
Abstract
Background:
Alzheimer’s disease (AD), the most common cause of dementia, is characterized by the intra- and extracellular aggregation and accumulation of proteins. The major molecular hallmark is the aggregation of amyloid-β (Aβ) and hyperphosphorylated tau proteins into plaques and tangles, respectively. Evidence points to the pre-fibrillary states of protein aggregates harboring the greatest neurotoxicity.
Objective:
This study was designed to identify and quantify pre-fibrillary protein species enriched by their insolubility in the detergent sarkosyl in the APPSWE/PS1ΔE9 (APP/PS1) transgenic mouse model of AD. Sarkosyl insoluble fractions were isolated from the brains of APP/PS1 and littermate wild type (Wt) mice to identify pre-fibrillary protein species associated with AD.
Methods:
Pre-fibrillary protein species were isolated from the brains of 3- and 24-month-old APP/PS1 and littermate Wt mice using sarkosyl extraction and subjected to quantitative proteomics analysis by the use of isobaric tags for relative and absolute quantitation (iTRAQ).
Results:
The sarkosyl-insoluble pre-fibrillary proteome showed differential age- and genotype-induced effects. In addition to Aβ and tau, old APP/PS1 mice showed significant enrichment in proteins in the sarkosyl fraction involved in oxidative phosphorylation and mitochondrial function.
Conclusion:
The results of this study implicate dysfunctional mitochondria as playing a key role of Aβ- and potentially tau-induced pathological events in the APP/PS1 transgenic mouse model of AD.
INTRODUCTION
Alzheimer’s disease (AD) is the most common cause of dementia, affecting more than 45 million people worldwide [1]. Protein aggregation and accumulation of aggregated proteins taking place both intra- and extracellularly are major molecular hallmarks of AD. The disease belongs to the class of neurodegenerative proteinopathies [2] and is clinically characterized by progressive, irreversible, cognitive and non-cognitive impairment. In particular, the pathological process of soluble amyloid-β (Aβ) and tau proteins converting into insoluble plaques and tangles, respectively, is known to compromise the function of synapses, likely underlying the clinical symptoms. Protein aggregation begins decades before the onset of clinical symptoms and is considered to plateau 5– 10 years before the diagnosis of dementia [3]. Although postmortem and positron emission tomography studies show that plaque deposition precedes tau pathology in the brain of AD patients [4–6], it is becoming increasingly clear that Aβ plaques and tau tangles are likely to be less toxic to neurons than the pre-fibrillar, intermediate aggregates of the proteins that compose them. For example, preclinical studies indicate that Aβ oligomers rather than fibrils [7] and hyperphosphorylated tau proteins rather than neurofibrillary tangles [8] impair synaptic function in mouse models of Aβ pathology and tauopathy. In addition, oligomeric species of Aβ and tau may interact to induce toxicity, as evidenced by the fact that injections of Aβ oligomers in tauopathy models accelerate tau-mediated neurodegeneration [9], while reducing levels of endogenous tau in amyloidosis models prevents cognitive impairment [10]. Although these studies indicate that pre-fibrillar Aβ can accelerate or even execute its neurotoxic effects via tau, the exact biochemical pathways by which it does so remain unknown.
The present study was designed to identify changes in pre-fibrillary protein species, associated with the accumulation of Aβ in the neuropil of the APPSWE/PS1ΔE9 (APP/PS1) mouse model of AD. APP/PS1 mice, overexpressing human chimeric amyloid precursor protein (AβPP) bearing the Swedish mutations (APP695SWE) and human presenilin 1 with exon 9 deletion (PSEN1DE9), show amyloid deposition in the cerebral cortex by the age of 3– 6 months [11]. As in patients with AD, amyloid pathology increases with age, reaching a plateau by 18 months of age [12, 13]. In addition, also the cerebellum is affected by amyloid pathology [14]. The model exhibits several additional characteristics of AD, including gliosis [12, 15], modest neuronal loss around plaques [16, 17], loss of monoaminergic afferents in the forebrain [18, 19], and memory deficits at 18 months of age [13, 20– 23]. Furthermore, a hyperphosphorylation of tau has also been observed in this model [24].
In order to study the effect of Aβ pathology on pre-fibrillary protein species [25], a sarkosyl-insoluble preparation was prepared from the brains of 3 and 24 month old APP/PS1 and littermate wild type (Wt) mice, and analyzed by quantitative liquid-chromatography tandem mass spectrometry (LC-MS/MS). The analysis showed differential, age- and genotype-induced effects on the sarkosyl-insoluble proteome of Wt and APP/PS1 mice. In addition to Aβ and tau, the sarkosyl insoluble material from the old transgenic mice showed significant enrichment in large networks of proteins involved in oxidative phosphorylation and mitochondrial function compared to control. These data implicate dysfunctional mitochondria as key mediators of Aβ- and tau-induced pathology in APP/PS1 mice. Furthermore, all identified proteins in the sarkosyl pellet were assigned cell type specificity based on RNAseq data published by Zhang et al. [26], indicating the majority of aggregated proteins as having a neuronal origin.
MATERIALS AND METHODS
Mouse brain tissue
Experiments were performed according to protocol 2011/561-1950, approved by the Danish Animal Ethics Inspectorate. Female, group-housed, 3- and 24-month-old APP/PS1 mice (B6.Cg-Tg (APPswe, PSEN1dE9)85Dbo/ Mmjax; line 85) [58] and littermate Wt mice were used in this study. The animals were bred and housed in the Biomedical Laboratory at the University of Southern Denmark, under a 12:12 h light:dark cycle (lights on 6:30 am). Food and water were available ad libitum. In total, 14 mice were killed by cervical dislocation (n = 3-4/genotype and age group), and their brains were rapidly removed and sectioned along the midline on a petri dish containing ice. The right half brain excluding the brainstem but including the cerebellum was frozen in isopentane on dry ice (– 30°C) and stored at – 80°C for further processing.
