
Abstract
Frontotemporal lobar degeneration (FTLD) is a group of complex neurodegenerative disease characterized by progressive deterioration of the frontal and anterior temporal lobes of the brain resulting in different heterogeneous conditions, mainly characterized by personality changes, behavioral disturbances, such as binge eating, and deficits in language and executive functions. Null mutations in progranulin gene (GRN) are one of the most frequent genetic determinants in familial frontotemporal dementia. Recently, progranulin was recognized as an adipokine involved in diet-induced obesity and insulin resistance revealing its metabolic function. Increasing evidence suggests that neurodegenerative dementias are associated with a higher prevalence of metabolic changes than in the general population. According to these findings, the aim of this study is to investigate putative alterations in markers linked to metabolic functions (i.e., C-peptide, ghrelin, glucose-dependent insulinotropic polypeptide, glucagon-like peptide-1, glucagon, insulin, resistin, and three adipokines as visfatin, leptin, and plasminogen activator inhibitor-1 total) in sporadic and GRN-related FTLD. We found that 1) C-peptide is increased in sporadic and GRN-mutated FTLD patients; in addition, we demonstrated an anticipation of the disease in patients with the highest C-peptide concentrations; 2) visfatin is slightly reduced in the whole FTLD group; 3) resistin, an adipokine involved in inflammatory-related diseases, is specifically increased in FTLD due to GRN null mutations; 4) ghrelin concentration is specifically increased in pre-symptomatic subjects and FTLD patients with GRN mutations. These findings support the hypothesis that alterations in metabolic pattern are involved in FTLD progression highlighting novel putative targets for the development of preventive and personalized therapies.
INTRODUCTION
Frontotemporal lobar degeneration (FTLD) is a neurodegenerative condition characterized by extensive heterogeneity at clinical, pathological, and genetic levels [1]. FTLD is clinically characterizedby personality changes, behavioral disturbances, and deficits in language and executive functions [2–5]. Binge eating is frequently described in the behavioral variant frontotemporal dementia (bvFTD) subtype, being included in the diagnostic criteria’s core symptoms, but might be frequently described even in language variants of FTD, such as the primary progressive aphasias [4, 7]. FTLD and other neurodegenerative diseases are associated with a high prevalence of metabolic changes such as fluctuations in weight, insulin resistance, and cholesterol [8]. It is well known that the most FTLD cases are sporadic but up to 50% of FTLD patients has a family history of the disease [9]. Up to date, seven disease-causing-genes have been identified including progranulin gene (GRN) [10, 11], which is at present one of the most frequent genetic determinant in our cohort [12, 13]. All pathogenic GRN mutations identified so far cause the disease through a uniform mechanism, i.e., loss of functional progranulin or haploinsufficiency [14]. Progranulin protein is a multifunctional secreted adipokine with trophic, anti-inflammatory, and growth factor-like properties, characterized by dynamic changes in the expression level, glycosylation status, and cellular localization during development and in the adult organism [15–17]. Recently, it has been demonstrated that progranulin is one of the adipose tissue hormones and its circulating levels and expression in visceral adipose tissue are increased in obesity [18]. As progranulin, other proteins are involved in metabolic changes and have multifunction properties. For example, resistin and PAI-1 are inflammatory markers altered in metabolic disorders such as obesity [19–21] and may exert disparate effects on cognition [22, 23].In a recent study on 205 patients with different type of dementia, resistin concentrations were significantly higher as compared to the controls and leptin concentrations did not show differences [24]. Moreover, Woolley and colleagues described a satiety-related hormonal dysregulation (i.e., ghrelin, cortisol, and leptin) in bvFTD and suggested that the observed aberrant hormone levels might represent a compensatory response to the behavioral or neuroanatomical abnormalities of bvFTD [25]. The C-peptide is formed from proinsulin and co-secreted with insulin; of note, the C-peptide is a biologically active peptide hormone in its own right, with biochemical and physiological characteristics that differ from those of insulin [26, 27].
