
Abstract
Alzheimer’s disease (AD) is a neurodegenerative disease with increasing incidence across the world and no cure at the present time. An ideal animal model would facilitate the understanding of the pathogenesis of AD and discovery of potential therapeutic targets. The Chinese tree shrew (Tupaia belangeri chinensis) has a closer genetic affinity to primates relative to rodents, and can attain ages of 8 years or older, which represents another advantage for the study of neurodegenerative diseases such as AD compared to primates. Here, we analyzed 131 AD-related genes in the Chinese tree shrew brain tissues based on protein sequence identity, positive selection, mRNA, and protein expression by comparing with those of human, rhesus monkey, and mouse. In particular, we focused on the Aβ and neurofibrillary tangles formation pathways, which are crucial to AD pathogenesis. The Chinese tree shrew had a generally higher sequence identity with human than that of mouse versus human for the AD pathway genes. There was no apparent selection on the tree shrew lineage for the AD-related genes. Moreover, expression pattern of the Aβ and neurofibrillary tangle formation pathway genes in tree shrew brain tissues resembled that of human brain tissues, with a similar aging-dependent effect. Our results provided an essential genetic basis for future AD research using the tree shrew as a viable model.
INTRODUCTION
Alzheimer’s disease (AD) is a neurodegenerative disease characterized by severe memory deficit and neuronal loss in elderly people over 65 years old [1]. The pathological features of AD are the accumulation of amyloid-β (Aβ) plaques and neurofibrillary tangles (NFTs), neuronal damage, and cognitive impairment [1, 2]. According to the World Alzheimer Report [3], 46.8 million people worldwide were living with dementia in 2015. This number will almost double every 20 years, reaching 74.7 million in 2030 and 131.5 million in 2050 [3]. To care for so many AD patients will be a huge burden for both families and society [1–4].
The ideal animal model is crucial for understanding the pathogenesis of AD, which is essential for designing a potential therapeutic approach [5–7]. Currently, several animals have been used to develop AD models. Among them, non-human primates were considered to be the ideal model for AD because of their similarities to human [8, 9]. Extensive investigation of primates including mouse lemurs and squirrel monkeys among other primates might become increasingly important models for the development of novel treatments in this domain [10–17]. However, the non-human primate model has not been used widely because of a high maintenance cost and the long aging processes in primates to form AD pathology [18, 19]. Transgenic mouse AD models have been widely used to study new therapeutic targets of AD [20, 21], but the problem with the mouse model is that therapeutic strategies are rarely translatable to AD patients [22]. Other animals, such as dog [23], roundworm [24], fruit fly [25, 26], and zebrafish [27], have also been used for AD studies. As the available AD models have many limitations, there is a pressing need to establish new experimental models of AD.
Chinese tree shrews (Tupaia belangeri chinensis) are squirrel-like, rat-sized animals inhabiting the tropical shrubs and forests of South and Southeast Asia [28] and South China [29]. The tree shrew has several characteristics and benefits as a good experimental animal, including small adult body size, easy and low-cost of maintenance, high brain-to-body mass ratio, short reproductive cycle, and a close relationship to primates [7, 30–33], for studying human diseases, such as Coxsackie virus A16 infection [34] and other diseases [7, 35]. Previous studies have shown that the tree shrew has an excellent learning ability [36–39]. There are several attempts to use the tree shrew in the study of visual cortex function [40, 41], brain development and aging [42], and neuropsychiatric disorders, such as social stress and depression [43, 44]. Moreover, Tupaia belangeri can live up to 8 years or more, which represents another advantage for the study of neurodegenerative diseases compared to primates. The important pathological feature, the deposition of Aβ, was reported in naturally aged tree shrews [45, 46]. Recently, Lin et al. [47] reported that they could induce cognitive impairment associated with neuronal apoptosis in the tree shrew by intracerebroventricular injection of Aβ1 - 40 into hippocampus. Based on these lines of evidence, there is a possibility and feasibility to use the tree shrew to replace rodents and primates in AD research.
In this study, we analyzed the AD pathway genes of the Chinese tree shrew, with an intention to learn more about the sequence identity between tree shrew and human, as well as mRNA profiling and protein expression in tree shrew brain tissues. Our results illustrate the genetic basis for using the Chinese tree shrew as a viable AD animal model.