Preparation of sarkosyl-insoluble fractions
The Greenberg and Davies method [25] was used to isolate sarkosyl-insoluble proteins from the brain of 3 and 24 month old, Wt and APP/PS1 mice. The half brain including the cerebellum but excluding the brain stem (due to a lack of Aβ pathology in this region) from 3-4 animals/group were pooled and weighed, to obtain sufficient amount of material for the proteomics workflow (Fig. 1). The tissue was homogenized with a Potter-Elvehjem tissue grinder (WHEATON), in a 5-fold excess (v/w) of 10 mM Tris-HCl buffer (pH 7.4), containing 800 mM NaCl, 1 mM EGTA, 10% sucrose, protease inhibitors (cOmpleteTM Protease Inhibitor; Roche Diagnostics) and phosphatase inhibitors (PhosSTOPTM; Roche Diagnostics; H buffer). The homogenate was centrifuged at 4°C for 20 min in a refrigerated ultracentrifuge (27,000× g; Sorvall RC M150 GX). The supernatant was decanted and kept on ice, the pellet (P1) suspended in 5 vol of H-buffer and re-centrifuged at 27,000× g for 20 min (4°C). The combined supernatants (S1) were brought to 1% sarkosyl in H buffer and incubated for 2 h at 37°C in a C24 incubator shaker (100 rpm; New Brunswick Scientific). Following centrifugation at 200,000× g for 40 min (4°C), the sarkosyl-soluble fraction was decanted and kept on ice, and the sarkosyl-insoluble pellet suspended in H buffer, containing 1% CHAPS hydrate (Sigma Aldrich Co.). After filtering through acetate cellulose filters (0.45μm; VWR International), the filtrates were centrifuged at 200,000× g for 70 min, and the final pellet suspended in ∼250μL dH2O. Sarkosyl-insoluble fractions were stored at – 80°C for the proteomics study.
Mass spectrometry-based proteomics
Reduction, alkylation, and enzymatic digestion
Sarkosyl pellets were redissolved in 6 M urea, 2 M thiourea, and 10 mM DTT with protease and phosphatase inhibitors (cOmplete Protease inhibitors and PhospSTOP, Roche diagnostics) at room temperature (RT). After vortexing and sonication, 100μg total protein per condition was alkylated in 20 mM iodoacetamide (IAA) for 20 min in the dark (Fig. 1). A total of 2μl of endoproteinase Lys-C (6μg/μl, Wako) was added to the protein sample and the solution was incubated for 2 h at RT. The sample was diluted 10 times with 20 mM TEAB, pH 7.5 and digested with trypsin (1:50 (w/w) trypsin:protein) overnight. The enzymatic digestion was stopped with 5% formic acid (FA) and the peptide sample was cleared by centrifugation (14000× g, 15 min). Protein and peptide quantification was performed by fluorometric quantification (QubitTM – Life technologies).
iTRAQ-4plex labelling
A total of 100μg of tryptic peptides were dried and desalted with R2/R3 columns (as described under Sample desalting with R2/R3 micro-column) before iTRAQ-4plex labeling (AB Sciex). Labeling was performed as follows: iTRAQ-114 for Tg-3 months, iTRAQ-115 for Wt-3 months, iTRAQ-116 for Tg-24 months and iTRAQ-117 for Wt-24 months (Fig. 1). The labeling was performed according to manufacturer’s instruction. The labeled peptides from all groups were mixed 1:1:1:1, dried down and stored for further enrichment and analysis. During labeling, a pellet of ethanol-insoluble peptides was observed, which was collected from all samples and redissolved in 100% FA for 30 min. These samples were combined 1:1:1:1 and included as a separate sample in the following workflow.
Sample desalting with R2/R3 micro-column
The samples were desalted before fractionation by hydrophilic interaction chromatography (HILIC) using home-made P200-tip-based columns packed with equal ratios of Poros R2 (Oligo R2 Reversed Phase Resin 1-1112-46, Applied Biosystems) and Poros R3 (OligoR3 Reversed Phase Resin 1-1339-03, Applied Biosystems) reversed-phase resin material. The end of the tip was blocked with C8 material (Model 2314, 3 m EmporeTM C8). The column was prepared by short centrifugation (1000× g) of the R3 reversed-phase resin (100% ACN). The column was equilibrated with 0.1% TFA and centrifuged again. The acidified samples were loaded onto the columns and washed three times with 0.1% TFA. Peptides were eluted with 50% ACN, 0.1% TFA and dried by vacuum centrifugation.
HILIC fractionation
The fractions containing unmodified peptides were fractionated prior to nanoLC-MS/MS analysis using HILIC as described previously [27, 28] (Fig. 1). Peptides were dissolved in 90% ACN, 0.1% TFA (solvent B) and loaded onto a 450μm OD×320μm ID×17 cm micro-capillary column packed with TSK Amide-80 (3μm; Tosoh Bioscience) using an Agilent 1200 Series HPLC (Agilent). The peptides were separated using a gradient from 100– 60% solvent B (A = 0.1% TFA) in 30 min at a flow-rate of 6μl/min. Fractions were collected every 1 min based on the UV chromatogram. Subsequently, the peptide fractions were dried by vacuum centrifugation.
Reversed-phase nanoLC-ESI-MS/MS
The peptides (resuspended in 0.1% FA) were automatically injected and loaded on a ReproSil-Pur C18 AQ (Dr. Maisch, Ammerbuch-Entringen, Germany) in-house packed trap column (2 cm×100μm inner diameter; 5μm). The peptides were separated at 250 nl/min on an analytical ReproSil-Pur C18 AQ (Dr. Maisch, Ammerbuch-Entringen, Germany) packed in-house column (17 cm×75μm; 3μm) by reversed phase chromatography which was operated on an EASY-nanoLC system (Thermo Fisher Scientific, Odense, Denmark). Mobile phase was 95% ACN/ 0.1% FA (B) and water/0.1% FA (A). The gradient was from 1% to 30% solvent B in 80 min, 30– 50% B in 10 min, 50– 100% B in 5 min and 8 min at 100% B. The nano-LC was online connected to an Orbitrap Fusion Lumos mass spectrometer (Thermo Fisher Scientific) operating in positive ion mode and using data-dependent acquisition. The Orbitrap acquired the full MS scan with an automatic gain control (AGC) target value of 3×106 ions and a maximum fill time of 100 ms. Each MS scan was acquired at high-resolution (120,000 full-width half maximum (FWHM)) at m/z 200 in the Orbitrap with a mass range of 400– 1400 Da. The 12 most abundant peptide ions were selected from the MS for higher energy collision-induced dissociation (HCD) fragmentation (collision energy: 34 V) if they were at least doubly charged. Fragmentation was performed at high resolution (60,000 FWHM) for a target of 1×105 and a maximum injection time of 60 ms using an isolation window of 1.2 m/z and a dynamic exclusion of 20 s.
