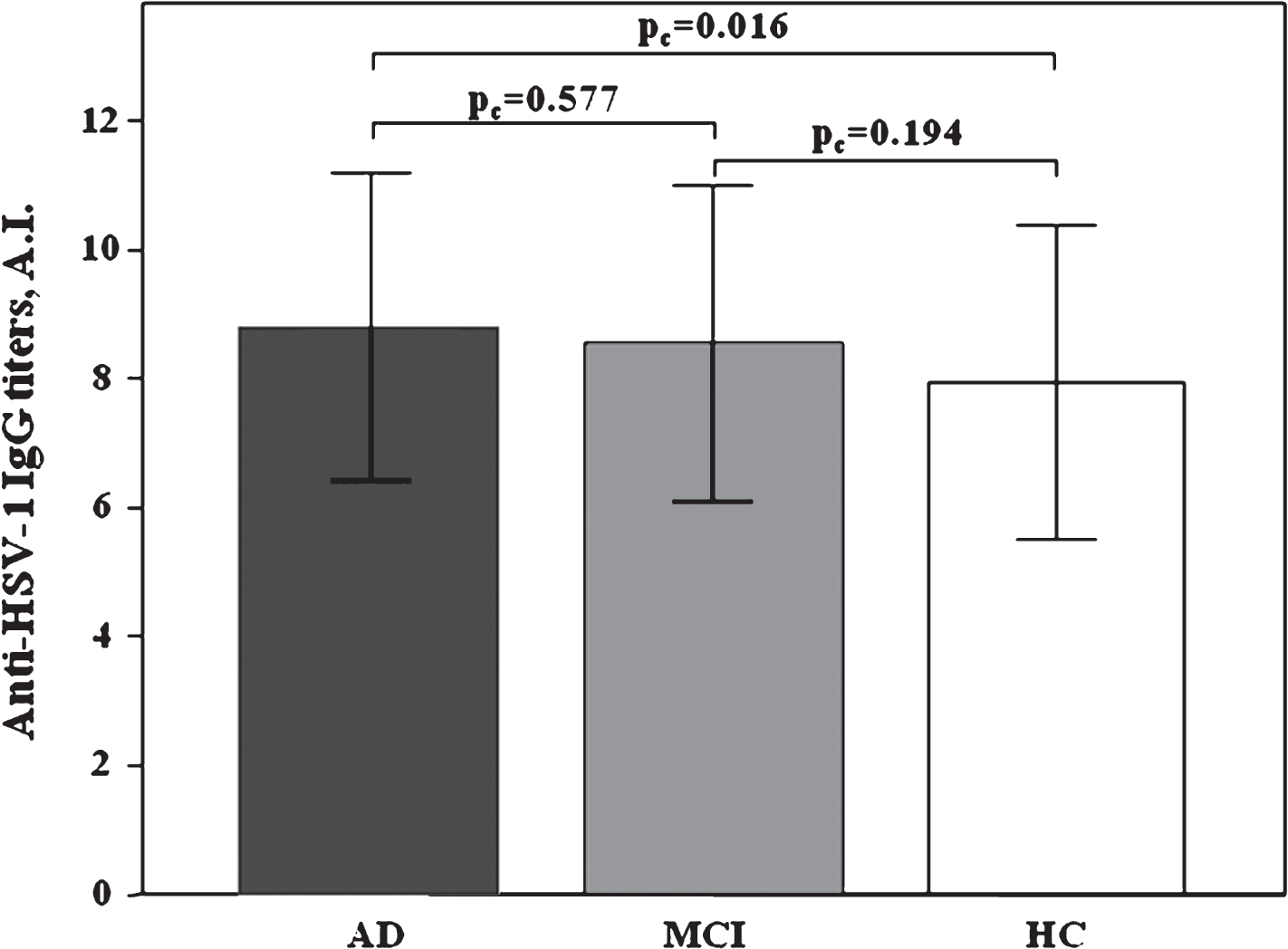
Distribution of serum anti-HSV-1 IgG antibody titers in AD, MCI, and HC subjects. AD versus HC (model 1) pc = 0.016, binomial logistic regression, MCI versus HC (model 2) pc = 0.194, and AD versus MCI (model 3) pc = 0.577. Results are expressed as mean±standard deviation. AI, antibody index.
IFN-lambda serum concentration
Serum IFN-λ concentration was higher in AD (median: 715.08; IQR: 470.05–757.74 pg/ml) and MCI (median: 712.44; IQR 498.08–758.38 pg/ml) individuals than in HC (median: 623.50; IQR:521.10–748.87 pg/ml). These differences appro-ached but did not reach statistical significance (Fig. 2). No significant correlation was found between IFN-λ concentration and anti-HSV-1 Ab titers(Supplementary Figure 1).
