
Abstract
Earlier we reported induction of neurotoxicity and neurodegeneration by tryptophan metabolites that link the metabolic alterations to Alzheimer’s disease (AD). Tryptophan is a product of the Shikimate pathway (SP). Human cells lack SP, which is found in human gut bacteria exclusively using SP to produce aromatic amino acids (AAA). This study is a first attempt toward gene-targeted analysis of human gut microbiota in AD fecal samples. The oligonucleotide primers newly-designed for this work target SP-AAA in environmental bacteria associated with human activity. Using polymerase chain reaction (PCR), we found unique gut bacterial sequence in most AD patients (18 of 20), albeit rarely in controls (1 of 13). Cloning and sequencing AD-associated PCR products (ADPP) enables identification of Na(+)-transporting NADH: Ubiquinone reductase (NQR) in Clostridium sp. The ADPP of unrelated AD patients possess near identical sequences. NQR substrate, ubiquinone is a SP product and human neuroprotectant. A deficit in ubiquinone has been determined in a number of neuromuscular and neurodegenerative disorders. Antibacterial therapy prompted an ADPP reduction in an ADPP-positive control person who was later diagnosed with AD-dementia. We explored the gut microbiome databases and uncovered a sequence similarity (up to 97%) between ADPP and some healthy individuals from different geographical locations. Importantly, our main finding of the significant difference in the gut microbial genotypes between the AD and control human populations is a breakthrough.
Keywords
INTRODUCTION
Alzheimer’s disease (AD) is a neurodegenerative disease with a yet unknown cause. Consequently, monitoring causative factors of AD as well as other neurodegenerative diseases is not possible. Such monitoring could both estimate AD risk and direct the treatment to blocking or eliminating causative factor(s). We previously reported that tryptamine and tryptophanol, the immediate metabolites of tryptophan (trp), an essential aromatic amino acid (AAA), induce cell death and neurodegeneration in cells and animals [1–3]. Trp is a product of metabolic shikimate pathway (SP), a seven-step metabolic route used by bacteria, fungi, algae, some protozoan parasites, and plants for the biosynthesis of AAA (phenylalanine, tyrosine, and tryptophan). SP is not found in human and animals, which require AAA to be obtained from bacteria or plants. The immediate decarboxylation of tryptophan results in the synthesis of tryptamine. Tryptamine upregulates at 4.1-fold the gene encoding amyloid β precursor-like protein 2 (Aplp2), which has been implicated in AD pathogenesis [4]. The tryptophan microbiota metabolites: indole-3-acetate, indole-3-aldehyde, indole, and tryptamine are aryl hydrocarbon receptor/xenobiotic-response elements (AhR-XRE) ligands [5]. The SP-AAA metabolism signifies an important role in AD pathogenesis. In the Netherlands, at the early stage of probable AD, the blood plasma total tryptophan levels were significantly reduced [6]. Studies conducted in Sweden [7], USA [8–10], Wales/United Kingdom [11, 12], Austria [13], and China [14] demonstrate reduced levels of tryptophan and other AAA in AD. Metabolomics identified alterations in blood and cerebrospinal fluid tryptophan metabolites and decreases in the plasma levels of tryptophan-containing tripeptides in AD [9, 10]. Higher plasma anthranilic acid levels were associated with greater risk of dementia [15]. Anthranilic acid biosynthesized from chorismic acid is the precursor to tryptophan. Although human cells lack SP and tryptophan biosynthesis pathway, the human-associated microorganisms, particularly gut microbiome has SP and AAA biosynthesis. The organization of trp genes within operons and the regulatory mechanisms used to control trp operon expression vary greatly—from whole-pathway operons to completely dispersed genes [16, 17]. Organisms are harmed and altered through damage to SP [18]. Altogether these findings warrant our interest in human cells-gut bacteria interactome that link the SP-dependent metabolites to neurodegeneration, albeit the data on the SP and related metabolic pathways are scarce for the human gut microbiota. Reports on the human gut microbiome in AD are not yet available.