Materials and METHODS
Experimental animals and tissue collection
The Chinese tree shrews were introduced from the experimental animal core facility of the Kunming Institute of Zoology (KIZ), Chinese Academy of Sciences (CAS). After euthanasia by diethyl ether as previously described [48], cerebral cortex and hippocampus tissues of three groups of tree shrews with different ages were quickly dissected, immediately frozen in liquid nitrogen, and stored at –80°C. These tissues were used for quantification of the AD pathway gene expression: group 1, juvenile Chinese tree shrews (1–6 months old, n = 8); group 2, adult Chinese tree shrews (3 years old, n = 8); and group 3, old Chinese tree shrews (over 5 years old, n = 8). The cerebral cortex, hippocampus, and full brain tissues from two male juvenile Chinese tree shrews (6 months old) were collected for mRNA expression profiling. For immunofluorescence assay, two male Chinese tree shrews (Juvenile, 6-month-old, n = 1; Old, 6-year-old, n = 1) were deeply anesthetized with sodium pentobarbital overdose (120 mg/kg), then were intracardially perfused with saline and 4% paraformaldehyde, following a previously described procedure for rodents [49, 50]. The experimental protocol was approved by the Institutional Animal Care and Use Committee of KIZ, CAS.
Source of gene dataset and identification of the orthologous genes
To determine the ortholog relationship between the Chinese tree shrew and human, we downloaded human protein sequence data from Ensembl (release 64; http://www.ensembl.org/index.html) and the Chinese tree shrew genome dataset (http://www.treeshrewdb.org) [51]. A total of 131 human AD-related genes were retrieved from the KEGG database [52] (map05010: Alzheimer’s disease) for a comparative analysis, following the same approach described in our previous study of the tree shrew locomotion system [53]. In brief, the longest transcript was chosen to represent each gene with alternative splicing variants. We aligned the corresponding human orthologous protein sequences of these genes onto the Chinese tree shrew genomes by tblastn [54]. The best hit regions of each gene with 5 kb flanking sequence were cut down and re-aligned using GeneWise [55]. We identified the ortholog relationship among other species through a two-step method: First, we obtained the ortholog relationship from Ensembl one2one orthologous gene list using human gene as the reference and downloaded coding region nucleotide sequences and protein sequences according to IDs from Ensembl (release 64; http://www.ensembl.org/index.html). Second, we identified the sequences of these genes not included in Ensembl one2one orthologous gene list by using the methods for computational gene prediction, as described in processing the Chinese tree shrew genome [30]. The sequence analysis of the Chinese tree shrew genome was performed at the High Performance Computing Platform, Large-scale Instrument Regional Center of Biodiversity of KIZ.
Phylogenetic analysis
To infer the phylogenetic position of the Chinese tree shrew based on the AβPP, PSEN1, PSEN2, ADAM10, and BACE1 protein sequences, we retrieved protein sequences of 7 other species (human, chimpanzee, rhesus monkey, mouse, rat, dog, and cat) from Ensambl (Supplementary Table 1). The protein sequences were aligned by Muscle 3.8 [56]. Maximum Likelihood (ML) trees were reconstructed based on protein sequence with the JTT as the amino acid substitution mode by using MEGA 6.0 [57]. Accuracies and statistical test of the phylogenetic tree were measured by bootstrap method with 1000 replications.
Detection of the positively selected genes
All coding sequences (CDS) from 6 species (human, rhesus monkey, Chinese tree shrew, mouse, rat, and dog) were aligned by MUSCLE 3.8 [56] with the guidance of aligned protein sequences. To reduce the rate of false positive prediction, we further carried out a series of filtering processes: First, we deleted all gaps and “N” from the alignments. Next, the aligned regions with more than 4 non-synonymous variants in 7 continuous residues were filtered. After deletion, if the remaining alignment was shorter than 33 residues, then the entire alignment was discarded.
To detect potential candidate genes under positive selection along the Chinese tree shrew lineage, the CODEML program from PAML4 package [58], which is a maximum-likelihood method of molecular evolution, was applied to the ortholog gene set of the 6 species. We used the same guide tree presented in our previous study [30]. First, we used a free-ratio model, which assumes an independent ratio (ω) of nonsynonymous (Ka) to synonymous (Ks) substitutions rate (ω= Ka/Ks) for each branch, to provide a rough measure of the selective pressure on each branch. Subsequently, we used branch-site model (test 2 of branch-site model) with the fixed foreground branch ω2 = 1 or non-fixed foreground branch ω2, to determine whether the gene has undergone positive selection on a foreground branch [59]. Likelihood ratio test (LRT) was performed to test whether a proportion of sites in the sequence had a statistically significant support for ω >1 on the foreground branch.