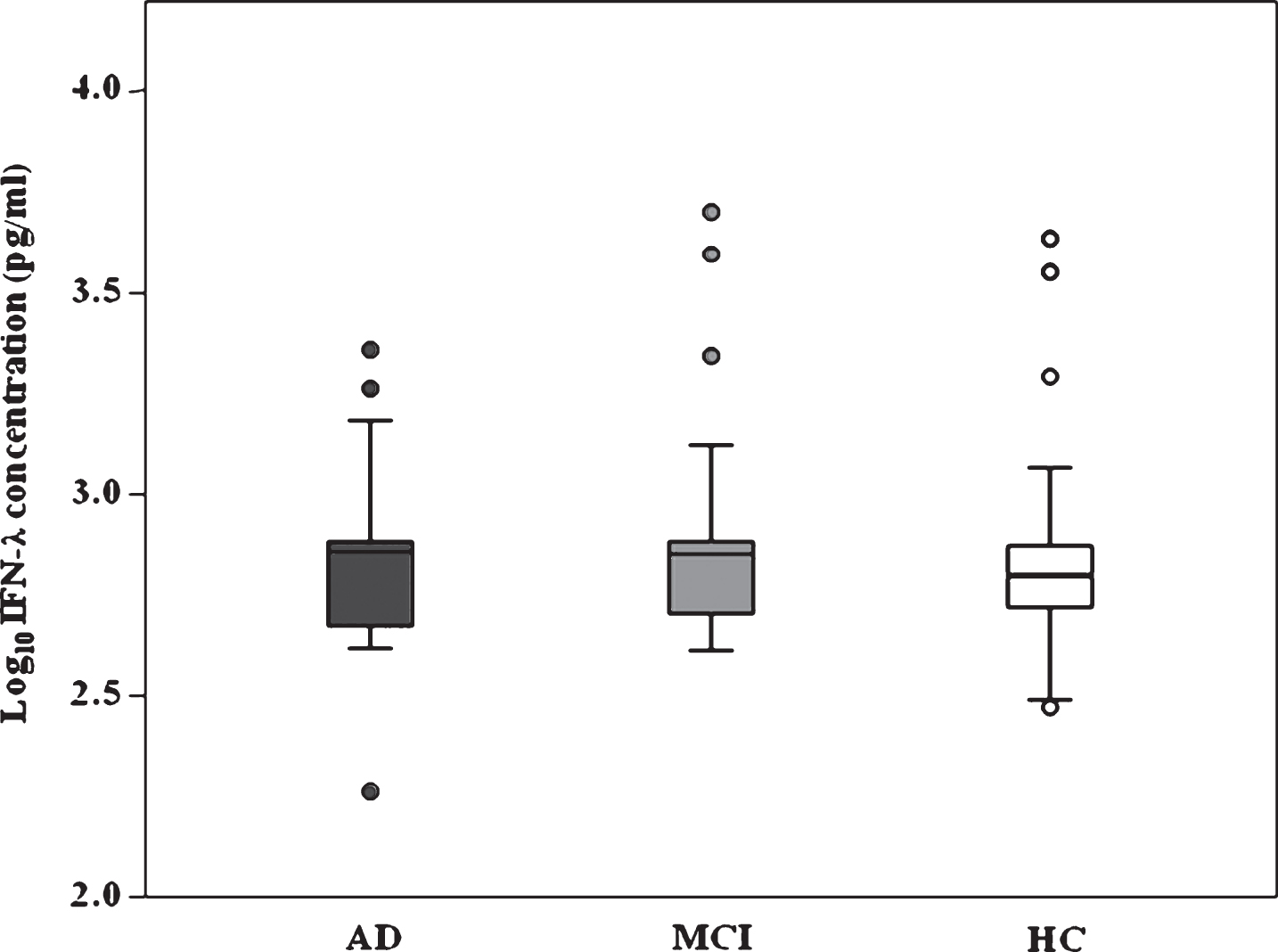
Distribution of serum IFN-λ (reported as Log-transformed) in AD, MCI, and HC subjects. The box-plot stretch from the 25th to the 75th percentile; the lines across the boxes indicate the median values; the lines stretching from the boxes indicate extreme values excluding outliers (represented by dots).