We created algorithm parameters and database consisting of sequences for environmental bacteria to design the nucleotide primers targeting the SP-related enzymes in PCR analysis. Here, by means of PCR with the usage of DNA isolated from stool samples (SS) of AD patients, we revealed the nucleotide sequence with a significant similarity to conserved domain of bacterial NQR enzyme that catalyzes conversion of its substrate ubiquinone (Q), a SP product [19] into ubiquinol. It was observed that [G-14C] shikimate was incorporated into both menaquinone and ubiquinone by E. coli, thus providing the first evidence for the involvement of the SP in the Q synthesis [20]. We found that genes coding NQR subunits locate close to genes related to SP thus organizing putative gene operon in some bacteria. NQR presents mainly in pathogenic and marine bacteria [21]. Of special interest is a physiological role of the NQR substrate Q, which is neuroprotective for human cells [22, 23]. Q is a substrate of human NADH-Ubiquinone oxidoreductase (mitochondrial enzyme Complex I or CI) in the respiratory chain of mitochondria functions as a redox driven proton pump complex [24]. The early synaptic mitochondria perturbations were demonstrated in AD [25]. In our study, the mitochondrial neuropathology observed in the tryptamine-induced model of neurodegeneration [3]. Q, an important component of the electron transfer system, is constituted of a quinone structure and a side chain isoprenoid. The side chain length of Q differs between microorganisms, and this difference has been used for taxonomic study [26]. Reportedly, Q10 is a main Q form circulated in human blood, while most bacteria synthesize Q8 [27–29]. Q10 is produced by human cells from tyrosine. In children plasma, Q10 was detected at 0.15–1.32 mg/l and Q9 at 0.04–0.06 mg/l [30]. The source of Q9 is not clear, but origin from gut microbiota cannot be ruled out. We hypothesize here that bacterial Q8, a close analog of human Q10, can be an effective CI inhibitor. Such potential CI inhibition and/or disassembly by Q8 can derail mitochondrial functions in human. The importance of the length of the side chain of Q for Saccharomyces cerevisiae was examined by producing Q with isoprenoid chain lengths between 5 and 10 [31]. The original species of Q (in the case of S. cerevisiae Q6) had maximum functionality. However, the other species of Q could replace Q6. The authors concluded that a broad spectrum of different Q species is biologically functional in yeast cells, although cells seem to display a preference for their own particular Q type. Q10 deficit has been determined in neurodegenerative and neuromuscular diseases [32–34] and is associated with increased risk of disabling dementia [35]. No data available on Q8 in human bodily fluids. Q, a key molecule for cellular bioenergetics that is conserved from proteobacteria to humans, appeared in an ancestral proteobacterium more than 2 billion years ago [36]. Q biosynthesis has been studied only in a few model organisms. NQR is a key component of the respiratory chain of pathogenic bacteria [37].