Data analysis
Raw data were searched against the Mus musculus reference databases from Swissprot and Uniprot via Mascot (v2.3.02, Matrix Science) and Sequest HT search engines, respectively, using Proteome Discoverer (v2.1, Thermo Fisher Scientific) (Fig. 1). A precursor mass tolerance of 10 ppm and a product ion mass tolerance of 0.02 Da were applied, allowing two missed cleavages for trypsin. Fixed modifications included carbamidomethylation of Cys and iTRAQ-4plex labelling for Lys and N-termini. The iTRAQ datasets were quantified using the centroid peak intensity with the “reporter ions quantifier” node. To ensure a high-confident identification of peptides, we used the Mascot percolator algorithm (q value filter set to 0.01), Mascot and Sequest HT peptide rank 1 and a cut-off value of Mascot score of ≥18 as well as Sequest HTΔCn of 0.1 and only high confident peptides were used for further analysis. Subsequently, these peptides were filtered against a decoy database resulting into a false discovery rate (FDR) to 0.01 (FDR <0.01). Significantly regulated proteins were determined based on the expression ratio being outside two standard deviations of the biological replicates thereby cut off values were set to 1.3 for upregulated and 0.75 for downregulated. The threshold criteria for the ratios were calculated on the average experimental technical variance of the multiple analysis of brain technical replications in mass spectrometry labeling analysis and has successfully been applied in proteomics studies using various mammalian brain regions [24, 29].
The mass spectrometry proteomics data have been deposited to the ProteomeXchange Consortium (http://proteomecentral.proteomexchange.org) via the PRIDE partner repository [30] with the dataset identifier <PXD009069 >.
Gene Ontology (GO) enrichment analysis
Sarkosyl insoluble proteomes were classified according to their GO terms using the Search Tool for the Retrieval of Interacting Genes/Proteins (STRING) database (version 10.5, confidence 0.9) and the Database for Annotation, Visualization and Integrated Discovery (DAVID) (version 6.8, NCBI) [31]. Proteins were categorized according to overrepresentation of a biological process, cellular component, molecular function and KEGG pathway. Due to the large numbers of regulated proteins, and to increase confidence, cut-off values of 2 and 0.4 were used for regulated proteins to be included in the GO analysis.
Cell type specific protein analysis
To find cell type specific proteins, proteomics data was compared to the RNAseq data set from Barres et al. as was done by Drummond et al. [26, 32]. Genes responsible for >60% of the total gene expression were considered cell type specific.
RESULTS
Characterization of the brain sarkosyl-insoluble proteome
The discovery of changes in the sarkosyl-insoluble proteome of APP/PS1 and littermate Wt mice was performed by iTRAQ-4plex labeling and off-line fractionation prior to LC-MS/MS. The iTRAQ labeling was performed on sarkosyl-insoluble half brains, mainly cerebral and cerebellar cortex fractions from APP/PS1 mice and Wt mice at 3 and 24 months of age. We quantified more than 2400 unique proteins with 2 or more unique peptides in the sarkosyl-insoluble fraction (Supplementary Table 1). The amyloid-β protein precursor (AβPP) was identified in the fraction with 4 unique peptides, herein peptides in the region of the Aβ42 peptide (amino acid 672– 713 of the AβPP protein sequence) [33]. In addition the tau protein (MAPT) was identified together with proteins involved in Aβ and tau binding and metabolism, such as clusterin [34], apolipoprotein E (APOE) [35], vimentin [36], cyclin-dependent kinase 5 (cdk5), glycogen synthase kinase-3 beta (gsk3β) [37] and succinate dehydrogenases [38]. All AD associated proteins are listed in Table 1, along with protein ratios for comparisons between the four experimental groups.
Known AD associated proteins in the sarkosyl-insoluble fraction (Colours are visible in the online version of this article https://dx-doi-org.web.bisu.edu.cn/10.3233/JAD-180197)
Red numbers indicate proteins in increased levels, green numbers indicate proteins in decreased levels, black numbers indicate unchanged levels of proteins. TG, APP/PS1; WT, Wild type; 24, 24 months old, 3, 3 months old.
When further analyzing the sarkosyl insoluble proteome in APP/PS1 and Wt mice using the enrichment of GO terms, biological processes such as translation, oxidation-reduction, RNA splicing, neurotransmitter secretion and actin cytoskeleton organization were observed to be enriched in the sarkosyl-insoluble fraction (Fig. 2A). The molecular functions of the enriched proteins were mainly associated with binding of RNA and actin in addition to binding of tau and Aβ (Fig. 2B). For the full analysis result also see Fig. 2C and D. Consistent with an involvement in AD pathogenesis, proteins involved in inflammation, kinase activity, synaptic plasticity as well as mitochondrial proteins involved in oxidative stress were identified.
To assess which cells contributed the most to the sarkosyl-insoluble proteome, proteins were assigned cell type specificity. For genes to be considered cell type specific, they needed to be responsible for >60% of the total gene expression. The identified proteins were analyzed for their cellular origin by comparison to RNAseq data of neurons, astrocytes, oligodendrocytes, microglia, and endothelial cells, purified by fluorescence assisted cell sorting from mouse cortices by Zhang et al. [26, 39]. The majority of proteins that could be assigned as cell type specific had a neuronal origin followed by oligodendrocytic and astrocytic origin (Table 2), indicating neurons to be more sensitive to Aβ pathology than glial cells. This was consistent with the GO-term analysis of enriched cellular compartments in which proteins mainly belonged to neuronal cell bodies, projections, synapses and myelin sheaths (Fig. 2C).