mRNA profiling and protein expression analysis
We used RNA sequencing (RNA-seq) to determine mRNA expression profiling of the cerebral cortex, hippocampus, and full brain tissues from two male juvenile Chinese tree shrews. The RNA-seq (500 bp library, 150 bp reads) was performed at the Novogene Corporation (Beijing) by using the Illumina HiSeq X Ten Platform. The raw sequencing data could be downloaded from the tree shrew Database (http://www.treeshrewdb.org) [51]. In addition, we retrieved the RNA-seq raw reads of brain tissues of human (GSE58604 [60] and GSE30352 [61]), mouse (GSE30352 [61]), rhesus monkey (GSE30352 [61] and data reported by Ye et al. [62]), and the Chinese tree shrew (GSE39150 [30]) from Gene Expression Omnibus (GEO; http://www.ncbi.nlm.nih.gov/geo/) and our previous studies [30, 62]. All the sample information was listed in Supplementary Table 2. The raw RNA-seq data were trimmed to remove sequencing adapters and reads containing one or more Ns >5%, or of low quality. We then aligned these filtered reads to the reference genome sequences using TopHat 1.4.1 [63]. We calculated the FPKM (fragments per kilobase per million mapped reads) values by using the Cufflinks software [64] to quantify gene expression level. The heatmap analysis of mRNA expression patterns in different species and tissues was performed on the basis of the FPKM values. The principal-component analysis of gene expression levels was performed using the ‘prcomp’ function in the ‘stats’ package in R program (http://www.R-project.org/). We also calculated the Pearson’s correlation coefficient matrices using the FPKM values and performed the hierarchical cluster analysis using R.
For quantification of protein expression, cortex and hippocampus tissues of the Chinese tree shrews were prepared using protein lysis buffer (Beyotime Institute of Biotechnology, P0013). Protein concentration was determined by using the BCA protein assay kit (Beyotime Institute of Biotechnology, P0012). Bovine serum albumin (BSA; Beyotime Institute of Biotechnology, P0007) was used as a protein standard. Tissue lysates (25μg total protein/sample) were separated by electrophoresis on 12% sodium dodecyl sulfate polyacrylamide gel and transferred to polyvinylidene fluoride (PVDF) membranes (Roche Diagnostics, IPVH00010) using the standard procedure, as described in our previous studies [65, 66]. After blocking with 5% nonfat dry milk in TBS containing 0.1% Tween 20 (TBST) for 2 h at room temperature, membranes were incubated with primary antibody against BACE1 (1 : 1000; Cell Signaling Technology, 5606p), GSK-3α/β (1 : 1000; Cell Signaling Technology, 5676p), Tau (1 : 1000; Cell Signaling Technology, 4019), P-Tau-181 (1 : 1000; Signalwayantibody, 11107), Neurofilament-L (1 : 1000; Cell Signaling Technology, 2837), total Aβ (1 : 1000; Cell Signaling Technology, 8243S), Aβ42 (1 : 1000; Cell Signaling Technology, 14974), GAPDH (1 : 10000; Proteintech, 60004-1-Ig), and β-actin (1 : 10000; Thermo, 1 : 10000), respectively, overnight at 4°C. The membranes were washed 3 times with Tris-buffered saline (Cell Signaling Technology, 9997) with Tween 20 (0.1%; Sigma, P1379), each time for 5 min, followed by incubation with the peroxidase-conjugated anti-mouse (lot number 474–1806) or anti-rabbit (474–1516) IgG (1 : 5000; KPL) (depends on the primary antibody) for 1 h at room temperature. The epitope was visualized using an ECL western blot detection kit (Millipore, WBKLS0500). Image J (National Institutes of Health, Bethesda, Maryland, USA) was used to evaluate the densitometry.
Immunofluorescence and confocal microscopy imaging
For immunofluorescence assay, the tree shrew cortex tissue was sectioned coronally at 10μm thickness on a cryostat (Leica, CM1850UV-1-1, Amtzell, Germany). Sections were collected on slides and were incubated for 16 h with the primary anti-mouse glial fibrillary acidic protein (GFAP) (1 : 200; Merck Millipore, MAB360), anti-mouse neurofilament-H (1 : 200; Cell Signaling Technology, #2836), and anti-rabbit total Aβ (1 : 200; Cell Signaling Technology, 8243S), respectively. After 3 washes with PBST (phosphate-buffered saline [Beyotime Institute of Biotechnology, C0221A] containing Triton X-100 [0.1%; Sigma-Aldrich, 10789704001]), each for 5 min, sections were then incubated with a FITC-conjugated anti-mouse IgG (1 : 500; Invitrogen, A21202) or anti-rabbit IgG (1 : 500; Life Technologies, A21207) secondary antibody, and nuclei were counterstained with DAPI. The slides were visualized under an Olympus FluoView™ 1000 confocal microscope (Olympus, America).