MATERIALS AND METHODS
Collection of stool samples (SS)
The non-invasive environmental SS from 20 clinically diagnosed AD patients and 5 cognitively normal individuals were collected from pumper/diaper (a unit used for the collection of stool samples) at the Bidwell Care Center, Connecticut, USA (BCC). The control SS were also collected from the cognitively normal volunteers living at the private residents in California and Florida (USA). The SS from AD patients (11 females and 9 males, 58 to 89-year old) and 13 control SS from: 1) Florida (58-year-old female and a companion dog); 2) California (6 samples, 19–89 years old); 3) BCC, Connecticut (5 samples, 44–65 years old) were used in this study. SS were freshly collected at home or at the BCC, as environmental cotton swabs in 2-ml plastic tubes. Two or more SS were collected for each individual. The SS were transported at room temperature, stored frozen at –80°C upon receipt and until DNA extraction and purification. DNA was extracted directly from SS or from stool culture. The list of SS is included in Supplementary Table 1. The SS of a healthy companion dog used in this study because many diseases of the dog have a human counterpart, and notably AD [38]. The study protocol was approved by the Institutional Review Board of Expert Biomed, Inc. (FL, USA).
Statistical analysis
Statistical analyses were performed with Prism 7.02 software (GraphPad, San Diego, CA). Data are expressed as mean±standard deviation (SD). The groups were compared using unpaired two tailed t test. p-values < 0.05 were considered significant.
Methods for DNA extraction from SS, DNA extraction from bacteria, design of nucleotide primers for PCR, polymerase chain reaction, cloning and sequencing PCR products are included in the Supplementary Material.
RESULTS AND DISCUSSION
PCR test and DNA sequencing of cloned PCR products
A single set of nucleotide primers selected out of several designed sets of forward/reverse primers (Supplementary Figures 1–3) enabled us to reveal the AD-associated PCR products (ADPP) in 90% of tested AD patients (Fig. 1 and Supplementary Table 1) and 10% of control individuals (Fig. 2). The expected PCR product size of 249 base pairs (bp) is close to the observed 230 bp PCR product/s determined by comparison with a DNA ladder (a molecular weight marker). Figures 1–3 of the typical PCR test include the positive and negative samples resolved in 3% agarose gel that stained with 0.01% ethidium bromide. The DNA sequences of the cloned 230-bp PCR products from different AD patients (Supplementary Figure 2) were identical to each other in inserts with correct and opposite orientations. The sequence analysis