
Abstract
Besides the typical cognitive decline, patients with Alzheimer’s disease (AD) develop disorders of the respiratory system, such as sleep apnea, shortness of breath, and arrhythmias. These symptoms are aggravated with the progression of the disease. However, the cause and nature of these disturbances are not well understood. Here, we treated animals with intracerebroventricular streptozotocin (STZ, 2 mg/kg), a drug that has been described to cause Alzheimer-like behavioral and histopathological impairments. We measured ventilation (
INTRODUCTION
Alzheimer’s disease (AD) is a neurodegenerative disorder, often associated with age, whose cognitive and neuropsychiatric manifestations result in a progressive impairment and eventual disability [1]. AD is the most common cause of dementia, affecting more than 45 million people worldwide [2]. This disease is characterized by the accumulation of amyloid-β (Aβ) plaques, intracellular neurofibrillary tangles (NFTs) of hyperphosphorylated tau protein, and decreased synaptic density, which eventually leads to widespread neurodegeneration and loss of neurotransmitters [3]. The neurodegeneration occurs in multiple areas of the brain, including the hippocampus, cortex, amygdala, neocortex, and brainstem structures [4–8].
AD is a multifactorial, neurodegenerative, age-related disorder with multiple components involved in its progression [9]. Therefore, the establishment of animal models that reproduce human pathology becomes complex. Despite this, the use of animal models has been crucial in defining critical disease-related mechanisms and has been at the forefront of evaluating novel therapeutic approaches [10]. An animal model that replicates many behavioral and histological aspects of AD was created by the intracerebroventricular (icv) injection of streptozotocin (STZ) [11–14]. This drug promotes a decrement in brain glucose/energy metabolism [11], which is considered an incipient sign of AD [11–14], and have been used to induce AD-like neurodegeneration in rats, since the metabolic abnormalities found in this model resemble those found in sporadic AD. For instance, these animals display progressive loss of memory and learning, increased brain ventricular volume, abnormalities in mitochondrial function, increased tau phosphorylation and increased Aβ peptide in brain areas [15–17]. These morphological changes result in damage and loss of neuronal cells [18]. Therefore, the use of the icv STZ model represents a viable experimental approach to explore changes involved in AD-like neurodegeneration in rats.
Aside from severe cognitive deficits, epidemiological studies have reported that up to 45% of patients with AD have sleep disturbances, such as increasing sleep fragmentation, nighttime awakenings, and a greater tendency for daytime sleep [19, 20]. These symptoms may occur at early stages of AD, but seem to be correlated with a more severe cognitive decline [21, 22]. In addition, it is known that over 70% of institutionalized patients with AD exhibit respiratory changes, and that these problems can contribute to cognitive decline [23–26]. According to Gaig et al. [25], obstructive sleep apnea may worsen or cause cognitive impairment in AD. Furthermore, there is a strong correlation of the severity of sleep-disordered breathing with the severity of AD [26]. The late stages of clinical AD are accompanied by shortness of breath [27]. This evidence shows that regions responsible for respiratory control are possibly changed.
There is a paucity of data in the literature on the study of respiratory control in AD progression. In this scenario, the use of animals as models are valid and essential in AD-related research, as they allow the assessment of early or late pathophysiological processes that are not accessible in human patients. Recently, Ebel et al. [28] demonstrated that icv STZ induces increased respiration at rest and blunted peripheral chemoreflex responses and a small change in the CO2-drive to breathe. However, the authors did not evaluate the respiratory pattern and ventilatory responses to hypoxic and hypercapnic stimuli during the different phases of the sleep/wake cycle, which was carried out in the present study. This is a crucial point, since the prevalence of sleep apnea in AD patients is high, with 70% to 80% of patients presenting five or more apnea-hypopnea episodes per hour of sleep, and 38% to 48% of individuals with 20 or more episodes of apnea during the sleep phase [27]. Thus, it is quite likely that there is a difference in the chemosensitivity of patients with the AD-dependent phase of the sleep/wake cycle. Therefore, we used the STZ model in order to induce AD-like neurodegeneration in rats to determine the ventilatory response to hypoxia and hypercapnia during wakefulness and non-rapid eye movement (NREM) sleep. In addition, we performed western blot analyses for phosphorylated tau, total tau and amyloid-β peptide (Aβ) in locus coeruleus (LC), retrotrapezoid nucleus (RTN), medullary raphe (Raphe), pre-Bötzinger/Bötzinger complex (PreBötz/BötzC) and hippocampus (HPC), and evaluated memory and learning acquisition using the Barnes maze.
MATERIAL AND METHODS
Animals
Male Wistar rats (3-4 months old; weight 300–350 g) were housed in a temperature-controlled chamber maintained at 24–26°C (ALE 9902001; Alesco Ltda., Monte Mor, SP, Brazil) with a 12:12-h light/dark cycle, and had free access to water and food. The experiments were performed between 08.00 a.m. and 17.00 p.m.
The study was conducted in compliance with the guidelines of the National Council for the Control of Animal Experimentation (CONCEA, MCT, Brazil) and with the approval of the Faculty of Agricultural and Veterinary Sciences and Animal Care and Use Committee (CEUA, FACV-UNESP, Jaboticabal campus; Protocol no. n° 6.030/016).
Surgical procedures
All surgical procedures were performed under anesthesia with an intraperitoneal injection of 100 mg/kg of ketamine (Union National Pharmaceutical Chemistry S/A, Embu-Guaçu, SP, Brazil) and 10 mg/kg of xylazine (Laboratories Calier S/A Barcelona, Spain). Postoperatively, the animals were treated with antibiotic (enrofloxacin, 10 mg/kg, intramuscular) and analgesic (flunixin meglumine, 2.5 mg/kg, subcutaneous) agents.