Cell-type specificity of proteins in the sarkosyl insoluble proteome
Proteins were mapped to the RNAseq dataset by Zhang et al. [26], assigning CNS cell-type specificity of genes. Tg, APP/PS1; Wt, wildtype; 24, 24 months old; 3, 3 months old.
Proteins were also analyzed for their involvement in KEGG pathways (Fig. 2D). Pathways significantly enriched included oxidative phosphorylation and neurological diseases, including AD, Parkinson’s disease (PD) and Huntington’s disease (HD), consistent with the notion that common molecular mechanisms may underlie the development of neurodegenerative proteinopathies. Furthermore, protein pathways involved in long-term potentiation (LTP) and long-term depression (LTD) were significantly enriched within the sarkosyl-insoluble proteome.
Age-related effect on the sarkosyl-insoluble proteome of Wt and APP/PS1 mice
To investigate the effect of aging on the aggregated proteins, within genotype, comparisons were performed at 3 versus 24 months of age (Table 3, Supplementary Table 1). Proteins were considered significantly differentially expressed when ratios were >2 SDs as well as >2-fold upregulated or <0.4 downregulated. In Wt mice, there were 69 significantly regulated proteins by age (Table 3), including mainly increased levels of IgG and histone proteins (Supplementary Table 1, Supplementary Figure 1). The aging effect on the sarkosyl-insoluble proteome was more pronounced in the APP/PS1 mice with 329 regulated proteins (Table 3, Supplementary Table 1). The difference in the number of regulated proteins demonstrated Aβ pathology to have a larger effect on the sarkosyl-insoluble proteome than aging. The overlap in regulated proteins between the two genotypes was approximately 4% and could be mainly attributed to age-specific changes in histones (Supplementary Figure 2). In addition, assigning cell type specificity to regulated proteins showed that neurons were the largest contributor to the sarkosyl-insoluble proteome in both APP/PS1 mice and Wt mice, suggesting neurons to be more sensitive to aging than other CNS cell types, and that Aβ pathology increases the sensitivity (Table 2). This indicated that the majority of the changes observed in the APP/PS1 proteome were due to the phenotype of the disease.

Sample preparation workflow for quantitative proteomics of sarkosyl-insoluble samples from APP/PS1 (Tg) and wild type (WT) mice at 3 and 24 months of age.
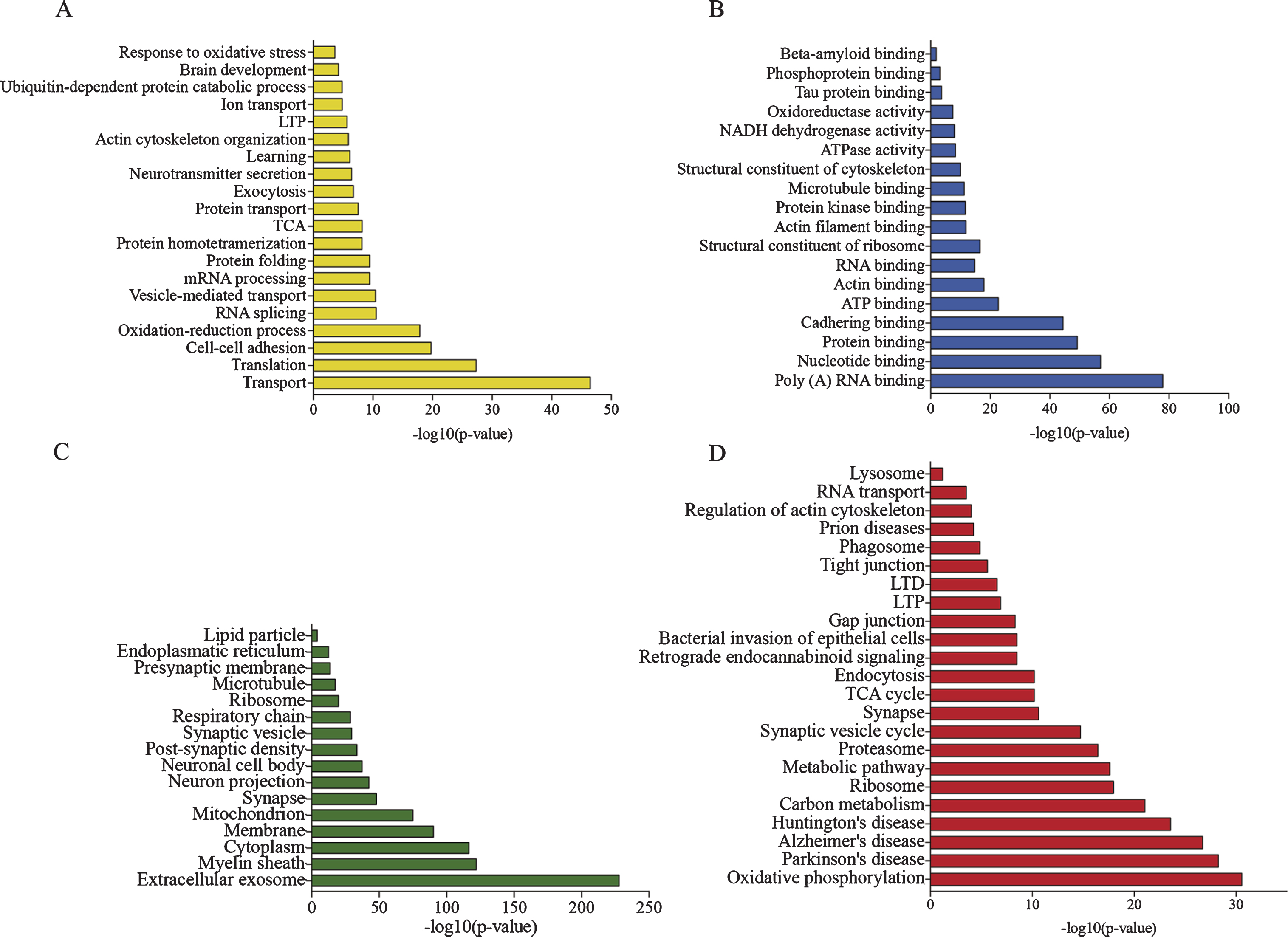
Characterization of the sarkosyl-insoluble proteome by GO-term enrichment of all identified proteins in the sarkosyl-insoluble fraction of all brain. A) Enriched biological processes. B) Enriched molecular functions. C) Enriched cellular compartments. D) Enriched KEGG pathways.