According to these findings, the aim of this study is to further clarify the role of differentmetabolic factors such as C-peptide, ghrelin, glucose-dependent insulinotropic polypeptide (GIP), glucagon-like peptide-1 (GLP-1), glucagon, insulin, leptin, plasminogen activator inhibitor-1 total (PAI-1), resistin, and visfatin in FTLD and specifically in GRN-related neurodegeneration. We hypothesized that a loss of circulating progranulin might results in metabolic changes affecting disease onset andprogression.
MATERIALS AND METHODS
Subjects
Patients underwent clinical and neurologic examination at the Memory Clinic of the IRCCS Centro San Giovanni di Dio-Fatebenefratelli, Brescia and at the Centre of Neurodegenerative Disorders- Neurology Unit, University of Brescia, Italy. Diagnosis of FTLD has been made in accordance with international guidelines [2–5]. We included in the present study a total of 157 subjects including 1) n = 43 sporadic FTLD patients; 2) n = 26 GRN mutated (GRN+) FTLD patients; 3) n = 24 pre-symptomatic GRN mutated (GRN+) family members; 4) n = 20 GRN negative (GRN –) cognitively intact family members; 5) n = 44 cognitively intact unrelated controls. Demographic and clinical characteristics of patients and controls are reported in Table 1. Biological samples were collected and stored at the Institutional Biobanks after obtaining informedconsensus (46/2004; 26/2014; 44/2016).
Demographic and clinical characteristics
*ANOVA; §t-test; °Chi-square test; ∫ Fisher exact test; BMI, body mass index; na, not available.
Genetic and biochemical analyses
DNA and serum were isolated according to standard procedures. GRN gene was analyzed by direct sequencing of exonic and flanking intronic regions as previously described [12]. The concentrations of C-peptide, ghrelin, GIP, GLP-1, glucagon, insulin, leptin, PAI-1, resistin, and visfatin were determined using a Bio-Plex Pro™ Human Diabetes Assays on a Bio-Plex 200 System array reader (Bio-Rad, CA, USA), following the manufacturer’s instructions. In brief, 12.5μl of each serum sample was diluted 1:4 by using sample diluents. Fifty microliters of the samples were incubated in a pre-wet filter plate for 1 h in the dark (300 rpm agitation, room temperature) with biotinylated detection antibody. Each captured analyte was detected by the addition of streptavidin-phycoerythrin and quantified using the BioPlex array reader. Standard curves were generated using the reference standard samples supplied by the manufacturer. Analyte concentrations were calculated using the Bio-Plex Manager software. All the measurements were performed in duplicate and patient and control samples were run together in the same plates.
Statistical analyses
The Shapiro-Wilk test was used to test for normality. Violation of normality assumption of the data was corrected by applying the logarithmic transformation. The chi-square test and the Fisher exact test were used for categorical variables. Student’s t test was used to evaluate differences between two study groups; one-way ANOVA with Bonferroni correction for independent post-hoc test was used for groups comparisons of normally distributed continuous variables. Spearman correlation analyses were performed to assess associations between the demographic and biometric variables (body mass index (BMI) and age) and biochemical markers. Analysis of covariance (ANCOVA) was used to compare the mean values of each marker between groups by adjusting for demographic and biometric variables (BMI and age) when necessary, i.e., when the variable was correlated with BMI or age. Two tailed p value of less than 0.05 was considered statistically significant. Disease free curves (Kaplan Meier) were used to compare the age of conversion of affected subjects by different concentrations of biochemical markers. The SPSS 22.0 software for Windows (IBM) was used for statistical analysis (SPSS, Inc., Chicago, IL, USA).