Intracerebroventricular injection of streptozotocin
The head was shaved and the skin was sterilized with betadine solution and alcohol. The rats were fixed to a Kopf stereotaxic apparatus (David Kopf Instruments, Tujunga, CA, USA). The scalp was incised over the sagittal suture, the periosteum was excised, and two small bilateral orifices were made using a sterilized dental drill to access both lateral ventricles of the brain, where icv injections were to be applied. The following coordinates were used: –0.8 mm posterior,±1.4 mm lateral, and –4.4 below bregma [29, 30]. Animals received bilateral icv injections of STZ (2 mg/kg dissolved in 0.05 mol/L citrate buffer, pH 4.5; Sigma, St. Louis, MO) or citrate buffer (2μL/ventricle) [31]. For the microinjection of drug and vehicle, a 5-μL Hamilton syringe was used, linked to a PE 10 polyethylene tubing, connected to a gengival needle for the application [30]. Thirty days after icv injection, rats were submitted to the behavioral, respiratory, or molecular tests. The respiratory tests were performed in a different group of animals, whereas the behavioral and molecular tests were performed in the same group.
Electroencephalogram (EEG) and electromyogram (EMG) electrodes
Seven days before the beginning of the respiratory tests, EEG and EMG electrodes were implanted in the group that would undergo the respiratory tests. Three EEG electrodes were introduced: the frontal electrode, located 2 mm anterior to bregma and 2 mm lateral to the midline; the parietal electrode, positioned 4 mm anterior to the lambda and 2 mm lateral to the midline; and the electrode “ground” which was inserted between the frontal and parietal electrodes. For EMG recordings, a pair of electrodes were inserted deep into the neck musculature of the rats. These electrodes allowed us to analyze ventilation in the different phases of the sleep/wake cycle.
Body temperature
On the same day as the EEG and EMG surgery, the rats underwent a second surgery for the implantation of a temperature datalogger (SubCue Dataloggers, Calgary, Canada) into the abdominal cavity through a midline laparotomy. The datalogger was programmed to acquire body temperature (Tb) data every 7 min.
Behavioral analysis
Barnes maze
The Barnes maze test was used to assess spatial memory and learning to confirm the success and reliability of the model.
The Barnes maze test consists of a non-aquatic test for memory and spatial learning [32]. The protocol was adapted from Sunyer et al. [33]. The maze was made from a circular, 13-mm thick, white PVC platform (110-cm diameter), which was maintained in the same position throughout the experiment. Twenty holes (10 cm in diameter, 7.5 cm between each hole) were made on the perimeter and the platform was mounted on top of a metal support, 105 cm above the ground. The maze was divided into target hole (T), opposite hole (OP), 9 holes clockwise (1 to 9, counting from T), and 9 holes counterclockwise (–1 to –9, counting from T). The escape cage, below the T hole, had walls covered with black plastic to make the inside dark and attractive to the rats. The platform was also illuminated with a fluorescent white light and visual cues (colored geometric figures) were placed around the labyrinth as points of spatial reference for the animal to escape from the open platform to the “target hole” These clues were not removed throughout the experiment. After testing each rat, as explained below, the whole maze was cleaned using 10% ethanol to avoid olfactory cues and the platform was rotated to avoid intra-maze odor or visual cues. All sessions were recorded by a video camera for further analysis. Prior to testing (probe day), training sessions were performed over 5 days to familiarize the animals with the maze and to allow them learn the location of the escape zone.
Acquisition phase
In this stage, spatial learning was evaluated through latency to enter the target hole. From days 1 to 4, each animal was placed in a square start chamber in the center of the maze. After 10 s, the chamber was removed and the animal was allowed to explore the maze for 3 min. The end of the trial was considered when the animal entered the target hole. Therefore, the animal was allowed to stay inside the dark cage for 1 min and, after this period, was returned to its home cage. If the animal had not entered in the target hole after 3 min from the beginning of the test, it was gently coerced to the target hole. These steps were repeated for 4 trials per day with an intertrial interval of 15 min over 4 consecutive days.
Probe trial
On day 5, 24 h after the last training day, the escape cage was removed, while the maze was maintained in the same position as training days. The animal was placed in the start chamber in the center of the maze and, after 10 s, it was removed and the animal was allowed to explore the maze for 90 s. The probe trial was performed in order to determine whether the animals remembered the location of the target hole. The number of times the animal searched for the “target hole” was used as a memory consolidation index [33–35].
Respiration test
Determination of pulmonary ventilation
We used the whole-body plethysmography method to measure of pulmonary ventilation (
VT was calculated by using an appropriate formula [37]:
EEG and EMG signals
Similar to other previous studies, the arousal state was determined by analyzing the EEG and EMG records [40, 42]. The signals from the EEG and EMG electrodes were sampled at 1 kHz, filtered at 0.3–50 and 0.1–100 Hz, respectively, and recorded on a computer. Both wakefulness and NREM sleep states were observed consistently through the experiments, but periods of rapid eye movement (REM) sleep were short and were not present in every experiment; thus, REM sleep phases were excluded from the analysis [40].
Experimental protocol
Seven days after implantation of EEG and EMG electrodes, the animals were placed in a plethysmographic chamber and ventilation was measured. The chamber was initially ventilated with moist atmospheric air (21% O2) for an acclimation phase of at least 30 min. Ventilation control measures were then performed. Ventilation measurements were first made during normocapnia for 63 min. The animals were then submitted to hypercapnia for 63 min, where the chamber was ventilated with a gas mixture containing 7% CO2, 21% O2 and balanced with N2 (White Martins Gases Industriais Ltda, Osasco, SP). After hypercapnia, the chamber was ventilated with atmospheric air again for recovery of baseline ventilation for 60 min. The animals were then submitted to hypoxia for 63 min, where the chamber was ventilated with a gas mixture containing 10% O2 and equilibrated with N2 (White Martins Gases Industriais Ltda, Osasco, SP). The order of exposure to hypercapnia and hypoxia was reversed randomly. The ventilatory measures were analyzed at 7, 14, 21, 28, 35, 42, 49, 56, and 63 min after gas exposure. Then, based on the sleep/wake cycle, for the
Neurochemical analysis
Brain area dissection
After each battery of behavioral tests, four rats were randomly selected out of each group and the animals’ brains were removed quickly, frozen in liquid nitrogen, and held at –80°C until dissection. In a cryostat at –22°C, the brains were coronary-sectioned to find target areas according to stereotaxic coordinates of the Atlas de Paxinos and Watson [43] as follow: locus coeruleus (LC - distance from bregma: –10.3 mm to –9.3 mm), retrotrapezoid nucleus (RTN, –11.3 mm to –10.3 mm), medullary raphe (–11.6 mm to –10.3 mm); pre-Bötzinger and Bötzinger complex (PreBötz/BötzC, –12.3 mm to –11.8 mm) and hippocampus (HPC, –4.16 mm to –3.16 mm). Samples of 0.7-mm thickness were removed with a 15-gauge needle.