revealed significant similarity between ADPP and conserved domain of Rnf-Nqr protein family. The Rnf-Nqr subunit is a family of integral membrane proteins including nitrogen fixation (rnf) proteins and Na+-translocating NADH: Ubiquinone oxidoreductase (Na+-NQR) subunits. Both NQR and electron complex RNF are multi-subunit membrane proteins. It was postulated that evolution of the Na+-NQR complex involved a functional divergence from its RNF homolog, following the duplication of the rnf operon [39]. We found that in some human gut bacteria, an apparently single gene operon includes both rnf and nqr genes. A closely-related complex, RsxABCDGE, reduces the 2Fe-2S-containing superoxide sensor SoxR, active as a transcription factor when oxidized. This family of putative NADH oxidoreductase complexes exists in many of the same bacterial species as the related NQR. We found that gene for an enzyme that supplies octaprenyl diphosphate, the precursor for the side chain of the isoprenoid quinones – ubiquinone and menaquinone – heptaprenyl diphosphate synthase component (HDS) locates in close proximity to the rnf-nqr genes probably in the same gene operon like in human gut Eubacterium sp. (GenBank: CBIL010000031.1). Another flanking gene, XRE (xenobiotic-response elements) family transcriptional regulator, locates upstream of the rnf-nqr genes in this bacteria. Therefore, we developed a method wherein the designed nucleotide primers produce PCR products of ∼230 bp in agreement with expected estimation of 249 bp, and the additional product of 300 bp in approximately 85–95% of AD samples and 10% of control samples (Fig. 4). The supplementary 800 bp PCR product was detected in 20% of AD samples. A semi-quantitative score in arbitrary (conventional) units (a.u.) was used to evaluate the ADPP abundance in SS. The intensity of the 230 bp ADPP band on agarose gel (Supplementary Table 1) was graded from 0 (negative sample) to 4 (maximal observed level). Statistical analysis demonstrated significantly higher ADPP levels in the SS of AD patients compared to control individuals (2.925±1.567 a.u. versus 0.136±0.234 a.u., respectively; p < 0.001, unpaired t test). In accordance with the quantitation of the DNA markers (Fig. 3 and Supplementary Figure 2A), 1 a.u. corresponds to ∼20 ng of DNA. Thus, the significant difference between the two groups supports the hypothesis of an essential role of gut microbiota in AD. The tested dog (Supplementary Table 1, SS13), a companion pet of a control individual (Supplementary Table 1, SS12), showed a weakly positive PCR response but no signs of mental and cognitive impairments. The analysis of the companion dog sample (SS13) might shed light on the transmission source for the agent leading to the PCR weak positivity in SS12. In the present study, this dog sample is used as a negative control. However, the risk for human persons to get infection from their pets, the possible carriers of AD-associated bacteria, remains to be more extensively tested. In spite of the small sample size, the statistically significant difference between the two groups of cognitively normal and AD gut microbial populations is a breakthrough finding.