Numbers of significantly up- and downregulated proteins
Tg, APP/PS1; Wt, wildtype; 24, 24 months old; 3, 3 months old.
When analyzing the regulated proteins in 24 versus 3-month-old APP/PS1 mice, the majority of proteins identified were associated with oxidative and reduction processes, including large numbers of NADH dehydrogenases, cytochromes and ion transporting proteins. The GO term analysis of enriched KEGG pathways included oxidative phosphorylation, AD, PD and HD, synaptic vesicle cycle and phagosomes (Fig. 3A). The GO term analysis of biological processes showed that transporter activities, ATP-dependent metabolic processes, as well as oxidative stress, responses to oxidative stress and apoptotic mitochondrial processes were significantly enriched (Fig. 3B). The molecular function of the proteins referred to binding of misfolded proteins and ribosomal constituents (Fig. 3C). When analyzing the cell-type specific proteins, the majority of the regulated proteins had neuronal origin, compatible with the cellular component enrichment analysis (Figs. 3D, 4).
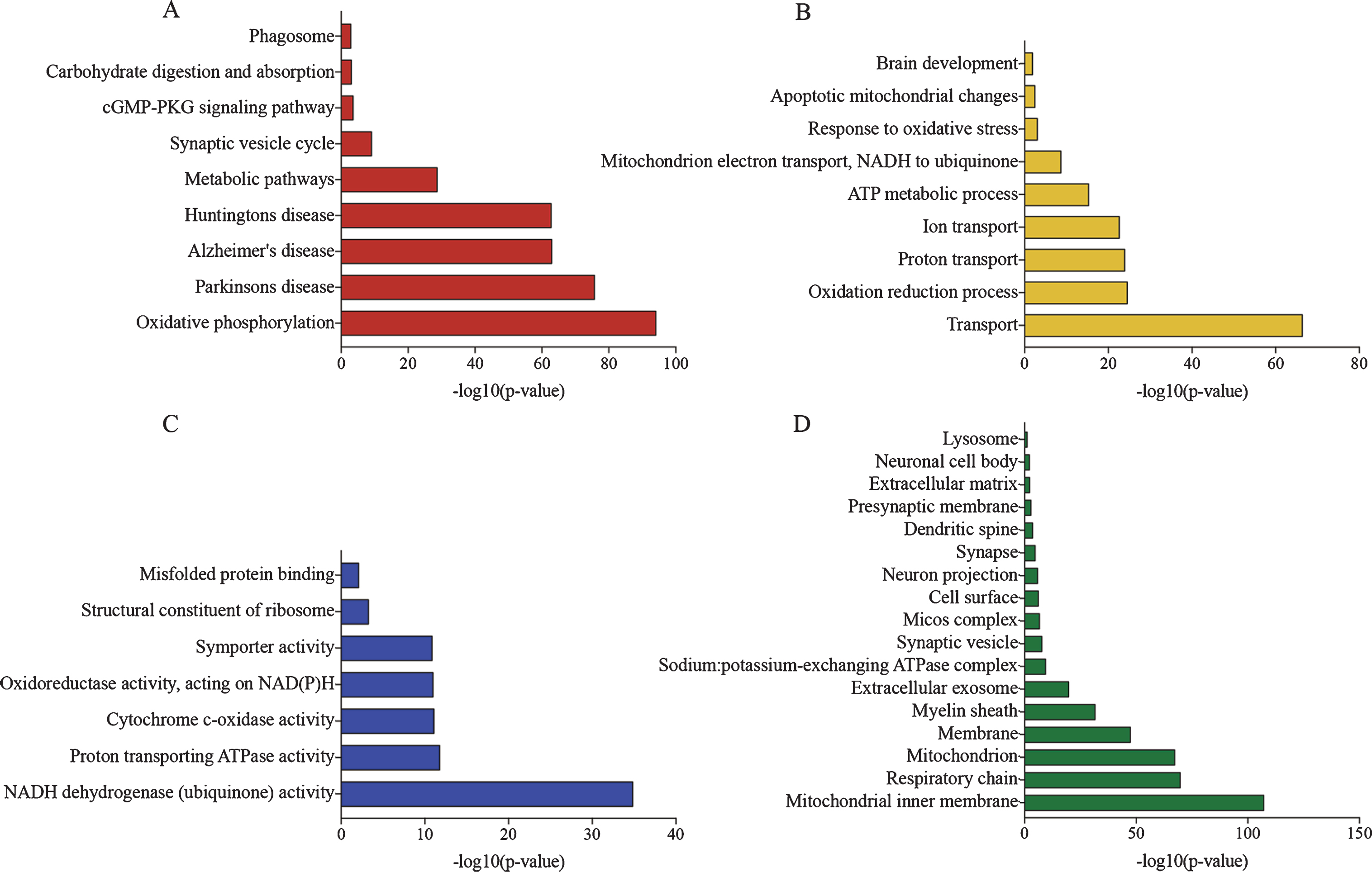
GO-term enrichment of regulated proteins in APP/PS1 24 months compared to APP/PS1 3 months old mice. A) Enrichment of KEGG pathways. B) Enriched biological processes. C) Enrichment of molecular function. D) Enrichment of cellular compartment.