Distribution of MED23, IRF7, and IFNL3 polymorphisms
MED23 (rs3756784), IRF7 (rs6598008), and IFNL3 (rs12979860) SNPs were in Hardy-Weinberg equilibrium in AD, MCI, and HC individuals, as shown by molecular genotyping (Table 2). Genotype and allelic distribution were not statistically different in the three groups. Interestingly, stratification for APOE4 positivity showed that frequency of the MED23 G allele (GG and TG genotypes) was increased in APOE4 non-carriers AD compared to HC, although did not reach statistical significance (p = 0.08; χ 2 = 3.05) (Table 3).
Genotype distribution of MED23 rs3756784, IRF7 rs6598008, and IFNL3 rs12979860 polymorphisms in AD, MCI, and HC subjects
AD, Alzheimer’s disease patients; MCI, mild cognitive impairment subjects; HC, healthy controls.
Genotype distribution of the rs3756784 MED23 in AD and MCI APOE4+/–compared to healthy controls (HC)
AD, Alzheimer’s disease patients; MCI, mild cognitive impairment subjects; HC, healthy controls.
MED23, IRF7, and IFNL3 polymorphisms and IFN-λ serum concentration
Possible associations between MED23, IRF7, and IFNL3 SNPs and IFN-λ concentration were investigated in all the individuals enrolled in the study. Results showed that, whereas MED23 and IRF7 polymorphisms did not correlate with IFN-λ concentration (Supplementary Table 1), the IFNL3, rs12979860 T allele (CT and TT genotype) was associated with IFN-λ expression in the three groups of individuals (p = 0.025). IFN-λ concentration in IFNL3, rs12979860 T allele carriers was similar in AD (median: 748.67; IQR: 466.28–758.06 pg/ml; p = 0.014) and MCI (median: 743.21; IQR: 491.12–758.69 pg/ml; p = 0.029) individuals, but was significantly reduced in HC (median: 599.62; IQR: 520.00–750.25 pg/ml) (AD versus HC: p = 0.014; MCI versus HC: p = 0.029)(Fig. 3).

Distribution of serum IFN-λ (reported as Log-transformed) in AD, MCI, and HC subjects according to IFNL3 T allele (rs1297860) (CT and TT carriers AD versus HC: p = 0.014; MCI versus HC: p = 0.029, Mann-Whitney test). The boxes stretch from the 25th to the 75th percentile; the lines across the boxes indicate the median values; the lines stretching from the boxes indicate extreme values excluding outliers (represented by dots).
MED23, IRF7, and IFNL3 polymorphisms and anti-HSV-1 antibody titers
We also analyzed possible relationships between anti-HSV-1-specific Ab titers and MED23, IRF7, and IFNL3 SNPs. A positive correlation between anti-HSV-1-specific Ab titers and IRF7 (p = 0.023) and IFNL3 (p = 0.031) distribution was detected in AD, MCI, and HC subjects. Moreover, anti-HSV-1-specific Ab titers were significantly increased in AD (mean±SD: 8.83±2.55 AI) and MCI (mean±SD: 9.00±2.53 AI) individuals carrying the IRF7 AA genotype compared to HC (mean±SD: 6.79±2.47 AI) (p = 0.018 in both cases) (Fig. 4A).

Anti-HSV-1 Ab serum levels in AD, MCI, and HC subjects according to the IRF7 genotypes (AA carriers AD versus HC and MCI versus HC: p = 0.018 for both, Student’s t-test) (A) and IFNL3 genotypes (CC carriers AD versus HC: p = 0.012, Student’s t- test) (B). Results are expressed as mean±standard deviation. AI, antibody index.
Interestingly, considering the IFNL3 CC genotype, HC were characterized by significantly reduced anti-HSV-1-specific Ab titers (mean±SD: 7.62±2.60 AI) compared to those observed in AD (mean±SD: 9.22±2.39 AI) (p = 0.012), whereas MCI individuals showed intermediate Ab titers between AD and HC (8.37±2.09 AI) (Fig. 4B). Finally, no relation with anti-HSV-1 Ab titers could be observed when MED23 genotype distribution was analyzed (data notshown).
DISCUSSION
HSV-1 is a highly contagious herpes virus belonging to the Alphaherpesvirinae subfamily. This virus is responsible for usually asymptomatic or mildly symptomatic infections that are common and endemic in human populations. A possible connection between HSV-1 and the development of AD was first suggested more than 30 years ago [11]. Subsequently, epidemiologic, virologic, and immunologic evidences have provided further support to this hypothesis (reviewed in De Chiara et al. [9]). Excessive viral reactivation, possibly associated with weaker HSV-1-specific immunological responses, in particular might play a role in AD pathogenesis [9 , 35]. Recent data indicated that higher avidity anti-HSV-1-specific antibodies characterizes those MCI individuals in whom AD conversion is less likely to be observed [36]. IFN-λ cytokines are important human effector that obstacle viral infection, and might play a role in containing HSV-1 reactivation [37, 38].