Immunoblotting
Phosphorylated tau protein levels (Ser199/202 and Ser396) and Aβ peptide in LC, RTN, Raphe, PreBötzC/BötzC, and HPC were evaluated.
For this procedure, a maceration of the tissue of each corresponding region was performed. This tissue sample was submitted to sonication to promote membrane lysis and eventual release of the intracellular material. The solution containing the lysed cells was collected and transferred to an Eppendorf tube. Then, the quantification of these proteins was performed by the Biochrom UV/Vis spectrophotometer, Biodrop Duo model, UV optical path.
Before running the gel, the samples were denatured at 100°C for 3 min and applied to a polyacrylamide gel (12%) for fractionation. As a control, 8μL of a molecular weight marker was applied to one well. The samples were applied to the gel, placed in a vial with running buffer and separated by applying 100 V for 2 h. After the run, proteins were transferred to a nitrocellulose membrane (Bio-Rad) using ice-cold transfer buffer for 1 h at 100 V. The membrane was then incubated with non-specific site blocking solution (5% milk in TBS-T) for 1 h at room temperature. After this blocking, the membranes were cut and incubated with the respective primary antibodies, Anti-phospho-Tau (rabbit monoclonal pSer199/202, cat. T6819, Sigma,-Aldrich, 1:1000 dilution and rabbit monoclonal pSer396, cat. EPR2731-Ab109390, Abcam, 1:1000 dilution), anti-beta-actin (mouse monoclonal beta-actin antibody AC-15, cat. NB600-501, Novus Biological, 1:1000 dilution), anti-beta amyloid (mouse monoclonal β-amyloid B-4, cat. sc-28365, Santa Cruz Biotechnology, 1:500 dilution), and anti-Tau Total (rabbit polyclonal human tau, Ab-356, cat. GWB-ASC840, GenWay, 1:1000 dilution), diluted in the same blocking solution for 24 h at 4°C under constant stirring. The beta-actin antibody was used to normalize the specific labeling values. After washing, the corresponding secondary (anti-mouse IgG and anti-rabbit, 1:1000 dilution, Jackson ImmunoResearch) antibodies were incubated for 2 h at room temperature. The membranes were then washed twice with TBS-T and once with TBS for 10 min each, and the reaction was carried out by incubation with a chemiluminescent reagent for 1 min. The membranes were immediately exposed to chemiluminescence-sensitive film for 30 s to 5 min, as instructed by the manufacturer.
The films were quantified through optical densitometry using a computerized image analysis system (NIH System, ImageJ developed at the US National Institute of Health, available at the website: http://www.rsb.info.nih.gov/nihimage/(ImageJ).
Statistical analyses
Results were expressed as mean±SEM. The Barnes maze results in the acquisition phase were evaluated by two-way ANOVA, followed by the Holm-Sidak post-test. The Barnes maze, on the day of the test, was evaluated by the two-way ANOVA. The results of ventilation, sleep/wake cycle and body temperature in normocapnic, hypercapnic, and hypoxic conditions were evaluated by two-way ANOVA, followed by Bonferroni post-test. The quantification of phosphorylation of tau protein, total tau and Aβ were validated through the t-test (Student) for independent samples. The significance level adopted for all results was p < 0.05.
RESULTS
Behavioral analysis for model validation
Barnes maze: Acquisition phase
Figure 1A represents the latency value (s) to find the escape box in the target hole in training 4 for 4 days (acquisition phase). We observed that the control group decreased the latency (s) to escape through the target hole as a function of training (day 1: T1 134.3±54.2 versus day 4: T4 29.0±35.1 s, p < 0.05; two-way ANOVA). However, the STZ group required more time to find the escape hole compared to the control group on days 1, 2 and 3 (p < 0.05; two-way ANOVA). No difference between treatments was observed on the fourth day. On the same day, the STZ group decreased latency, thus improving performance on the platform to find the escape box (day 1: T1 165.8±38.9 versus day 4: T4 39.8±62.4 s, p < 0.05; two-way ANOVA).

Effect of icv STZ (2 mg/kg) on spatial learning and memory retention in the Barnes maze test. A) Time to reach the target hole in seconds of the control and STZ groups in the 4 training sessions per day (1st to 4th day). B) Number of attempts to enter into the holes in the platform test pattern in the control and STZ group. Values are expressed as mean±SEM. * indicates a significant difference between the control and STZ groups.
Barnes maze: Proof test
Figure 1B shows the number of attempts to enter the holes in the platform test pattern. In the Barnes maze test, the STZ-treated group had a lower number of hits in the “target hole” compared to the control group (control: 4.5±1.8 versus STZ: 2.0±2.2, respectively, p = 0.035; two-way ANOVA). There was no significant difference between treatments in the other holes of the platform (p > 0.05; two-way ANOVA).
Respiration test
During all experimental protocols, the mean chamber temperature was 25.7±0.3°C, and the mean room temperature was 24.6±0.2°C.
Respiration during wakefulness
We observed no difference in respiratory variables in room air conditions in the STZ-treated group compared to the control group during wakefulness (Figs. 3A and 4A). In hypercapnia and hypoxia, all groups showed a significant increase in ventilation when compared to room air conditions (p < 0.0001; two-way ANOVA) (Figs. 2A and 3A).