RESULTS
Evolutionary conservation analysis of the AD pathway genes in the Chinese tree shrew
We identified the orthologs of all 131 AD-related genes in the Chinese tree shrew based on the gene list of the KEGG database [52] (map05010: Alzheimer’s disease). The average protein sequence identity between human and Chinese tree shrew was 88.29%, higher than that of between human and mouse (85.11%) (Fig. 1A; Supplementary Table 3). In particular, mouse only had 52 genes with a high sequence identity (proportion of identical residues in the protein >90%), while the Chinese tree shrew had 68 such genes. These results suggested the Chinese tree shrew had a higher protein sequence identity relative to human for the AD related genes compared with that of mouse versus human.
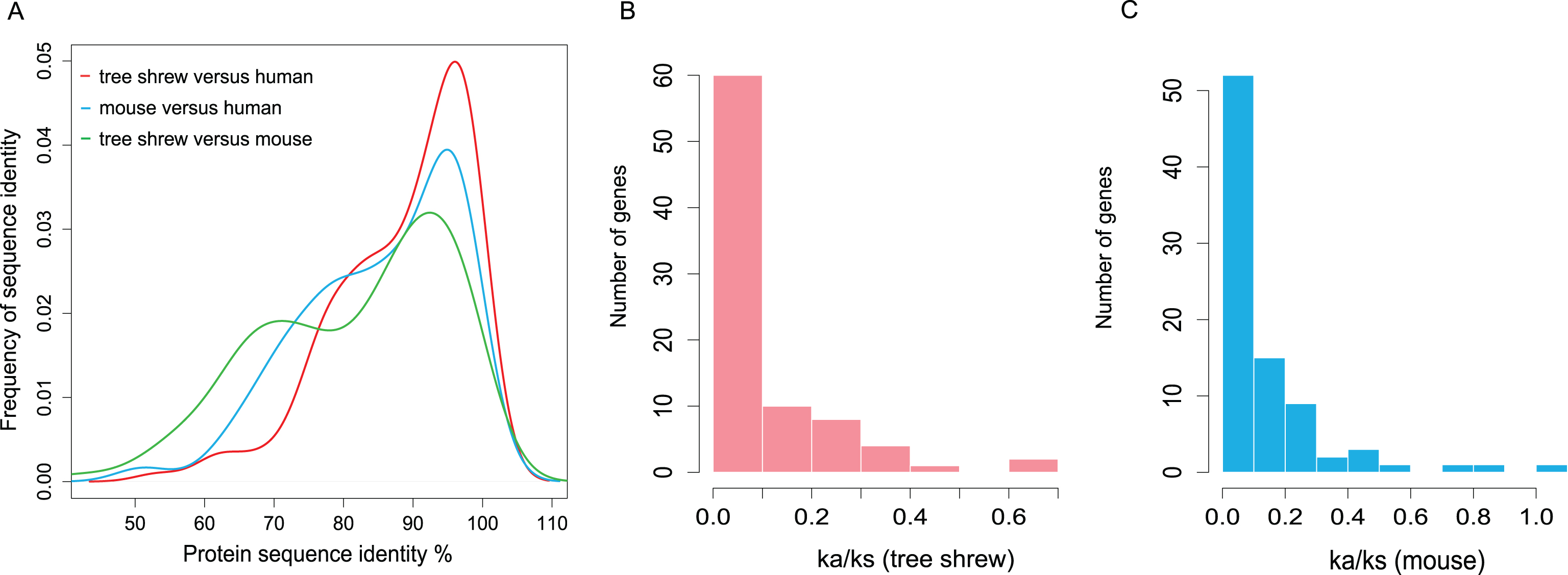
Overall protein sequence identity and Ka/Ks distribution of one-to-one orthologs between human and tree shrew and between human and mouse for the AD pathway genes. A) Protein sequence identity of one-to-one orthologs between human and tree shrew, between human and mouse and between tree shrew and mouse. B) Ka/Ks distribution of one-to-one orthologs between human and tree shrew. C) Ka/Ks distribution of one-to-one orthologs between human and mouse.