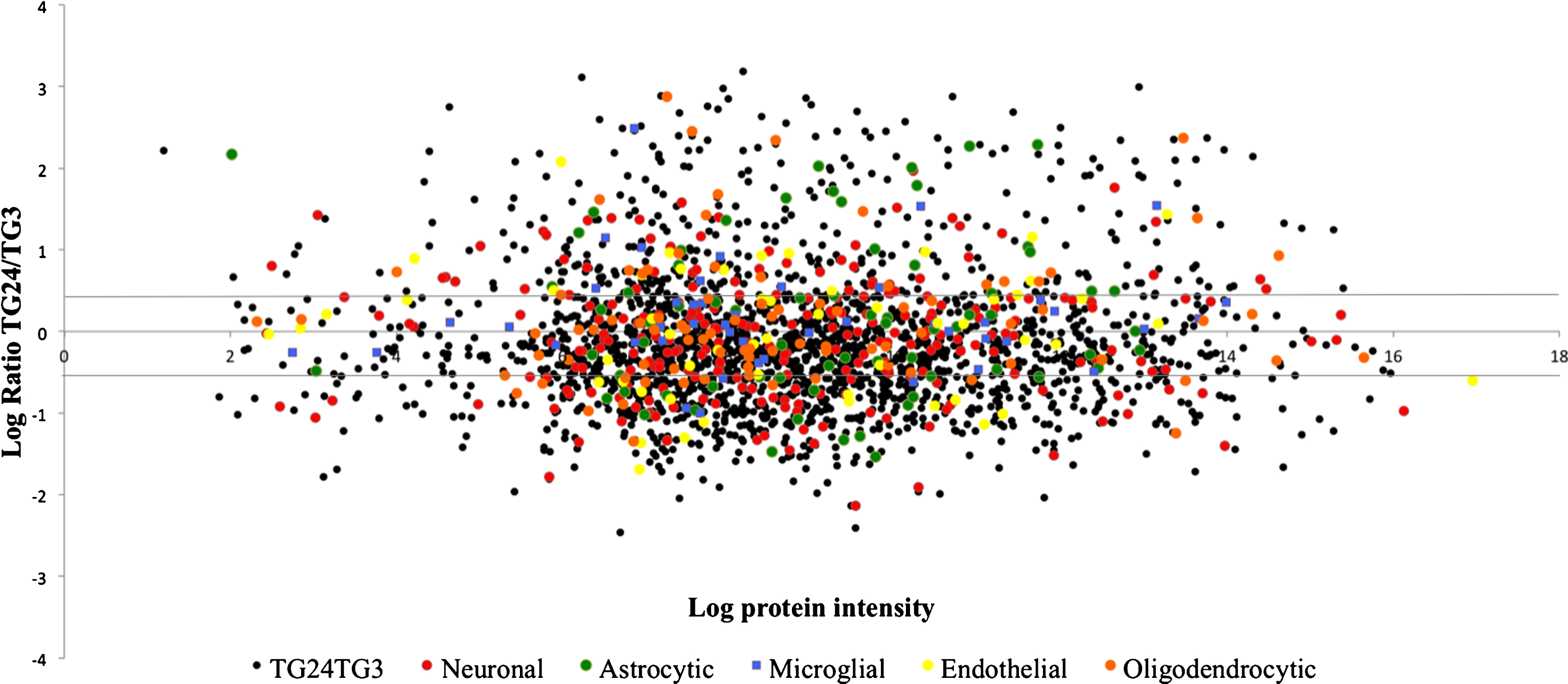
Cellular origin of proteins. Proteins were ascribed cell-type specificity according to Zhang et al. [26]. Black dots: all quantified proteins, red: neuronal proteins, green dots: astrocytic proteins, blue dots: microglial proteins, orange dots: oligodendrocytic proteins, yellow dots: endothelial proteins. Lines indicate limits for significantly regulated proteins on the log scale, indicating mainly neuronal proteins to be accumulated in the sarkosyl-insoluble fraction of the proteome.
Genotype-induced effects at 3 and 24 months of age
To investigate disease-associated changes of the sarkosyl-insoluble proteome, age-matched Wt and APP/PS1 mice were compared (Table 3). At the age of 3 months the changes observed between APP/PS1 and Wt mice involved the significant regulation of 51 proteins (Table 3). The differences were more pronounced at the age of 24 months, when 211 proteins were differentially regulated between genotypes (Table 3). At 3 months of age the regulated proteins included mostly DNA and RNA binding proteins as well as actin cytoskeletal proteins with the majority being involved in regulating gene expression (Fig. 5A-C). At 24 months of age, the proteins that were increased in the detergent insoluble fraction from APP/PS1 mice were mainly involved in mitochondrial function and pathways such as oxidative phosphorylation and oxidative stress, as well as phagosomal proteins and proteins involved in the function of synapses (Fig. 5D-G, Supplementary Figure 3).