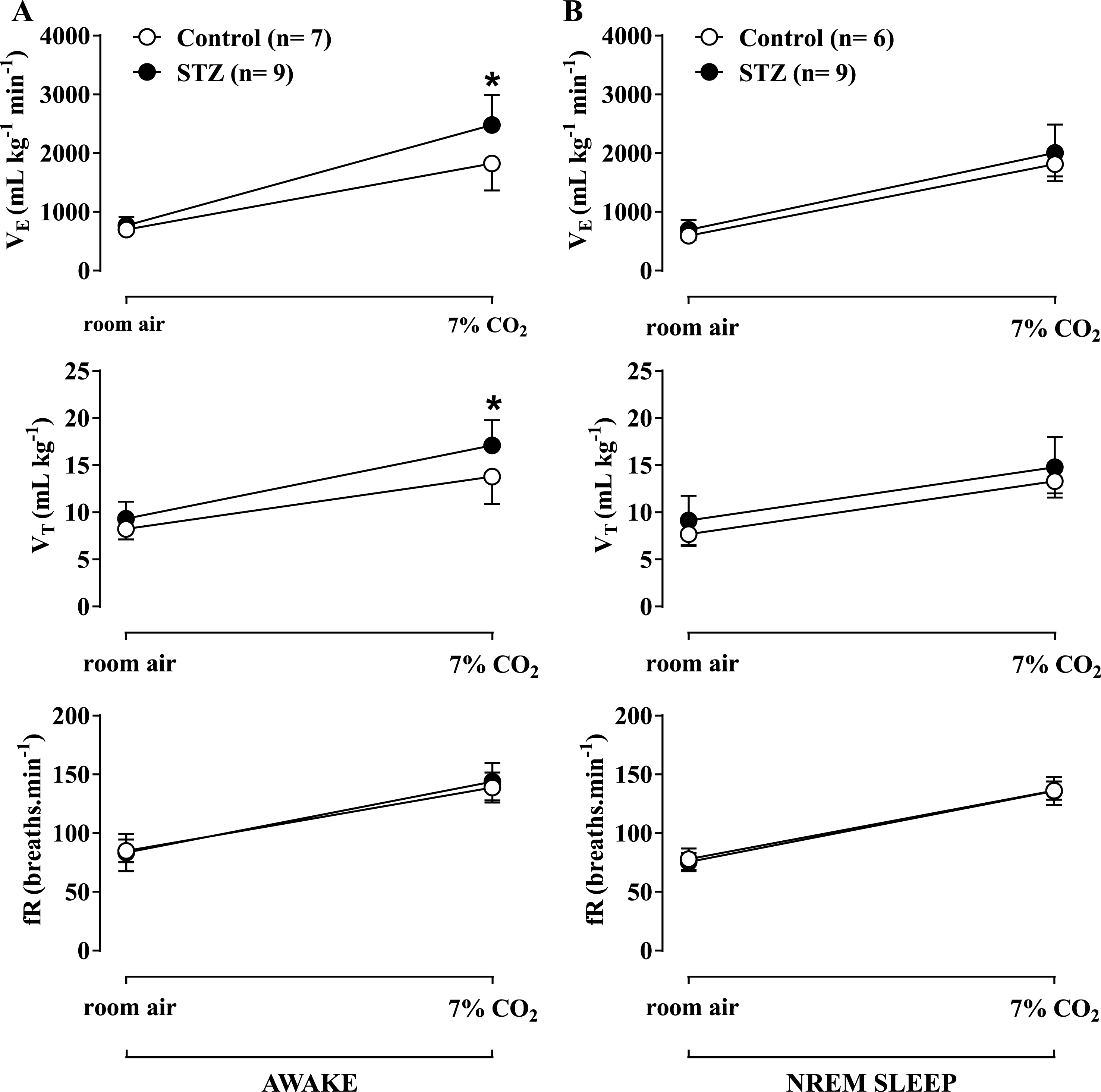
Effect of icv STZ (2 mg/kg) on ventilation (VE), tidal volume (VT) and respiratory frequency (fR) during hypercapnic exposure (7% CO2) during wakefulness (A) and NREM sleep (B). Values are expressed as mean±SEM. * indicates a significant difference between the control and STZ groups.