To compare the substitution rates between the Chinese tree shrew lineage and mouse lineage, we used a one-ratio model (M0) to calculate the ratio of nonsynonymous to synonymous substitution rates (Ka/Ks) on each branch of the species tree [30] using the PAML package [58]. A relatively lower level of variability in substitution rates was observed in the Chinese tree shrew lineage (average Ka/Ks was 0.1074) than in mouse lineage (average Ka/Ks was 0.1382) (Fig. 1B, C; Supplementary Table 4). We also detected the difference of selective pressure between the Chinese tree shrew and other species. Each aligned gene was evaluated for their Ka/Ks value (ω value) by using the codeml program in the PAML package [58], under the guide tree described in our previous study [30]. Then, improved test 2 of the branch-site model was used to detect signals of positive selection on each alignment. However, we did not identify any positively selected gene after adjusting for multiple testing using the false discovery rate (FDR) (Supplementary Table 4).
mRNA expression pattern of the AD-related genes
To study mRNA expression pattern of the AD pathway genes in the Chinese tree shrew, we calculated the FPKM values by using RNA-seq data of brain tissues. A total of 129 genes had an expression in brain tissues of the Chinese tree shrew (Supplementary Table 5); note that the other two AD pathway genes CACNA1C and TNF were not expressed in tree shrew brain tissues, and TNF was also not expressed in human and mouse brain tissues. We performed a two-dimensional hierarchical clustering to group the 129 genes in brain tissues of human, rhesus monkey, Chinese tree shrew, and mouse according to their Euclidean distances [67] (Fig. 2A). These genes could be classified into several expression clusters based on the similarity of their expression patterns. We observed a more similar expression pattern between the Chinese tree shrew and human than between mouse and human.
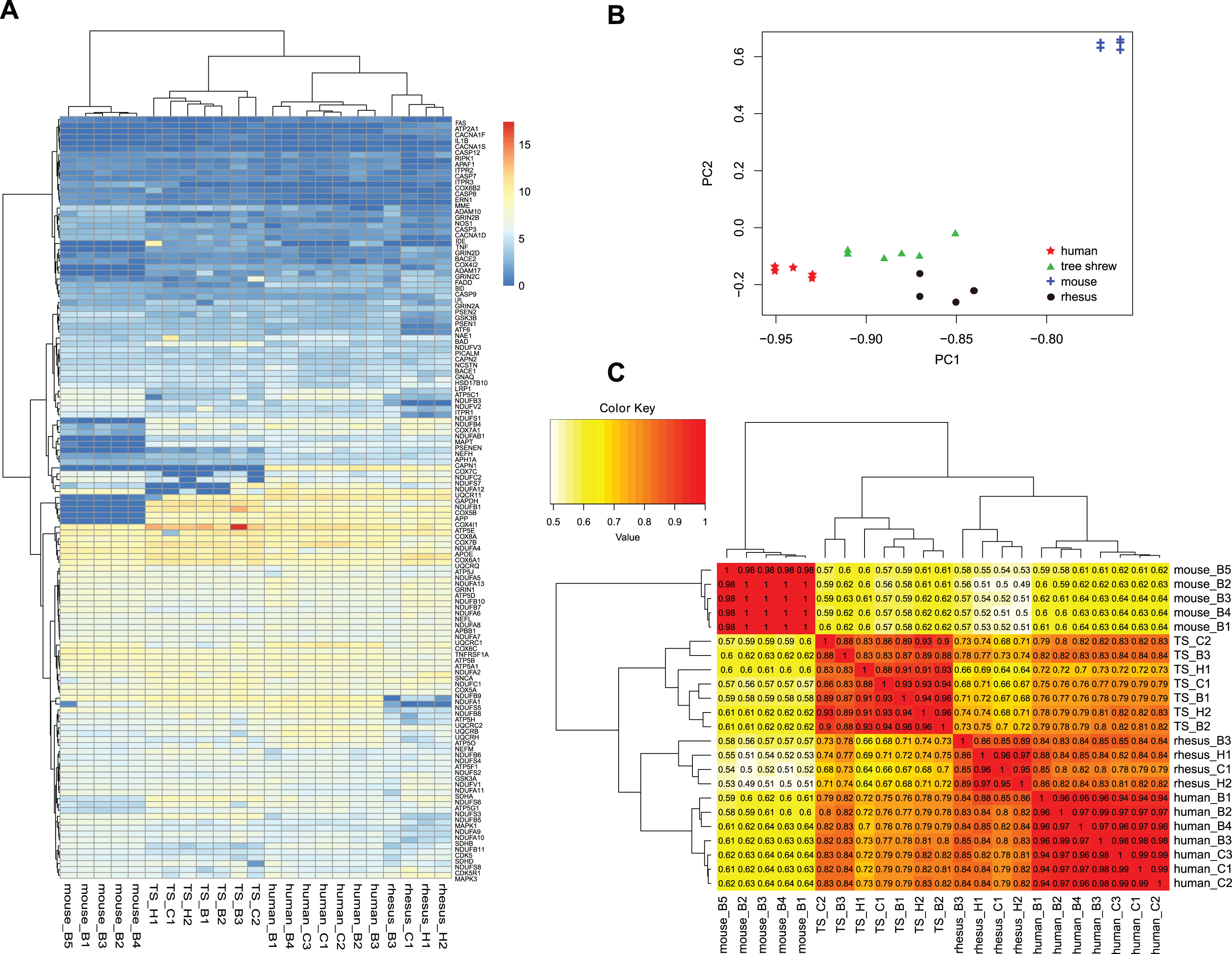
mRNA expression patterns of the AD pathway genes in brain tissues of human, rhesus monkey, tree shrew and mouse. A) A heatmap of mRNA expression levels for 129 genes from brain tissues of human (n = 7), rhesus monkey (n = 4), tree shrew (n = 7), and mouse (n = 5) based on the RNA-seq data from published sources (GSE58604 [60], GSE39150 [30], and GSE30352 [61]) and the study by Ye et al. [62]. B) Principal component analysis (PCA) and (C) hierarchical cluster analysis of mRNA expression in brain tissues of human, rhesus monkey, tree shrew, and mouse. The numbers on the cluster map refer to the Pearson’s correlation coefficients based on the FPKM values. B, brain; H, hippocampus; C, cortex; TS, tree shrew; rhesus, rhesus monkey. The reported human, mouse and rhesus monkey data were all from brain and the original sample information was listed in Supplementary Table 2.