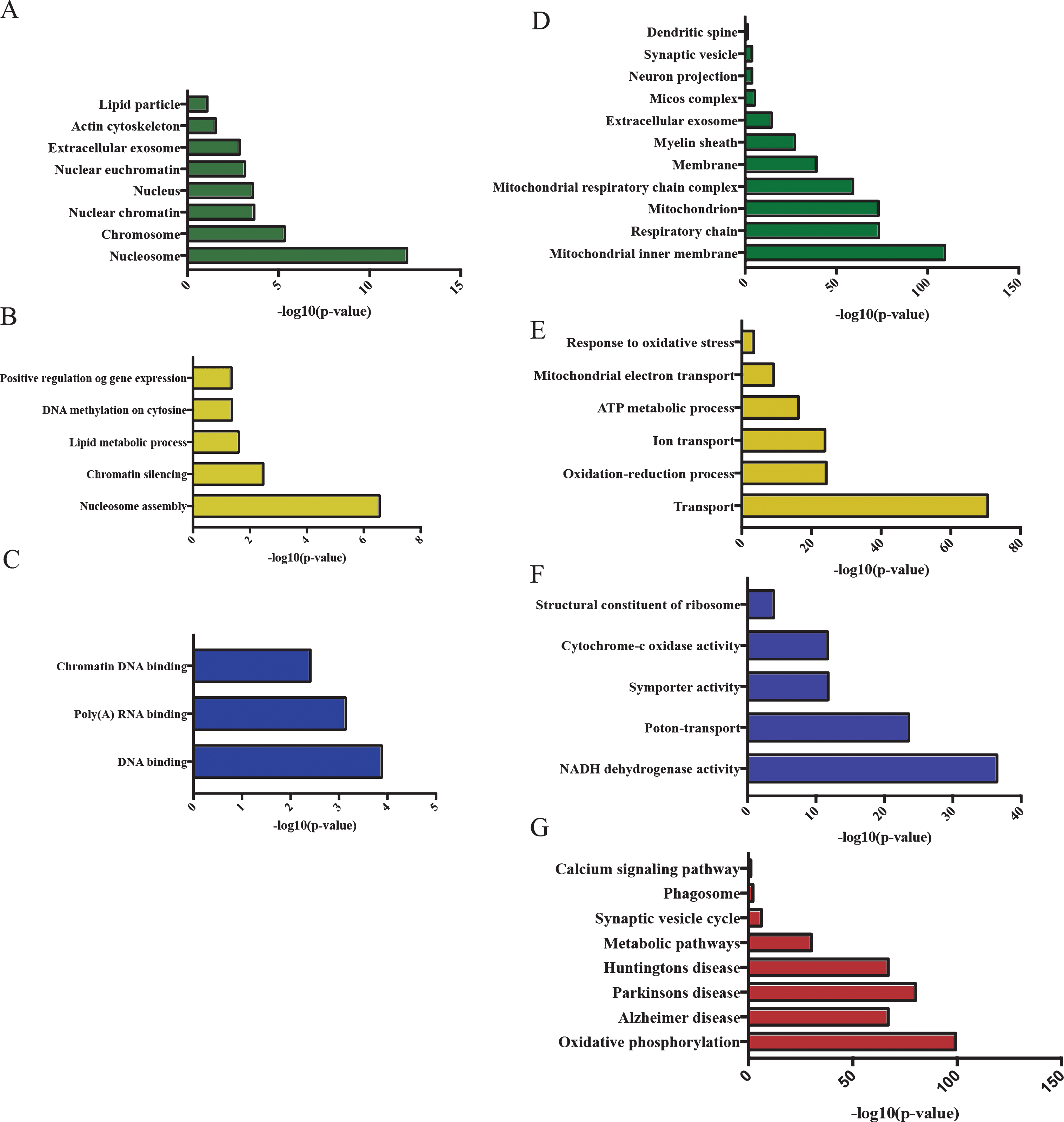
GO term enrichment of regulated proteins in APP/PS1-3months relative to Wt-3months (A-C) and in APP/PS1-24months relative to Wt-24 months (D-G). A, D) Enriched cellular compartments. B, E) Enriched biological processes. C, F) Enriched molecular function. G) Enriched KEGG pathways.
DISCUSSION
In this study proteomics employing iTRAQ-4plex labeling was used to investigate the sarkosyl-insoluble proteome of APP/PS1 mice from young adults with no apparent Aβ pathology (3 months) to aged APP/PS1 mice with severe Aβ pathology (24 months of age). Comparisons were performed to Wt littermate mice to subtract changes in the proteome that could be ascribed to age.
More than 2400 proteins were quantified in the sarkosyl-insoluble proteome and both AβPP and tau were identified with the highest level of AβPP in the APP/PS1 mice at 24 months of age. Due to Aβ42 being a proteolysis product of AβPP, it does not have its own entry in the databases used to search the proteomics data. Thus, peptides spanning the Aβ42 region will be mapped to the AβPP protein. In our data we quantified peptides spanning both the Aβ42 and the AβPP sequence thereby indicating both Aβ42 and AβPP to be present in the sarkosyl-insoluble proteome of the APP/PS1 mice. Furthermore, proteins previously associated with AD, such as APOE, clusterin, vimentin, cdk5 and gsk3β were identified in the sarkosyl-insoluble fraction of the proteome associated with the development of plaques.
A recent study by Hales et al. investigated the brain sarkosyl-insoluble fraction of patients with sporadic AD and identified approximately 2700 proteins using LC-MS/MS techniques [40]. When comparing the sarkosyl-insoluble proteome of APP/PS1 mice to the sarkosyl-insoluble proteome of AD cases a >50% overlap between the two data sets was observed (Fig. 6 and Supplementary Table 1, list of overlapping proteins). When mapping only the significantly regulated proteins from AD, mild cognitive impairment (MCI) and asymptomatic AD to our dataset, most of the proteins in AD and MCI were identified, with the largest percentage of the mapped proteins from each condition belonging to the MCI state. This indicates that the APP/PS1 mouse model shares many of the characteristics with sporadic AD, and especially with MCI.