RESULTS
Serum concentrations of the metabolic markers C-peptide, ghrelin, GIP, GLP-1, glucagon, insulin, leptin, PAI-1, resistin, and visfatin were measured in FTLD patients (n = 69) and control subjects (n = 88). Comparing patients and controls, we found 1) increased serum concentrations of C-peptide (controls: 1041.63±739.36; FTLD: 2014.01±2002.26, p = 0.001) and resistin (controls: 7366.50±4944.76; FTLD: 10036.38±7248.11, p = 0.008) in FTLD patients; 2) decreased serum concentrations of visfatin (controls: 2983.78±2387.17; FTLD: 2254.76±1488.27, p = 0.005) in FTLD patients (Fig. 1). After excluding FTLD patients with diabetes (n = 17 FTLD patients: n = 11 sporadic FTLD; n = 6 GRN+ FTLD) all results were confirmed (data not shown). In order to evaluate the influence of C-peptide, resistin, and visfatin on disease onset, we generated Kaplan-Meier survival curves in patients stratified on the basis of protein concentration quartiles. We observed a significant anticipation of disease onset in subjects with C-peptide concentration higher than the third quartile (C-peptide > 1854.36 pg/ml; log rank test p = 0.002); while we estimated that 50% of subjects with C-peptide concentration lower than 1854.36 pg/ml reached the age of 67.63 years without presenting symptoms, this estimated age was 62.62 in subjects with the highest C-peptide concentrations (Fig. 2).

Serum biomarkers levels in FTLD patients and control subjects. A) Serum C-peptide levels (pg/ml) are increased in FTLD patients as compared to controls (****p≤0.001). B) Serum resistin levels (pg/ml) are increased in FTLD patients (**p≤0.01). C) Serum visfatin levels (pg/ml) are decreased in FTLD patients as compared to control subjects (***p≤0.005).
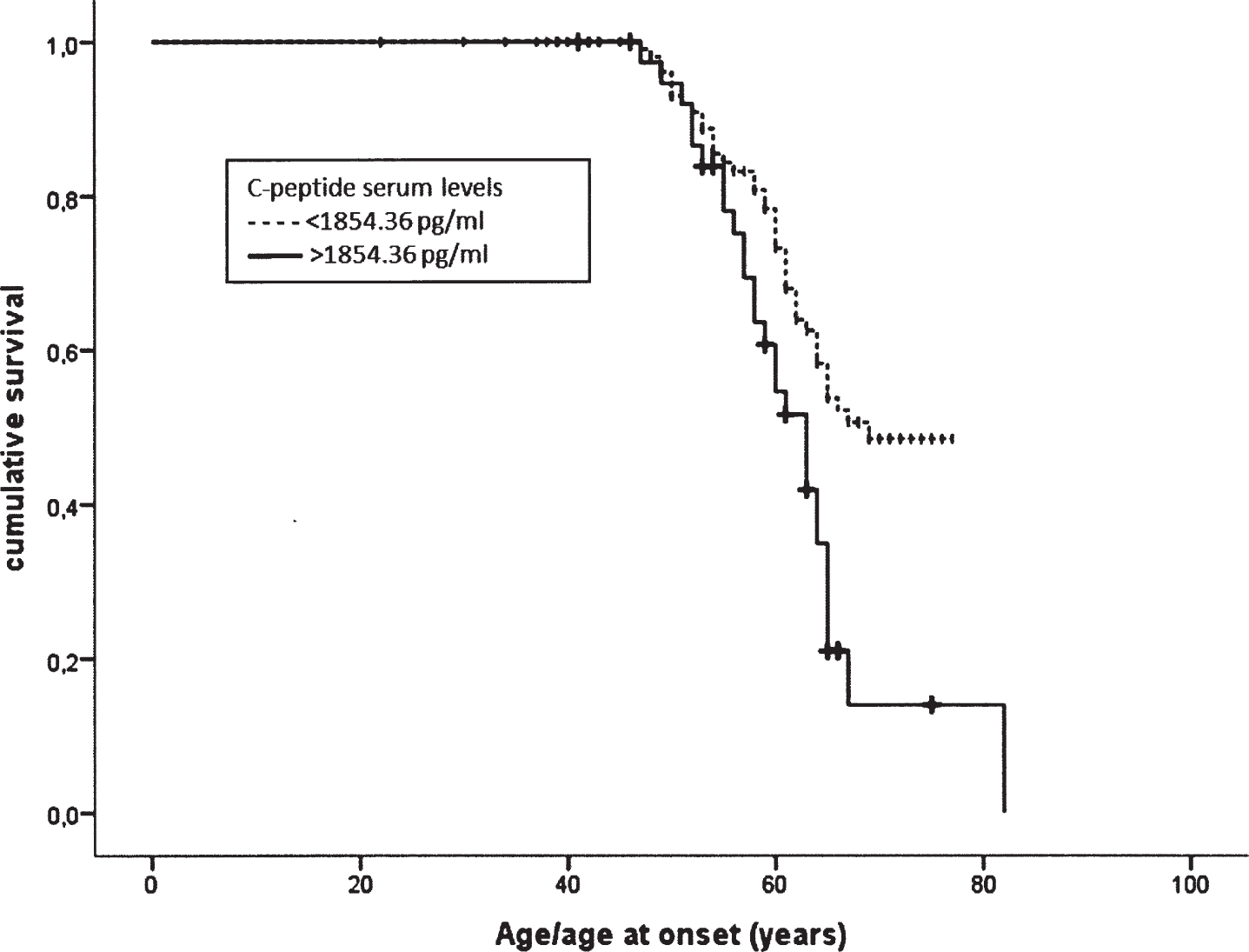
Kaplan-Meier survival curves representing age/age at onset in patients stratified on the basis of their C-peptides serum levels: a significant anticipation of disease onset was found in subjects with C-peptide levels higher than the third quartile (>1854.36 pg/ml).