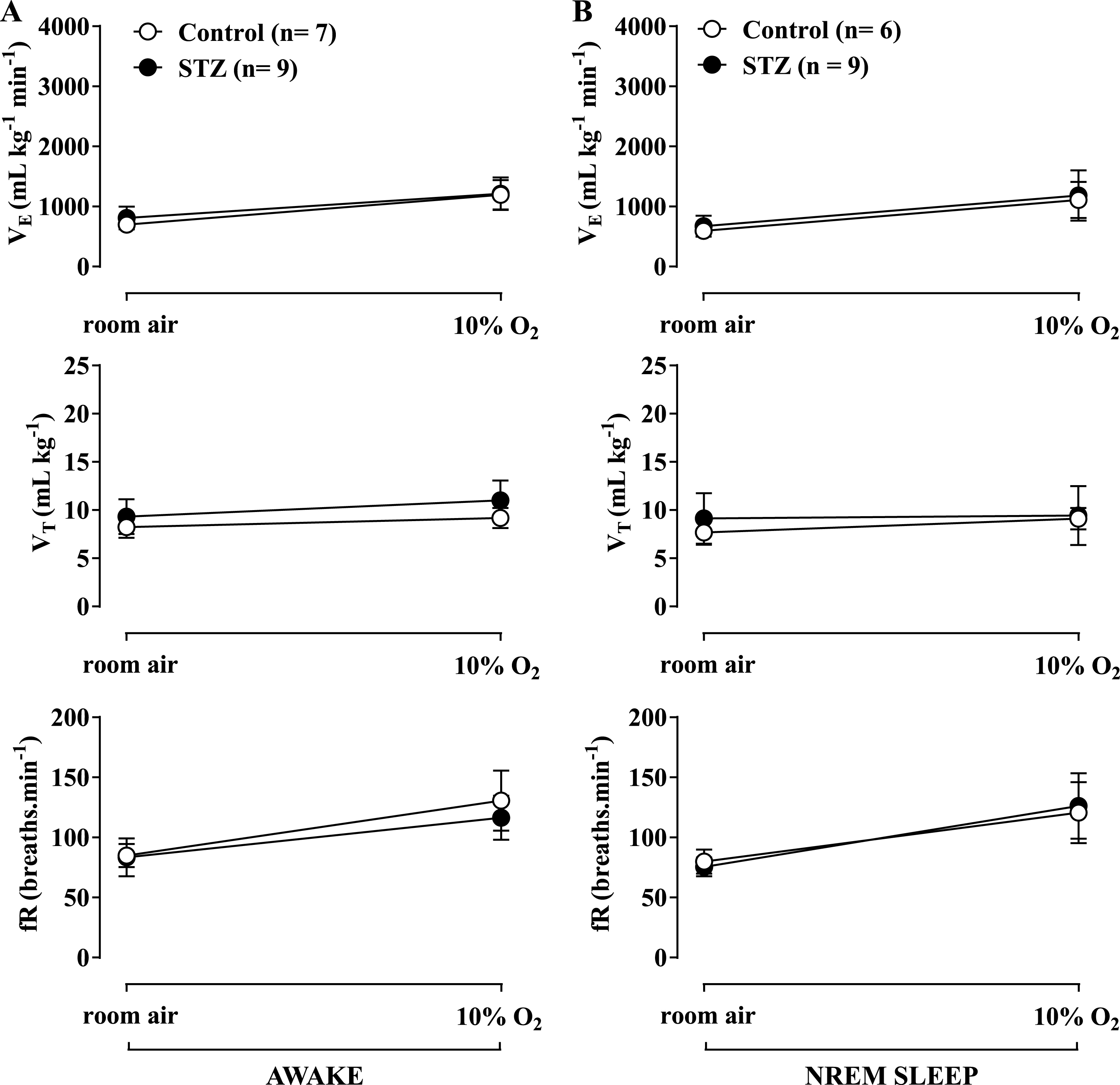
Effect of icv STZ (2 mg/kg) on ventilation (
During hypercapnia (Fig. 2A), the increase in
Hypoxia caused a similar increase in
Respiration during NREM sleep
Figures 3B and 4B show the effects of STZ treatment on ventilatory parameters under room air conditions in rats during the sleep cycle. No difference was observed between the control and STZ groups. Both hypercapnia (Fig. 2B) and hypoxia (Fig. 3B) promoted a similar increase in ventilation in both groups, with no difference between them.
Body temperature (Tb)
No significant difference was observed between treatments during room air conditions and hypercapnia (Fig. 4). Hypoxia caused a similar decrease in body Tp of both control and STZ groups (Fig. 4).

Effect of icv STZ (2 mg/kg) on body temperature of rats during room air, hypercapnia and hypoxia. Values are expressed as mean±SEM. # indicates a significant difference from room air and hypercapnia.
Sleep/wakefulness
Under room air condition (Fig. 5A), the rats spent more time awake, but the STZ-treated rats increased the percentage of total wakefulness compared with the control group (control: 73.9±4.0% versus STZ: 87.0±3.0%, p < 0.05; two-way ANOVA). This effect was due to a significant reduction in the length of the episodes of wakefulness (Fig. 5B), while the number of episodes did not change (Fig. 5C). CO2 exposure and hypoxia significantly increased the time the rats were awake (Fig. 5A), with no difference between treatments during hypoxia. However, under hypercapnic conditions, STZ treatment enhanced the time that the animals spent awake (control: 81.0±5.7% versus STZ: 93.5±2.3%, p < 0.05; two-way ANOVA) due to an increase in episode duration (Fig. 7B) (control: 502.4±179.6 versus STZ: 2096.8±537.9 s, p < 0.05; two-way ANOVA) and a reduction in the number of episodes (Fig. 5C) (control: 5.2±1.3 versus STZ: 2.5±0.6, p < 0.05; two-way ANOVA).