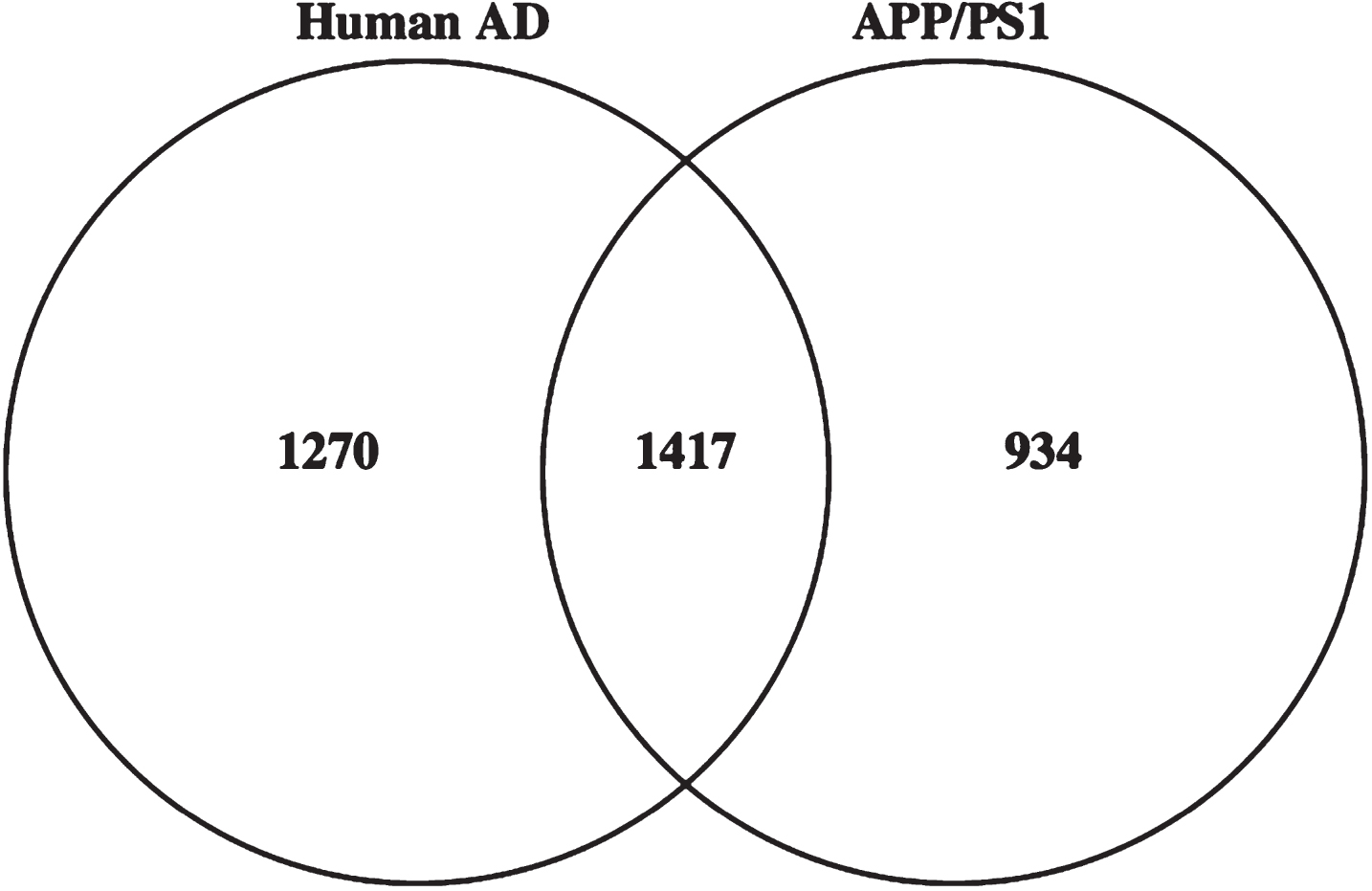
Comparison of APP/PS1 sarkosyl-insoluble proteome to sarkosyl-insoluble proteome from sporadic AD cases. The Venn diagram showed more than 50% overlap between the sarkosyl-insoluble proteome of sporadic AD cases and APP/PS1 mice. The datasets on sporadic AD were obtained from Hales et al. [40].
Despite the young age of the youngest studied mice, 51 significantly differentially expressed proteins were found between the APP/PS1 and Wt sarkosyl-insoluble proteome, including proteins regulating gene expression. Among these, especially histones were enriched. Histones and changes in acetylation patterns of these have previously been linked to memory consolidation and neurodegenerative diseases [41]. In addition, histone deacetylation inhibitors have been indicated to stimulate learning abilities and memory in the APP/PS1 mouse as well as other amyloidosis models [42, 43]. The fact that changes in these proteins are seen as early as 3 months could be of interest in the understanding of early disease mechanisms of AD as well as providing potential therapeutic targets.
At 24 months of age the number of regulated proteins between APP/PS1 and Wt insoluble proteomes increased to 211, emphasizing the development of pronounced differences between APP/PS1 and Wt mice as they age. Here, significantly regulated proteins included a large proportion of NADH dehydrogenases, cytochromes and other proteins involved in oxidative phosphorylation pathways amongst the highest regulated. Mitochondria are known to play a central role in apoptosis and maintenance of cellular function through the generation of ATP, and mitochondrial dysfunction appears to impact on the pathogenesis of neurodegenerative diseases, such as PD and HD [44, 45]. Morphological, biochemical and genetic abnormalities of mitochondria have been observed in AD, laying the ground to a mitochondrial cascade hypothesis [46–48]. Morphological changes of mitochondria have also been observed by electron microscopy in the APP/PS1 mouse starting at 3 months of age, thereby preceding the onset of behavioral changes in this model [49]. That aggregation of mitochondrial proteins takes place in the APP/PS1 mouse is interesting based on the lack of overt degeneration of cortical neurons in this model [12], suggesting that mitochondrial proteins turn insoluble while still present intracellularly. This is supported by the fact that aging and hypoxic conditions can induce changes in the solubility of mitochondrial proteins [50]. This mechanistic view could be readily transferred to AD, where there is overlap in dementia caused by AD and cerebrovascular diseases, spanning small vessel disease to post-stroke dementia [51].
More than 300 proteins were significantly regulated between the sarkosyl-insoluble proteome of APP/PS1 mice of 24 months and 3 months of age. Only 4% of these proteins were found regulated in the Wt mice indicating the majority of regulated proteins to be related to disease development. The disease related changes in proteins showed the same pattern of mitochondrial dysfunction, oxidative stress as well as ion transporting proteins as mentioned earlier, further indicating mitochondrial dysfunction and energy metabolism to be affected in this model. Furthermore, a large proportion of RNA processing proteins were found significantly enriched in the sarkosyl-insoluble proteome, a category of proteins previously found associated with plaques and tangles in AD [40], as well as being present in sarkosyl-insoluble fractions of AD and chronic traumatic encephalopathy [40, 53].
The majority of proteins assigned as cell type specific had neuronal origin, supported by the cellular component GO analysis showing neuronal projection, synapse, and cell body proteins to be enriched. This indicates possible degeneration of synapses in the APP/PS1 model, perhaps induced by oxidative stress pathways initiated by the accumulation of Aβ, explaining why these proteins would accumulate in the sarkosyl-insoluble fraction of the brain [24]. A further possibility could be apoptotic events of microglia, astrocytes, or oligodendrocytes, which would contribute to the aggregation of intracellular proteins, including those in mitochondria.
In this study we have found protein changes, which are directly associated to the genotype of the APP/PS1 mouse and shared with human sporadic AD including the AD risk factors APOE and Clu. Furthermore, large networks of mitochondrial proteins were found accumulating in the sarkosyl-insoluble fraction of the proteome, suggesting that oxidative stress is a major contributor to AD pathogenesis.