To better understand the variation of gene expression across brain tissues of the Chinese tree shrew, rhesus monkey, human, and mouse, we performed principal component analyses and hierarchical clustering based on log2 transformed FPKM values for the 129 genes in 23 RNA-seq samples (Supplementary Table 5). Similar gene expression patterns were clearly discerned when differentiation among samples was visualized in a reduced-dimension space via the first two principal components (Fig. 2B). Together, the first principal component (PC1) and the second principal component (PC2) could explain more than 87% of the total variance of this dataset. The overall distribution of the Chinese tree shrew samples was similar to that of human samples. The resulting cluster map based on the Pearson’s correlation coefficients clearly displayed the differentiation of expression patterns in human, rhesus monkey, Chinese tree shrew, and mouse brain tissues: Chinese tree shrew had an expression pattern closer to human than to mouse (Fig. 2C). Collectively, these analyses displayed gene expression differences of the AD pathway genes among human, rhesus monkey, Chinese tree shrew, and mouse brain tissues, and suggested that the Chinese tree shrew had a more similar expression pattern to human than mouse to human. Note that the clustering pattern of species based on the overall gene expression in brain tissues showed a similar pattern to that of 129 genes (Supplementary Figure 1).
Molecular characterization of the Aβ formation pathway
Abnormal accumulation of an aggregation-prone 42-residue Aβ (Aβ42) would lead to the formation of Aβ plaques in brain, triggering the development and pathogenesis of AD [2, 68]. Aβ42 is cleaved from the protein amyloid-β protein precursor (AβPP) sequentially by β-secretase (BACE1) and then by γ-secretase [4, 69]. The accumulation of Aβ42 may be relatively rare in normal brain, and the AβPP is cleaved first by α-secretase (ADAM10), rather than by γ-secretase, precluding Aβ42 production and abnormal accumulation [1, 69]. We therefore focused on the protein sequences of AβPP, BACE1, PSEN1, PSEN2, and ADAM10 in human, Chinese tree shrew, and mouse. These five proteins in the Chinese tree shrew all showed a high sequence identity with human and the average of variance was below 3%. Consistently with previous report [70], the Aβ42 peptide sequence of the Chinese tree shrew was completely identical with human, whereas mouse was not (Fig. 3A). Note that the DNA sequences of the Aβ42 had some nucleotide differences between tree shrew and human, and had a nucleotide sequence identity up to 93%. We reconstructed a maximum-likelihood tree based on the combined protein sequences of AβPP, BACE1, PSEN1, PSEN2, and ADAM10. The tree showed that the Chinese tree shrew was clustered with primates with a high bootstrap support (Fig. 3B), which was consistent with the recognized species tree [71].

Evolutionary analysis of genes in the Aβ formation pathways. A) Peptide sequence alignment of Aβ42 in 14 vertebrate species (accession numbers in the Ensemble database were listed in Supplementary Table 6). B) Maximum likelihood trees based on the five-protein combined data (AβPP, BACE1, PSEN1, PSEN2, and ADAM10) for the Aβ formation pathway. The number on the branch referred to bootstrap value based on 1000 replications.