In order to evaluate the concentrations of the investigated metabolic markers in GRN-related neurodegeneration, analyses were run in the following 5 sub-groups: GRN+ FTLD patients, GRN+ presymptomatic relatives, GRN- families’ members, sporadic FTLD and unrelated controls. GRN+ subjects were carrying previously described GRN null mutations (p.Leu271LeufsX10, p.Gln341X, p.Thr278SerfsX7, p.Asp22fs [28–30]. GRN+ and GRN - unaffected subjects were matched for age (p = 1.000); GRN+ FTLD patients were matched for age and BMI with sporadic FTLD and unrelated controls (p = 1.000; p = 0.054, respectively). All groups were matched for gender (Table 1).
Concentrations of serum C-peptide, ghrelin, resistin, and visfatin were different among the five study sub-groups (Table 2, Fig. 3). After excluding FTLD patients with diabetes, all results were confirmed with the exception of visfatin (p = 0.243). Moreover, post-hoc analyses revealed 1) increased C-peptide concentrations in sporadic and GRN+ FTLD patients (p = 0.001 unrelated controls vs sporadic FTLD; p = 0.010 unrelated controls vs GRN+ FTLD); 2) increased resistin concentrations in GRN+ FTLD (p = 0.015 unrelated controls vs group GRN+ FTLD); 3) increased ghrelin concentrations in GRN+ subjects (p = 0.047 GRN+ FTLD versus unrelated controls; p = 0.001 GRN+ pre-symptomatic subjects vs unrelated controls; p = 0.024 GRN+ pre-symptomatic subjects vs sporadic FTLD) (Fig. 3).
Biochemical markers serum levels
*ANCOVA; Data are reported as means±SD.

Serum biomarkers levels in the five study groups. A) Serum C-peptide levels (pg/ml) are increased in sporadic FTLD (****p≤0.001) and in FTLD GRN+ (**p≤0.01) as compared to unrelated controls. B) Serum resistin levels (pg/ml) are increased in FTLD GRN+ (*p < 0.05) with respect to unrelated controls. C) Serum ghrelin levels (pg/ml) are increased in FTLD GRN+ (*p < 0.05) and in unaffected GRN+ (****p≤0.001) as compared to unrelated controls; serum ghrelin levels in unaffected GRN+ are also increased as compared to sporadic FTLD (*p < 0.05).