A representative 3% agarose gel stained with 0.01% (v/v) ethidium bromide. PCR products obtained after amplification of DNA (50 ng per reaction) extracted from stool specimens of AD patients (SS1 to SS5, lanes 1 to 5, respectively) using the KTHR-F and KTHR-R primer pair. The control bacterial DNA (1 ng per PCR reaction) was isolated from Pseudomonas putida (PP) KT2440 and E. coli. DNA markers are the left lane. PCR conducted in 20 μl, the 5 μl loaded per lane. Markers (100 bp DNA Ladder, Invitrogen) loaded at 7 μl per lane (1 μg/μl). Note, the specific PCR product was undetectable with DNA extracted from the same SS of AD patients 1–5 and the usage of control sets of primers (Supplementary Figure 3 and Materials and Methods).
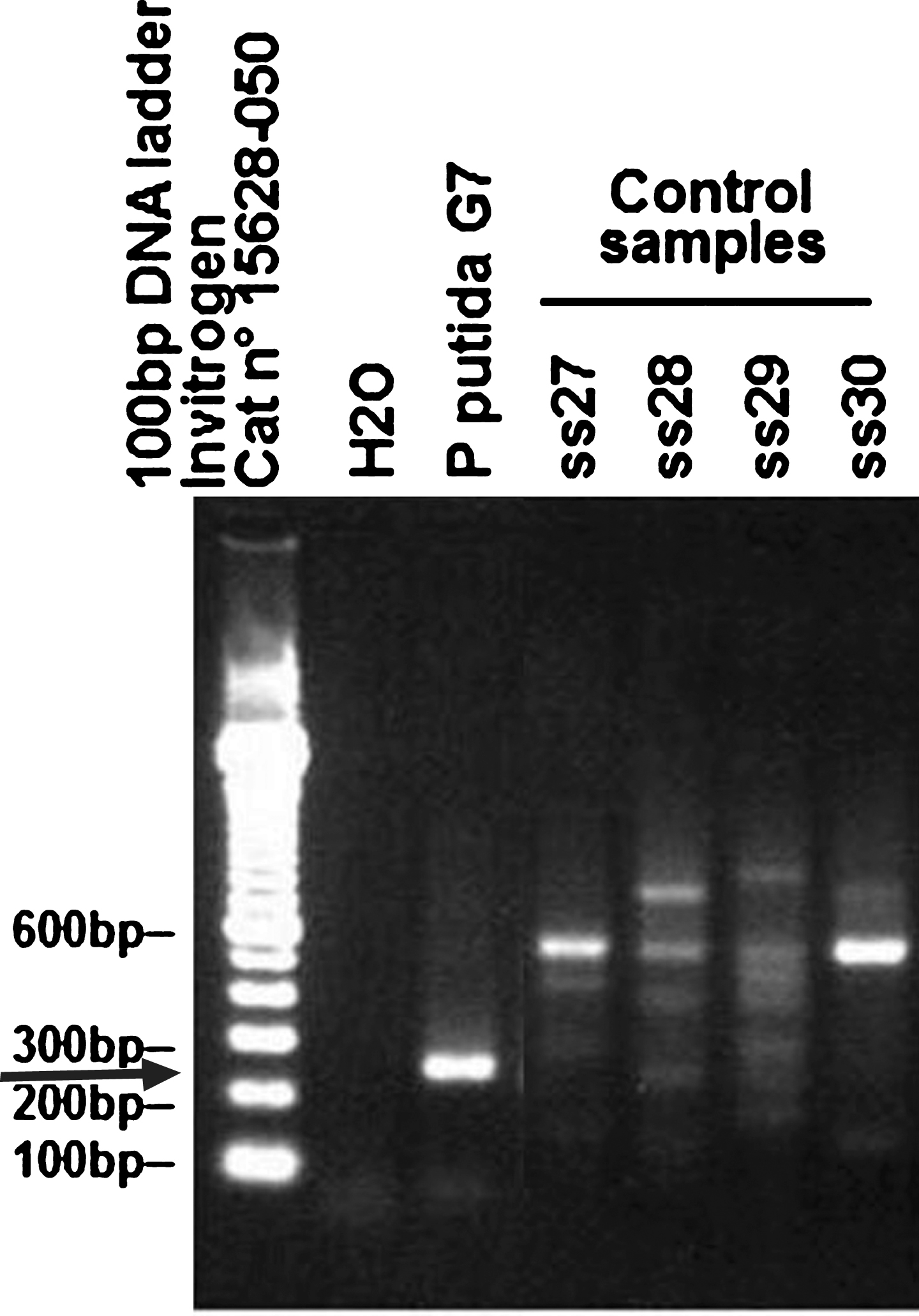
A representative 3% agarose gel electrophoresis of PCR products obtained after amplification of DNA extracted from control human stool samples (SS27 to SS30) using the KTHR-F and KTHR-R primer pair. The gel was stained with 0.01% (v/v) ethidium bromide. DNA was extracted from SS of donors with no prior diagnosis of AD. DNA markers: the left lane (Invitrogen, 100 bp ladder Cat No 15628-050). DNA extracted from Pseudomonas putida (P. putida) G7 used as a positive standard control. Arrows indicate a position/size of the expected PCR product.
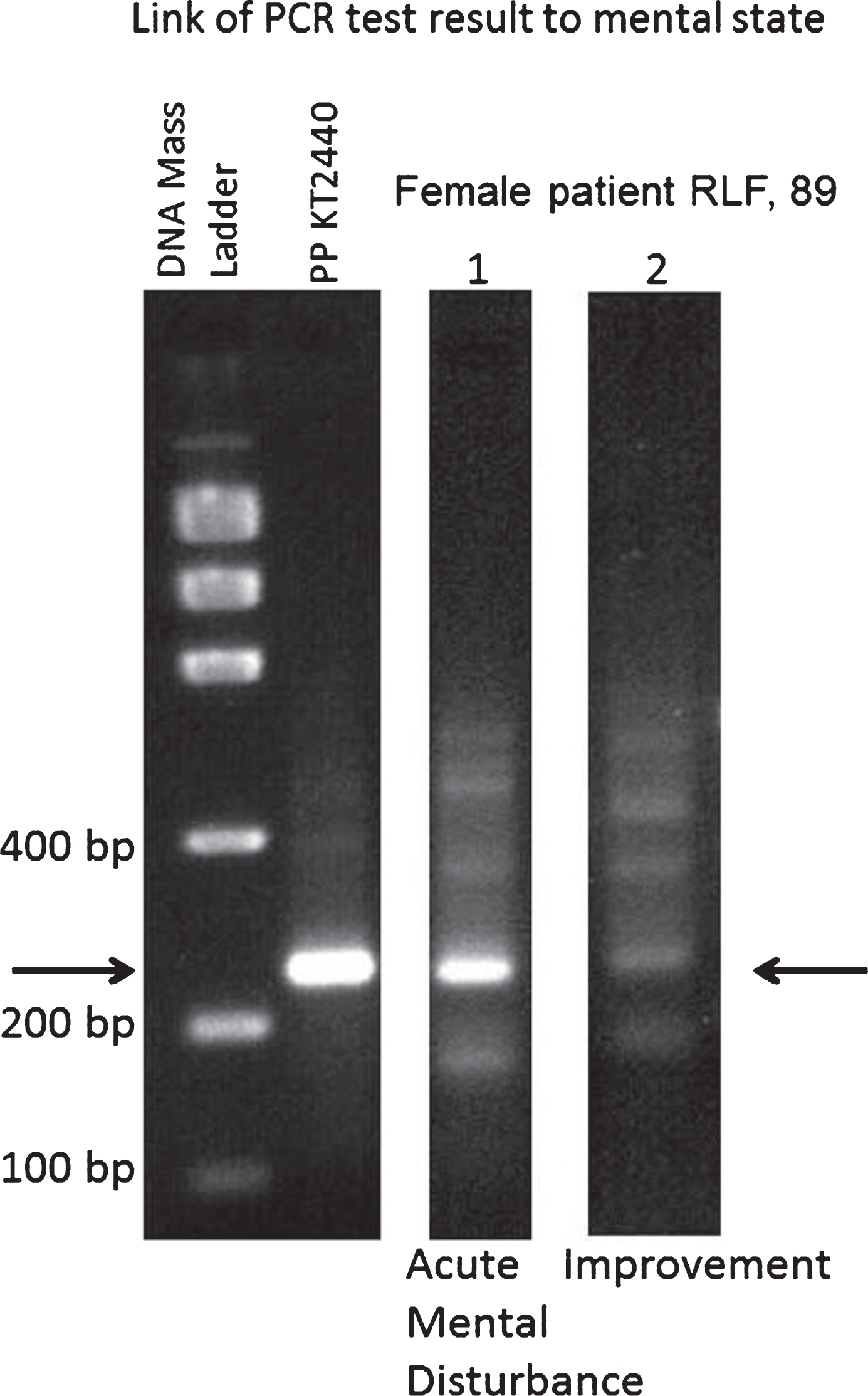
The 3% agarose gel electrophoresis of PCR products for analysis of DNA extracted from stool of case RLF with no prior diagnosis of AD: comparison of DNA specimens of the periods of active mental disturbance and mental state improvement with synthetic primers KTHR-F and KTHR-R. DNA mass ladder (Life Technologies) used for estimating the mass (quantity) of unknown DNA samples by 0.01% (v/v) ethidium bromide staining. The Low DNA Mass Ladder is composed of an equimolar mixture of six blunt-ended DNA fragments of 2000, 1200, 800, 400, 200, and 100 bp. Electrophoresis of 4 μL of the Low DNA Mass Ladder results in bands containing 200, 120, 80, 40, 20, and 10 ng of DNA, respectively.
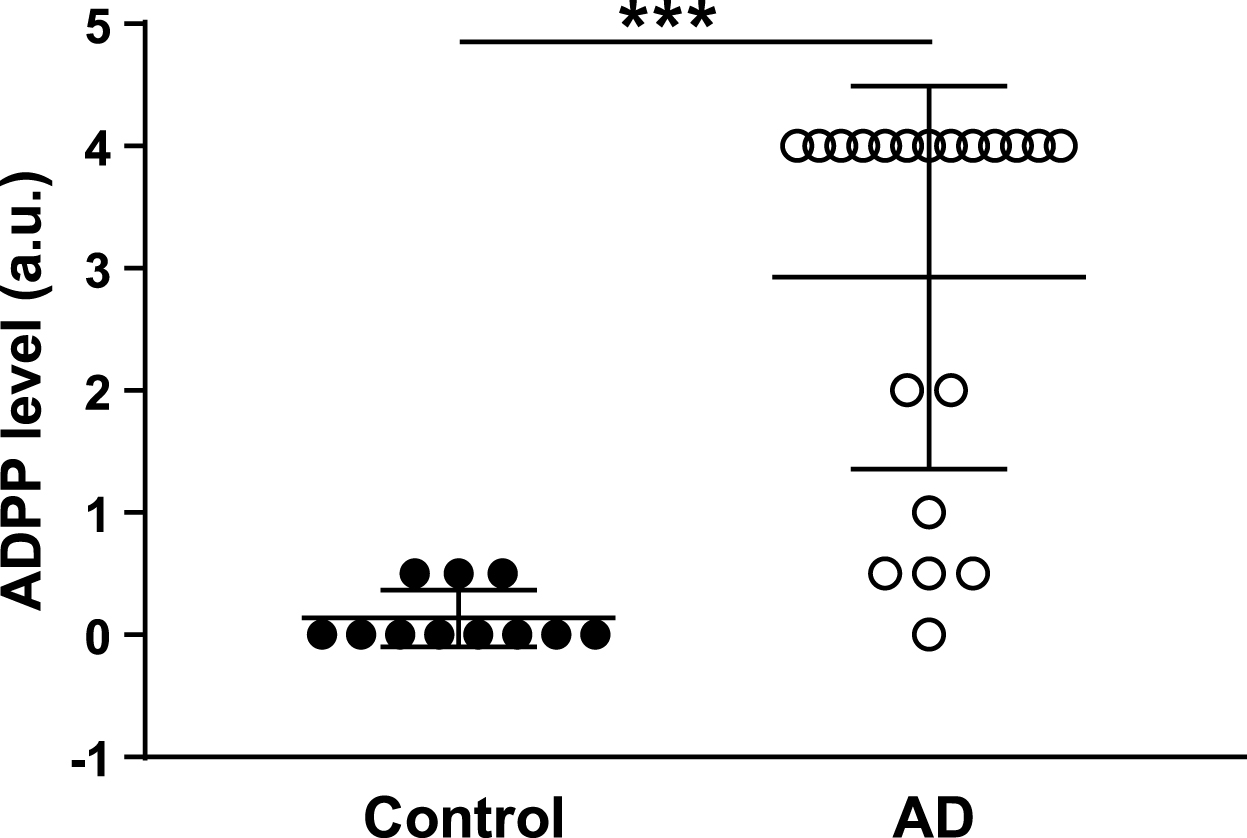
ADPP abundance in stool samples from AD patients and control individuals. PCR products generated with KTHR-F and KTHR-R nucleotide primers were analyzed by 3% agarose gel electrophoresis following by ethidium bromide (0.01%, v/v) staining. A relative intensity of the 230 bp ADPP band was scored using an arbitrary (conventional) scale (a.u.) from 0 to 4 (Supplementary Table 1). ***p < 0.001. In accordance with the quantitative DNA markers (Fig. 3 and Supplementary Figure 2A), the 1 a.u. corresponds to ∼20 ng of DNA.