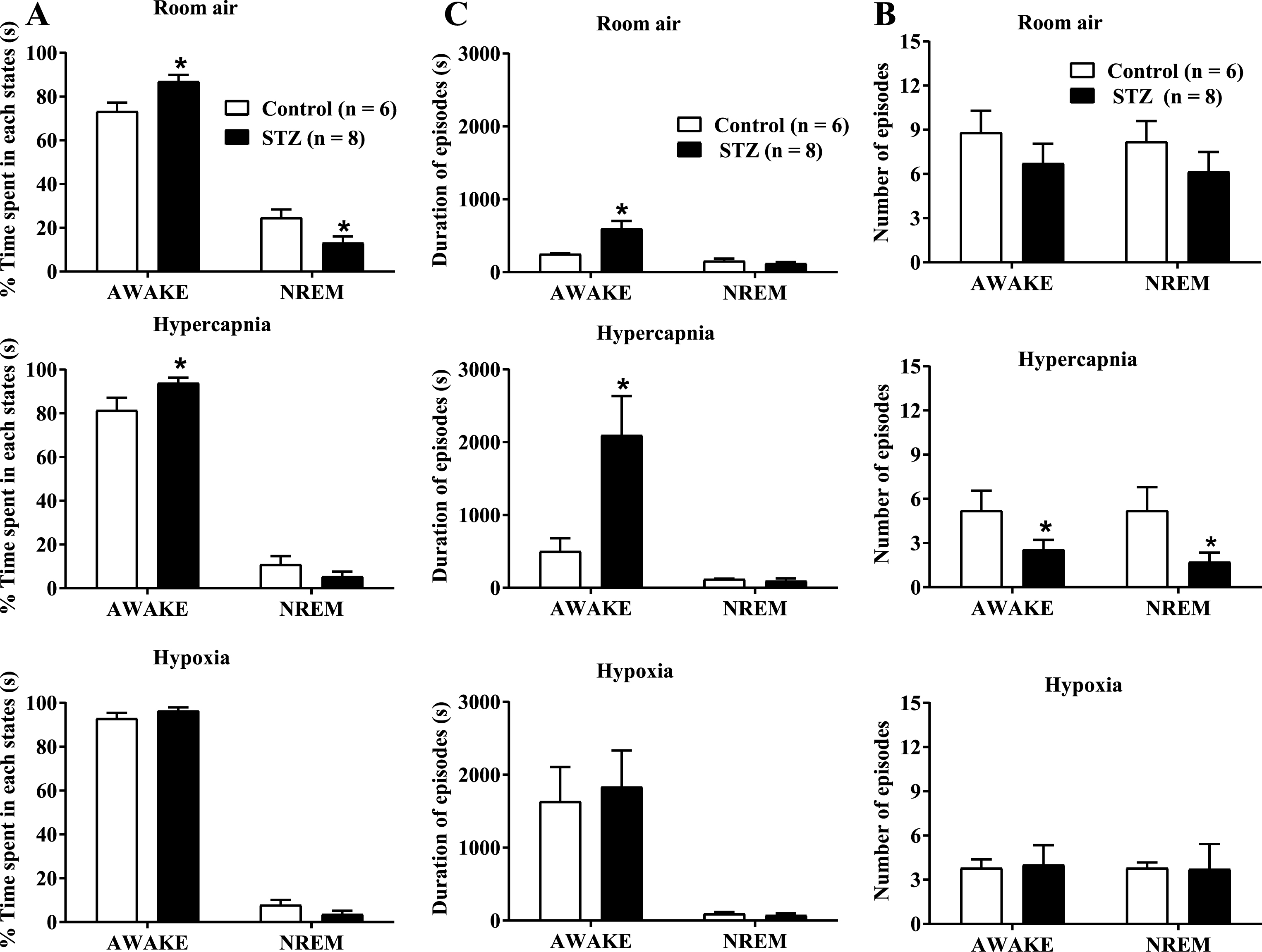
Effect of icv STZ (2 mg/kg) on the percentage (%) of time spent in each state (A), the duration of episodes in seconds (B) and the number of episodes (C) in control and STZ groups during room air, hypercapnia and hypoxia. Values are expressed as mean±SEM. *indicates a significant difference between the control and STZ groups.
Neurochemical analysis
Tau protein phosphorylation
In the analysis of tau protein phosphorylation at the Ser199/202 and Ser396 sites, we evaluated the ratio of phosphorylated tau protein (pTau) levels to total tau protein levels in the regions of interest: LC, RTN, Raphe, PreBötz/BötzC, and HPC.
STZ injection did not alter the levels of phosphorylation of tau protein at Ser199/202 site on LC, RTN, Raphe, PreBötz/BötzC, and HPC (Fig. 6). In addition, STZ injection did not alter the levels of phosphorylation of tau protein at the Ser396 site in the same regions (Fig. 6).

Effect of icv STZ (2 mg/kg) on the phosphorylation levels of tau protein (Ser199/202) (A) and the phosphorylation levels of tau protein (Ser396) (B). The graphs represent the mean radius of the phosphorylated tau protein (pTau) relative to total tau protein levels. The values are expressed as percentage values in relation to the control group (100%). Values are expressed as mean±SEM.
Aβ protein
Expression analysis of Aβ protein demonstrated a 73% increase in this protein in the LC of the STZ group compared to the control (p = 0.0325, Student’s t-test) (Fig. 7). The other structures did not present significant differences between the treatments.
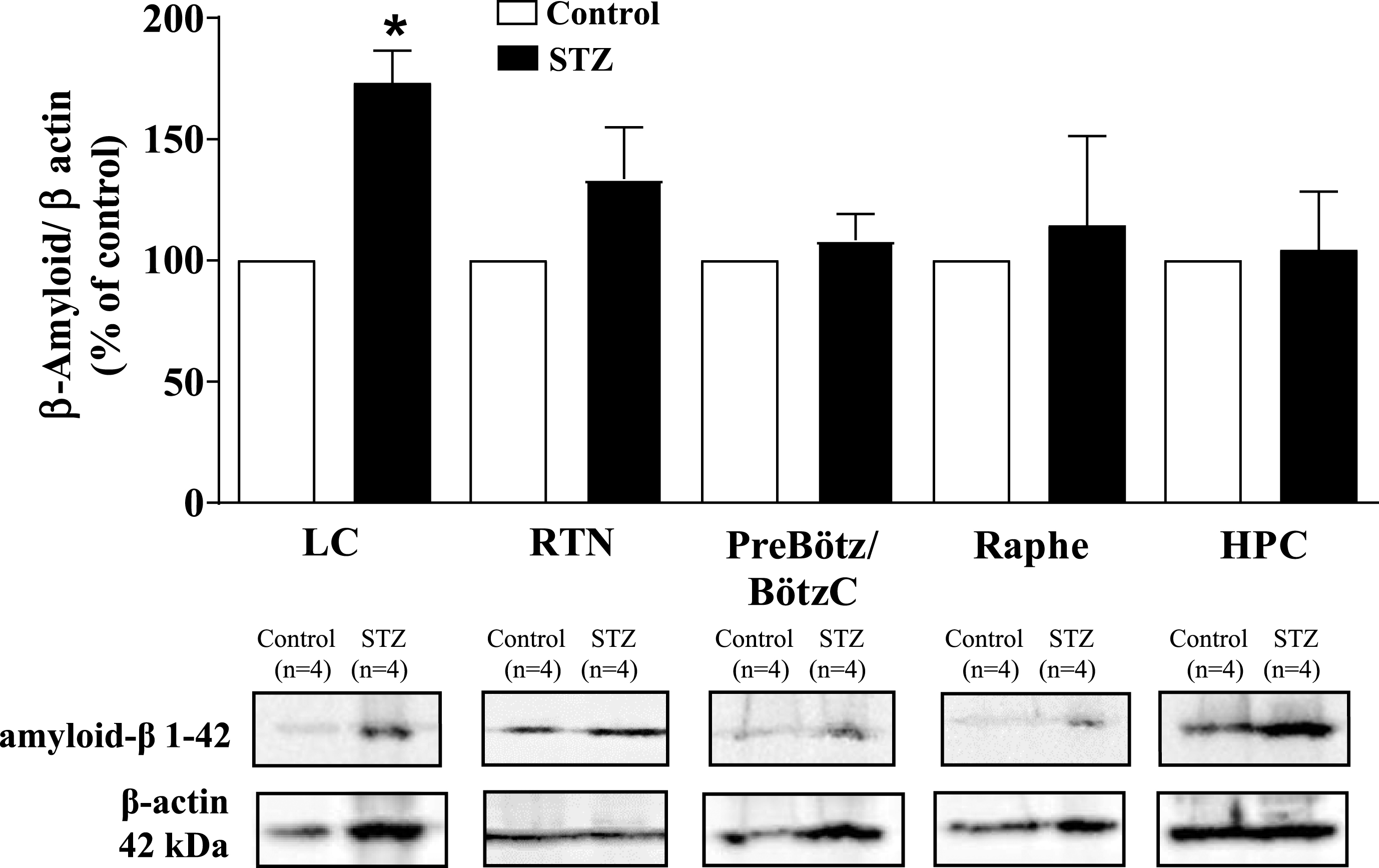
Effect of icv STZ (2 mg/kg) on the expression of Aβ protein. The values are expressed as percentage values in relation to the control group (100%). Values are expressed as mean±SEM. * indicates a significant difference between the control and STZ groups.