DISCUSSION
Progranulin is a widely expressed, cysteine-rich, secreted glycoprotein originally discovered for its growth factor-like properties. The discovery that loss of function mutations in the gene encoding for progranulin cause dementia due to FTLD as brought renewed interest in progranulin and its functions in the central nervous system [14]. More recently, progranulin was recognized as an adipokine involved in diet-induced obesity and insulin resistance revealing its metabolic function [18, 32]. Increasing evidence suggests that neurodegenerative dementias are associated with a higher prevalence of metabolic changes (including fluctuations in weight, insulin resistance, and cholesterol concentration) than in the general population [8, 33–35]. Whether these changes result from the neurodegenerative process or whether they drive the degenerative process/have a modulating effect on neurodegeneration is still an unsolved question. A genetic association between the metabolic biomarker small apolipoprotein(a) and FTD supports the hypothesis that alteration in metabolism could be involved in the pathophysiological mechanisms of FTD [36].
Herein we investigated a panel of metabolic markers (i.e., C-peptide, ghrelin, GIP, GLP-1, glucagon, insulin, leptin, PAI-1, resistin, and visfatin) in sporadic and GRN-associated FTLD. In order to assess the role played by these metabolic markers in disease onset and progression, we included a group of pre-symptomatic subjects.
The main evidences are: 1) C-peptide is increased in FTLD; this was true both for sporadic and GRN-mutated FTLD patients. In addition, we demonstrated an anticipation of the disease in patients with the highest C-peptide concentrations. Our data are in accordance with literature data demonstrating that high C-peptide concentration is associated with cognitive impairment, rapid cognitive decline [37–39] as well as with regional cortical thinning [40] in elderly subjects; 2) visfatin, an adipokine suggested to play a role in neurogenesis [41], is slightly reduced in the whole group of FTLD patients: nevertheless, this reduction was not confirmed by post-hoc analysis on distinct sporadic and GRN-FTLD groups and after exclusion of FTLD patients with diabetes; and 3) resistin, an adipokine involved in inflammatory-related diseases [24, 43], is specifically increased in FTLD due to GRN null mutations. Resistin and progranulin are novel and putative regulators of the fat-brain axis by their ability to cross the blood-brain barrier under physiological and pathophysiological conditions [44]. A link between systemic inflammation and progranulin protein deficiency were recently established in FTLD patients carrying GRN mutations; similarly to what observed by us for resistin, serum pro-inflammatory cytokines were unchanged in pre-clinical stages but were increased in frontotemporal dementia patients [45]; iv) ghrelin concentration is specifically increased in GRN mutated subjects (both pre-symptomatic and affected). Ghrelin, a peptide hormone mainly synthetized in the stomach, has been shown to play a major role in the regulation of energy metabolism. Interestingly, a plethora of evidence indicates that ghrelin can exert important effects on neurogenesis in the hippocampus of the adult brain [46, 47] and can attenuate anxiety [48] and depression and enhance cognitive functions [49]. Thus, the serum ghrelin increase might be interpreted as a protective compensatory mechanism to counteract neurodegeneration due to progranulin deficiency as reported for other neurodegenerative diseaseincluding Alzheimer’s disease [50].
We are aware that the present study has some limitations: the results obtained in this study should be confirmed in multicenter studies; possible confounders (the presence of diabetes and BMI) were taken into consideration; other possible confounding variables (such as fatty liver and HOMA index) were not available. Thus, further studies are mandatory to confirm our findings.
Altogether, our data confirm the potential role of metabolism in FTLD pathophysiology and progression, with a specific involvement of resistin and ghrelin in GRN-related neurodegeneration. These evidences further support the existence of a cross-talk between adipose tissue and central nervous system via secreted factors [51].
These findings support the hypothesis that alterations in metabolic pattern are involved in FTLD progression highlighting novel putative targets for the development of preventive and personalized therapies in this disease.