In the control DNA isolated from SS of female RLF, 89, living in a private residence, California, USA, we revealed ADPP positivity. RLF had no clinically diagnosed AD but had the symptoms of acute mental disturbance with balance impairment and auditory deficits characterized by fluctuating hearing levels at the time of the first SS collection (October, 2010). The PCR product for RLF was similar to that revealed for AD patients. A second biosample of RLF (February, 2011) was collected following antibacterial treatment with Avelox (400 mg/day) prescribed due to pneumonia (Supplementary Table 1). The PCR product was barely detectable after the treatment with Avelox (Fig. 3, lane 2, Fig. 4). The mental status of RLF improved following the antibacterial therapy. In a 3-year follow-up observation, RLF had been diagnosed with dementia. In 2017, RLF has late stage AD-type dementia and does not recognize family members. The samples from RLF were provided with informed consent given by RLF’s son (his sample SS16 showed no ADPP). RLF’s sister (SS17) who lives in different location (one year younger than RLF) showed no ADPP in SS and no mental problems during the observation period 2010–2017. In the case of RLF, the appearance of ADPP correlates with the acute mental disturbance that can be suggested as an early sign of dementia that was later diagnosed. The antibacterial treatment resulted in significant reduction of ADPP that correlated with a prolonged mental improvement (Fig. 3). Note, that in May 2016, the FDA issued an Avelox (Fluoroquinolone antibiotic) warning, indicating that the drug is too dangerous to be used for many common types of infections. The young 19-year-old male, MAF (Supplementary Table 1, SS14), is a grandson of RLF (SS15, SS32, SS33). The analysis of the family members of RLF (Supplementary Table 1)—sister (SS17), son (SS16), and grandson (SS14)—might shed light on the nature of the RLF test positivity. Our data demonstrate that only one of 4 tested family members showed PCR positivity with a consequent development of AD-type dementia. The other 3 family members were negative and showed no signs of cognitive impairment. The analysis of the NCBI databases revealed human sequences similar to AD-associated PCR sequence in young individuals (Table 1). We tested SS collected in different locations and different vectors for cloning and sequencing to exclude effects of putative nosocomial infection, contamination in the research laboratory, or errors in cloning/sequencing PCR products.
Percent of nucleotide (NT) and amino acid (AA) sequence identity/similarity between AD-associated PCR product (ADPP) and human gut microbiome databases from different geographical locations (cutoff is from 86% to 97% in amino acid sequences in National Center for Biotechnology Information (NCBI) Blast alignment
Cl, Clostridium sp.; Ru, Ruminococcus sp.; Eu, Eubacterium sp.; a and b, the same individual tested twice; NT, nucleotides; AA, amino acids; UC, ulcerative colitis.
Comparison of ADPP sequence with human gut microbiome NCBI databases
USA study
The 76 amino acids deduced from a single open reading frame (ORF) of ADPP shows 97% identity (Table 1) to the sequence deduced from a single ORF of ∼30-kilobase (kb) DNA-fragment from whole genome shotgun (WGS) of uncultured unclassified bacteria Clostridium sp. from a SS of a 26-year-old obese identical co-twin (Missouri, USA). This USA human gut metagenome presents sequenced fecal microbial communities of 154 individuals including adult female monozygotic and dizygotic twin pairs concordant for leanness or obesity, and their mothers [40, 41]. Note, that metagenomic data acquired by deep sequencing of human stool collections is immensely complex, lacks apparent structure, and is typically dominated by unknown species. A large proportion (73%) of the MetaGenomic Species displays no sequence similarity to any previously sequenced organism. The human gut databases include unsupervised structuring of a highly complex series of metagenomic samples into biological entities, including a global analysis of the genetic interdependencies between bacteria, plasmids, phages, and genetic islands in the human distal gut. In some human gut bacteria containing fragment of high sequence similarity to ADPP, the rnf-ngr gene operon appears to include the gene coding HDS that supplies octaprenyl diphosphate, the precursor for the side chain of the isoprenoid quinones ubiquinone, and menaquinone. Hereupon, the hypothesis that we suggested in the Introduction that bacterial Q8 can compete with human Q10 for the binding with human Q-related enzyme(s) and affect enzymatic activity of human mitochondrial complex I (CI) is plausible. In addition, we revealed that at least 12 contigs comprising HDS present in different uncultured gut bacteria (Clostridium sp., Ruminococcus sp., Eubacterium sp., Collinsella sp.) in a US co-twin with the ADPP nucleotide-like sequence of a high identity (E value 6e-69) versus 4 contigs in uncultured Ruminococcus sp. of a co-twin with a lower similarity/identity to ADPP (E value 6e-24). The potential role of CI misassembly in CI deficiencies, AD, and Parkinson’s disease is suggested [42]. Reduced level of CI subunits in multiple zones of postmortem AD brains was reported [43]. Mammalian CI is built of 45 subunits. Fourteen “core” subunits are conserved from bacteria [44]. The Q-binding site lies at the interface of the core subunits – hydrophilic 49 kDa/PSST and membrane ND1/ND3 subunits.