We performed western blot for BACE1, total Aβ, and Aβ42 in tree shrew brain tissues using human antibodies. The significantly decreased expression of BACE1 with aging was observed in tree shrew cortex tissues and there was a similar tendency in hippocampus tissues (Fig. 4A-D). Both immunofluorescence staining (Fig. 5A, B) and western blot assay (Fig. 5C) showed that total Aβ had an age-dependent increase in the Chinese tree shrew cortex tissues. Some astrocytes, as indicated by GFAP positive structures, showed Aβ-immunoreactivity (Fig. 5A). Double immunofluorescent staining revealed a co-localization of total Aβ and neurofilament-H proteins in neurons (Fig. 5B). However, protein level of Aβ42 was similar in brain tissues from the Chinese tree shrews with different ages. These lines of evidence suggested that the Chinese tree shrew had a similar AβPP pathway with human at the protein sequence identity and expression levels. It should be mentioned that the increased total Aβ spots in the old tree shrew, which seemed to be intracellular amyloid deposits and had a diffuse or compact pattern (Fig. 5A, B), did not show a typical feature as the amyloid plaques in human AD brain tissues [72, 73]. Consistent with an early study by Pawlik et al. [70], we were unable to demonstrate the amyloid plaques in brain tissues of old tree shrew by using the immunohistochemical analysis (data not shown). However, in previous studies by Yamashita et al. [45, 46], they showed the amyloid plaques in brains of naturally aged tree shrews using the same technique.

Protein expression levels of BACE1, GSK3α, GSK3β, Tau, Tau-P181, and neurofilament-L in the cortex and hippocampus tissues from the Chinese tree shrews with different ages. A, B) Quantification of protein expression in the cortex tissues from the Chinese tree shrews with different ages. C, D) Quantification of protein expression in the hippocampus tissues from the Chinese tree shrews with different ages (each age group contains 8 individuals). *p < 0.05; **p < 0.01, Student’s t test.

Immunofluorescence staining and protein expression levels of total Aβ in brain tissues from the Chinese tree shrews with different ages. Immunofluorescence staining of (A) total Aβ/GFAP and (B) total Aβ/neurofilament-H in cortex tissues from one juvenile (6-month-old) and one old (6-year-old) Chinese tree shrew. C) Quantification of protein expression levels of total Aβ and Aβ42 in the brain tissues from the Chinese tree shrews with different ages (each age group contains 3 individuals). *p < 0.05, Student’s t test.
Molecular characterization of the NFT formation pathway
NFT is another important pathological feature of AD. NFTs are mainly composed of abnormally hyperphosphorylated tau (MAPT) proteins [1, 74]. Abnormal distribution of neurofibrillary (NF) proteins (including three subunits neurofilament light [NEFL], neurofilament medium [NEFM], and neurofilament heavy [NEFH]) were also involved in the formation of the NFTs. These proteins had an increased expression level in human AD brains [75, 76]. Glycogen synthase kinase 3 (GSK3, including its two isoforms GSK3α and GSK3β) could phosphorylate tau protein and promote Aβ production [77]. We found that the key genes of the NFTs formation pathway, including MAPT, NEFL, NEFM, NEFH, GSK3A, and GSK3B, all showed a high protein sequence identity with human orthologs and the average of identity was above 91%.
We detected the protein expression of tau (MAPT), phosphorylated tau (Tau-P181), neurofilament-L (NEFL), GSK3α, and GSK3β in the Chinese tree shrew brain tissues by using human antibodies (Fig. 4A-D). GSK3β and GSK3α showed a significantly decreased expression with aging in cortex tissue (Fig. 4A, B). Tau-P181 had a significantly age-dependent increased level in hippocampus tissues (Fig. 4C, D) of the Chinese tree shrew. The other proteins, including tau and NEFL, had a similar level of expression in brain tissues from tree shrews with different ages. Taken together, the Chinese tree shrew shared a similarity with human in the NFTs formation pathway. We did not perform typical immunohistochemical assay concerning NFTs to show the presence or absence of tangles in the old tree shrews. The underlying reason for the increased level of phosphorylated tau in hippocampus tissues, but not in cortex tissues of tree shrews during aging (Fig. 4), awaits further study.
DISCUSSION
AD is a complicated neurodegenerative disease [1], and an ideal animal model would accelerate the research of this disease [5, 22]. At the present time, transgenic mouse is the most extensively used model, which had made a significant and valuable contribution for AD research [20, 78]. Due to the species difference, therapeutic strategies found in the transgenic mouse model of AD have not been successfully translated to AD patients [22].