DISCUSSION
In the present study we successfully demonstrated that STZ-treated rats had learning and memory deficits and sleep disturbances, with increase time spent in an awake state. Our main results were that STZ rats showed increased Aβ peptide in the LC region and increased ventilatory response to hypercapnia in wakefulness but not during sleep, with no changes in ventilation during room air conditions and hypoxia. Additionally, no changes were observed in body Tp.
In our study, we were able to reproduce the STZ-induced memory deficits. We showed an impairment in learning and retention of spatial memory (as assessed by a higher latency to find the escape box in the acquisition phase of the Barnes maze), as well as difficulty in maintaining spatial reference information (as assessed by the lower number of correct pokes into the target hole of the Barnes maze on the test day). Thus, our results confirm previous studies that have shown that STZ injection causes learning and memory deficits [17, 45]. These findings corroborate the use of STZ in neurodegeneration-induced dementia of the Alzheimer-type in rats, since animals present one of the main cognitive characteristics found in patients with AD [1, 46]. Indeed, evidence that AD patients have problems with spatial orientation—one of the earliest cognitive symptoms—as well as progressive decline in memory, is reported in a number of different studies [47–49].
AD is also characterized by the deposit of extracellular amyloid plaques, composed of Aβ peptide, and intracellular NFTs of hyperphosphorylated tau protein (P-Tau) [3]. In the present study, we did not observe increased phosphorylation of tau protein (Ser396 and Ser199/202) in the LC, RTN, Raphe, PreBötz/BötzC, and HPC. The absence of increased P-Tau after STZ was also reported previously as P-Tau at Ser 199/202 in both rats [30] and mice [17] were unaltered in the hippocampus after 30 and 21 days post-STZ, respectively. On the other hand, increases in P-Tau at Ser396 and Ser199/202 in the hippocampus have been reported after STZ using higher doses (3 mg/kg) than the present study [50] and/or later evaluation [14, 51]. Besides, Grünblatt et al. [31], using lower doses of STZ (1 mg/kg), observed changes in P-Tau protein in the hippocampus, but did not perform a ratio analysis of phosphorylated/total tau; therefore, this may not be considered hyperphosphorylation. In fact, some studies have shown that the onset of NFTs is chronological and hierarchical, since it arises primarily in some regions and late in others [4, 52]. In this sense, it is known that some phosphorylation sites of tau protein hyperphosphorylate at early stages, and other sites in late stages of AD [53, 54], which would also explain the discrepancy in the literature regarding the effects of STZ in P-Tau.
The Aβ peptide in the brain of AD patient was initially considered to be a primary cause of AD dysfunction; however, later studies suggest that the presence of Aβ would be a consequence of early AD events, rather than the cause itself [55, 56]. Similar to what occurs with tau protein phosphorylation, the prevalence of Aβ results from the different stages of AD and different brain regions [57]. In the present study, we observed a 73% increase of Aβ peptide expression in the LC after 30 days of icv administration of STZ. Our data corroborate previous studies showing that central injection of STZ promotes increases in the expression of Aβ peptide [14, 30]. In addition, it is well described that the LC region is affected in AD [58–61]. In this context, we did not observe changes in Aβ peptide expression in the RTN region, Raphe, PreBötz/BötzC, and HPC, which reinforces that the histopathological emergence of AD occurs in a hierarchical and chronological manner [57, 62]. Despite the fact that we did not find histopathological molecular evidence in the present temporal window (30 days after the icv STZ) in the HPC, we observed a cognitive deficit in the model. Therefore, it is possible that other cognitive regions in this temporal window may be affected.