Canadian study
The bacterial DNA sequence of the Clostridium sp. revealed in the USA study is near identical to DNA fragments (contigs) of ∼4 kb and ∼2 kb (99% identity, 100% cover, E value 0.0) isolated from SS of 3 healthy adults of Canadian human gut metagenome study. Bacterial fragments containing sequence near identical (97% identity in amino acid sequences) to ADPP marker were revealed in two healthy young adults from a Canadian human gut metagenome study after treatment with antibiotic Cefprozil (Table 1). This study included 24 healthy volunteers [45]. SS collected before antibiotic exposure, at the end of the treatment (7-day course), and three months later were analyzed using shotgun metagenomic sequencing. In 5 of 10 individuals, the amino acid sequence similar (85–97%) to ADPP appeared following treatment with antibiotic Cefprozil. Hence, we conclude that bacteria comprising ADPP sequence are hidden in many samples. The hidden bacteria can be uncovered following removal of antibiotic-sensitive bacteria from human gut. By other words, the Canadian study supports our suggestion that ADPP-minus bacteria can successfully compete with ADPP-plus bacteria in human gut microbiome. Note, in Canada with a 35.6-million population, over 747,000 individuals are living with AD or another dementia (http://www.alz.org/ca/dementia-alzheimers-canada.asp).
European study
Comparison of human gut metagenomes (Table 1) using the best-BLAST-hit against ADPP (E-value cutoff of 9e-42, minimum 92% coverage, 86% identity) revealed a maximum 87% identity (E-value 1e-42, 93% coverage) in amino acid sequences between the ADPP and 5 different SS of 3 females (Spanish, 36-year old, overweight with ulcerative colitis (UC); Spanish, 30-year old with UC, and Danish, 49-year old healthy overweight/obese) in the European study [46, 47]. This study includes 396 SS from 177 Danish and 141 Spanish individuals [46]; 124 samples were sequenced previously [47]. Spanish study included 13 individuals with Crohn’s disease and 69 with UC; 77 of the Spanish individuals were sampled twice with, on average, 6 months between the samplings. For two Spanish females with UC, appearance/disappearance of ADPP sequence in WGS sequence database correlated with weight gain/loss between the samplings, respectively. The Danish samples include healthy individuals ranging in body mass index 18–42. Study of prevalence of AD and other dementing disorders in Pamplona, Spain (Fig. 5) revealed 194 subjects affected by dementia and 119 had AD in the total population 1,127 [48]. The prevalence rate was 7.1%, including the very mildly demented in Denmark [49].