In this study, we aimed to test whether the Chinese tree shrew, a close species to primates, has the potential as a model for future AD research. We were able to find the following lines of evidence to show that the tree shrew may be better for demonstrating the formation of the two pathological hallmarks of AD than rodents. First, we could identify all 131 AD-related genes in the Chinese tree shrew. The overall protein sequence identity of these genes between tree shrew and human was higher than that of between mouse and human. Second, the Chinese tree shrew lineage had a smaller Ka/Ks value than the mouse lineage, suggesting a higher conservation of the AD pathway gene relative to the consensus of all six mammalian species (including human) under study (Fig. 1). There were no positively selected genes in the AD pathway in tree shrew (Supplementary Table 4). Third, comparison of the overall expression pattern of the AD-related genes in brain tissues of human, monkey, Chinese tree shrew, and mouse showed that the Chinese tree shrew had a high similarity to human, resembled that of monkey. Finally, an in-depth analysis of the two AD pathological pathways that were involved in the accumulation of Aβ plaques and NFTs, showed a higher similarity in both protein sequence identity and expression pattern between tree shrew and human. In particular, the overall protein expression of BACE1, GSK3α, and GSK3β had an age-dependent decrease in the Chinese tree shrew cortex tissues; Tau-P181 showed an increased trend in hippocampus tissues. Moreover, the protein expression of total Aβ showed a significant increase in both hippocampus and cortex tissues of old Chinese tree shrews, which was consistent with those of human [79] and rhesus monkey [80]. Immunofluorescence analysis further confirmed that total Aβ had an age-dependent accumulation in naturally aged Chinese tree shrew cortex tissues, which was also reported in previous studies [45, 46]. As we did not analyze the protein expression in human brain due to inaccessibility of human samples, we checked the age-dependent pattern for these key proteins reported in previous studies [79, 81] and identified a similar pattern as observed in Chinese tree shrew: Tau protein showed an age-dependent decrease in all brain regions [81] and GSK3β was slightly downregulated during aging in human hippocampus [79]; the expression of total Aβ was significantly upregulated in human hippocampus [79]. Note that the BACE1 had a significant aging-dependent decrease in tree shrew cortex, which was different from that of human, in which a relatively stable protein level with aging was reported in human prefrontal cortex [82, 83]. The overall similarity pattern of the AD pathway genes in tree shrew with human was consistent with a previous neuropeptidomics study of the tree shrew brain tissues by Petruzziello et al. [84], in which they identified a significantly higher degree of neuropeptide homology to the equivalent sequences in humans than those in rodents. Moreover, recent studies of distribution of neuropeptides [85, 86] and whole-brain mapping of afferent projections to the bed nucleus of the stria terminalis in tree shrew brain [87] also provided an anatomical basis for using tree shrew as a good model for study neuropsychiatric diseases.
In short, the Chinese tree shrew had a higher protein sequence identity and an overall similar expression pattern of the AD-pathway genes with human compared to mouse versus human. This would suggest that the Chinese tree shrew has the potential to be a viable model for AD study, at least it appeared to be better than the extensively used mouse model of AD [20, 21]. Given the recent advance of genetic manipulation techniques, e.g., the cutting-edge CRISPR/Cas9 technique [88–92] and the establishment of the first transgenic tree shrew using the spermatogonial stem cells [93], we believe that there is a high chance of successfully creating a valid tree shrew AD model for exemplifying the key AD pathological features. Currently, we are attempting to create a transgenic tree shrew by overexpressing human AD-causing AβPP and PSEN1 mutants in tree shrew brain. With the successful creation of this AD transgenic tree shrew, we could detect whether the misfolded Aβ or tau, or post-translational modification of these proteins might have higher neurotoxicity, or whether the tree shrew brain would be more vulnerable to these molecules. Nonetheless, more neuropathological and neurodegenerative studies as a function of age are necessary to clarify these key features in the Chinese tree shrew model before concluding its superior suitability in preclinical AD research. In addition, we need to customize the related behavioral tests to characterize tree shrew’s abnormalities associated with AD. We are enthusiastic about the potential application of the AD transgenic tree shrew model in the future.
Footnotes
ACKNOWLEDGMENTS
We thank the members in Yao lab for helpful discussion. This study was supported by the Chinese Academy of Sciences (CAS; XDB13000000, QYZDJ-SSW-SMC005 and CAS zsys-02), Yunnan Province (2015HA038), and National Natural Science Foundation of China (31730037 and 31601010).