IFN-λ expression was shown to be regulated by two different proteins: MED23 and IRF7. Thus, Griffiths and coworkers showed that the interaction between MED23 and IRF7 specifically regulates IFN-λ expression at the mRNA as well as at protein level, without any significant effect on other cytokines of interferon family [20]. Notably, recent results showed that specific SNPs of MED23, IRF7, and IFNL3 are related with different titers of Epstein Barr Virus (EBV)- and Human Herpes Virus 6 (HHV-6)-specific Ab in AD patients [25].
Possible relationships between MED23, IRF7, and IFNL3 SNPs and IFN-λ serum concentration, and between these SNPs and anti-HSV-1-specific Ab titers have not been investigated; we therefore decided to analyzed rs3756784, rs6598008, and rs12979860 polymorphisms in relation with anti-HSV-1-specific Ab titers and serum IFN-λ protein concentration in an Italian cohort of HSV-1 seropositive AD, MCI, and HC individuals. Our results confirmed previous data indicating that anti-HSV-1 Ab titers are increased in AD patients compared to the healthy controls [34, 39] and indicated that no statistical differences could be detected in IFN-λ serum concentration when results obtained in AD, MCI, and HC were compared. No correlations could be detected either between the rs3756784 and rs6598008 SNPs, which are located upstream of MED23 and IRF7 respectively, and IFN-λ serum concentration either in patients or in healthycontrols.
Notably, though, multivariate analysis results of possible correlations between MED23, IRF7, and IFNL3 SNPs and IFN-λ serum concentration as well as anti-HSV-1-specific Ab titers showed that: 1) the IFNL3 T allele (CT and TT genotype) resulted in significantly higher IFN-λ serum concentrations in AD and MCI individuals; and 2) the IRF7 AA as well as the IFNL3 CC genotypes were associated with significantly higher anti-HSV-1 serum Ab titers in AD and MCI individuals. Finally, no correlation was detected between the MED23 SNP and any of the other parameters. Interestingly, although no associations were found between MED23 and anti-HSV-1 Ab titers, significantly increased serum concentrations of these antibodies were detected in those AD and MCI individual carrying the IRF7 AA genotype, as well as in AD patients carrying the IFNL3 CC genotype, suggesting a specific relation between IFN-λ pathway and humoral immunity against HSV-1. Comprehensively, these results indicate that specific IRF7 and IFNL3 SNPs correlate with IFN-λ production and with the robustness of HSV-1-specific humoral immunity, in analogy with recent results described in EBV and HHV-6 infection [25].
In our study population, polymorphisms of MED23, IRF7, and IFNL3 were not associated with an increased risk of AD but, in accordance with previous results [25], the frequency of the MED23 G allele (GG and TG genotypes) was increased, although not significantly, in AD patients without APOE4 allele compared to HC, indicating that MED23 involvement in AD is independent from APOE4 risk factor.
Data herein indicate that the IFNL3 T allele was associated with significantly higher IFN-λ serum concentrations in AD and MCI individuals. Recent analyses performed in individuals with recurrent herpes labialis, a condition which is caused by HSV-1 reactivation, showed that the presence of this same IFNL3 allele was associated with more severe disease relapses [20]. These findings suggest that individuals with the IFNL3 T allele produce higher quantities of IFN-λ, possibly as the HSV-1 reactivation is more severe or, alternatively, because this allele might negatively influence the functionality of the IFN-λ protein and its capacity to contrast HSV-1.
An important weakness of our study is the low number of subjects enrolled; these data will need to be confirmed in a larger and independent group.
Further analyses to correlate our results with immune markers of reactivation of HSV-1 infection (i.e., IgM antibodies and increased IgG avidity) and measuring also IFN-λ mRNA expression will be necessary to shed light on the possible association between the IFN-λ pathway and HSV-1 reactivation.
Overall, results herein suggest that variations in genes involved in antiviral defensive mechanisms may play a role in modulating the pathogenesis of AD, reinforcing the hypothesis of a possible link between viral infections and dementia.
Footnotes
ACKNOWLEDGMENTS
The supplementary material is available in the electronic version of this article: https://dx-doi-org.web.bisu.edu.cn/10.3233/JAD-170520.