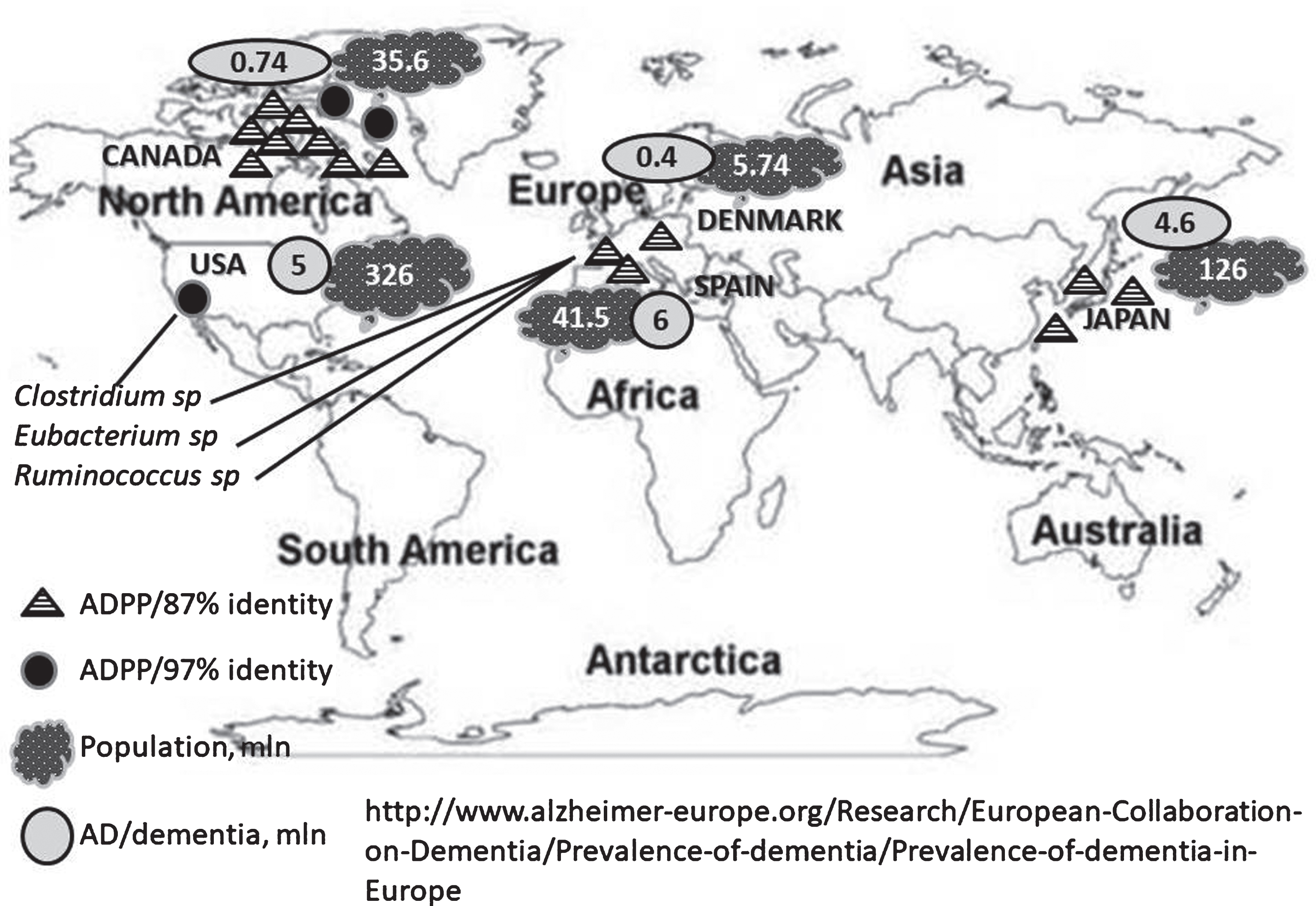
Geographical distribution of gut microbial ADPP variants in the world population. The total populations and AD prevalence in different countries are indicated in millions (mln).
Japanese study
The 87% of amino acid sequence identity (E-value 1e-42, 93% coverage) occurs between ADPP and 3 adults (one individual and a family of married couple) of the Japanese study [50]. This study included 13 healthy individuals. In Japan with a 126.6 million-population, more than 4.6 million people are living with dementia (http://www.alz.org/jp/dementia-alzheimers-japan.asp). The finding of fecal bacterial ADPP in the family (Table 1) supports the infectious nature of the environmental insult (Fig. 5).
Conclusions
This study is a first attempt toward gene-targeted analysis of human gut microbiota in AD fecal samples. The oligonucleotide primers newly-designed for this work target the environmental bacteria associated with human activity (Supplementary Figure 4). The results of the follow-up case suggest ADPP as an AD predictor. A human gut metagenome database comparative analysis and geographical distribution of the NQR-ADPP sequence is explored. We analyzed samples of our own collection and also NCBI human gut microbiota databases of the studies conducted in different geographical locations (Fig. 5). We found that human gut nucleotide/protein sequences with a high percent identity to ADPP present in USA and Canada and with a lower similarity in Europe (Denmark and Spain) and Japan. The ADPP sequence can be a part of bacterial chromosome, plasmid, or a mobile bacterial element. This prediction is in agreement with the earlier report indicating that some plasmids or plasmid families are present in the gut microbiomes of geographically isolated human hosts with a broad global distribution (America, Japan, and Europe), and are potentially unique to the human gut microbiome [51]. To create any testable model for the quantitative information on the parameters of the substrate (product) binding (release) for possible interaction between human and gut microbial NQR enzymatic activities is needed. Here the current state of knowledge of bacterial NQR conserved protein domain associated with AD is briefly summarized and discussed. Our data support an important role of microbe-host interactions in AD. The NQR bacterial sequence can be used in the population studies as a new marker for monitoring AD risk and as a new target for anti-bacterial therapy.
Footnotes
ACKNOWLEDGMENTS
This research was funded by Art Medicus PLLC, Yonkers, NY and in part (DNA cloning and sequencing) by the National Eye Institute–National Institutes of Health Grants EY02238 and P30 EY014801 to the University of Miami Miller School of Medicine Department of Ophthalmology. We thank Dr. George Perry of the University of Texas at San Antonio for reading the manuscript and comments.