Regarding breathing, STZ treatment did not cause alterations in ventilation under room air conditions and hypoxia during sleep and wakefulness. In the literature, there is a paucity of information on the study of the respiratory system in the progression of AD. Menuet et al. [63], working on a model of taupatia, an important clinical aspect found in AD, observed alterations of the respiratory system, such as upper airway dysfunction, changes in respiratory pattern, and compromised ventilatory response to hypercapnia. However, the authors observed respiratory disfunction in late phases of the disease. In this context, Ebel et al. [28], after 14 days of icv injection of STZ (3 mg/kg), observed respiratory dysfunction in normocapnia, attenuation of the peripheral chemoreflex, as well as a small decrease in respiratory frequency during hypercapnia. Our results differ from those of Ebel et al. [28], as we observed no changes in breathing during room air conditions or hypoxia, and a 26% increase of the ventilatory response to hypercapnia after 30 days of the icv STZ injection (2 mg/kg) only during wakefulness. We believe that these differences may reside in the fact that the authors performed the experiments using a higher dose (3 mg/kg) and a different time window (14 days after the icv injection), whereas in our study, we performed the experiments after 2 mg/kg at 30 days after STZ injection. According to Grieb [64], the neurochemical changes triggered by icv STZ injection(s) follow a time-dependent pattern. Therefore, the effects observed by Ebel et al. [28] may be due to impairment of other brain structures, since they observed a reduction in hypoxic ventilatory response, whereas we showed an exacerbation of the CO2-drive to breathe. Moreover, higher doses may induce more severe and acute neurodegenerative lesions, which could be associated with inflammation and local oxidative stress, whereas lower doses may lead to less severe but more chronic and widespread effects [64]. In this regard, Kraska et al. [65] stated that doses of 3 mg/kg and higher could be considered a model of very aggressive neurotoxic lesions, rather than subtle alterations due to small mechanistic alterations, as would be expected during the slowly evolving dementia processes or during aging. According to the authors, an intermediate dose, e.g., 2 mg/kg, is more relevant as a sporadic AD model.
The exacerbated CO2 ventilatory response during wakefulness observed in the present study may result from changes in the central CO2 chemosensitive areas. In fact, we observed an increase in Aβ peptide expression in the LC region, an important chemosensitive nucleus of the central nervous system [39, 66–68]. We suggest that increased peptide expression may be deregulating the Ca2 + channels, increasing their influx within the neuron, which in turn would increase the release of neurotransmitters, such as noradrenaline (NE) [69]. Increased NOR in the medullary respiratory neurons would trigger a more pronounced ventilatory response to CO2. Previous studies have shown that LC/subcoeruleus neurons represent an early starting point for AD pathology, even preceding the occurrence of cortical lesions [70–72]. Measurements of 3-methoxy-4-hydroxyphenylglycol (MHPG), the principal metabolite of noradrenaline (NE), was found to be increased in postmortem brain tissue of AD patients [73–76]. The authors also found an enhanced ratio of MHPG/NE, which indicates that NE metabolism is augmented in these patients, suggesting an increased activity of the remaining LC neurons to compensate for decreased cerebral NE levels in AD [73–75, 77]. As suggested previously [78], enhanced LC activity may occur in the first stages of AD, prior to cell loss or significant reduction in NE neurotransmission, and this would be harmful due to the enhanced spread of P-Tau, whereas at late stages of AD, LC would be degenerating, and NE levels would be low, and tau pathology is already abundant in the forebrain. Therefore, the activity of LC neurons in AD may be dependent on the disease stage.
Around 45% of patients with AD are affected by sleep disturbances [19, 22] and rodent models of AD also show greater sleep fragmentation and shorter amounts of NREM and rapid eye movement sleep [79, 80]. Our data corroborate these previous studies, since STZ-treated rats spent more time in wakefulness than in NREM sleep during the experimental period in normocapnia and hypercapnia. During hypoxia, both groups increased their wakefulness similarly. In fact, high levels of Aβ correlate with sleep deficiencies [81]. In this context, immunization with Aβ prevents amyloid plaque formation in transgenic mice that develop Aβ aggregation and normalizes sleep/wake patterns [79]. Since we found an increase in Aβ in the LC, and since this nucleus is necessary for maintaining normal durations of wakefulness and to promote immediate sleep-to-wake transitions [82], it is likely that the increase in wakefulness in the STZ-treated rats is related to this fact.
Regarding Tb, the main risk factor for sporadic AD is age [83]. In fact, aging alone is associated with a decrease in Tb, a consequence of a deficit in thermoregulation and, in particular, in thermogenesis [84, 85]. However, studies have shown that patients with AD exhibit increased Tb [83, 86]. Interestingly, rats with icv infusion of Aβ select a higher ambient temperature and show a lower heat tolerance compared to control animals [87]. Increased Tb was also shown in a 6 to 10-month-old 3xTg-AD mouse—a transgenic AD animal model [88]. Recently, the thermoregulatory profile of rats injected with 2 mg/kg of STZ was evaluated, and it was shown that STZ-treated rats presented a higher Tb when compared to controls from day 6 to 25 post-STZ injection [89]. Although at later stages, (>30 days post-injection) STZ-treated animals present a cold-avoidance response, and their basal Tb is no longer different from controls [89]. In accordance, in the present report, at 30 days post STZ treatment no differences in Tb of animals treated with STZ under resting conditions were observed. Furthermore, treated animals seem to keep intact the thermal response to hypoxia, which is considered a regulated fall in temperature due to a drop in O2 consumption during hypoxia [90–92]. Therefore, we suggest that, in the temporal window analyzed in the present study, STZ treatment does not impair thermoregulation in rats.
Conclusion
Our study provides evidence that icv STZ-treated rats present an increased sensitivity to CO2 during wakefulness, but have no changes in basal ventilation or in the hypoxic chemoreflex. These changes in the CO2-drive to breathe might be associated with enhanced Aβ in the LC, since this nucleus is highly involved in hypercapnic ventilatory response, mainly during the wake period [40]. Furthermore, STZ animals showed an increase in the percentage of total wakefulness, which correlates with sleep disturbs observed in AD. Therefore, a better comprehension of respiratory alterations in different phases of the sleep/wake cycle, and the role of LC neurons in this modulation in AD models, is needed in order to target novel approaches for the treatment of this disorder.
Footnotes
ACKNOWLEDGMENTS
This work was supported by Fundação de Amparo á Pesquisa do Estado de São Paulo (FAPESP – 2016/24577-3, 2015/02991-0 and 2015/23426-9) and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq - 442560/2014-1 and 449102/2014-9). The authors thank Euclides Seccato for his technical assistance.