
Abstract
This article aims to reevaluate our approach to female vulnerability to Alzheimer’s disease (AD) and put forth a new hypothesis considering how sex differences in the locus coeruleus-noradrenaline (LC-NA) structure and function could account for why females are more likely to develop AD. We specifically focus our attention on locus coeruleus (LC) morphology, the paucity of estrogens, neuroinflammation, blood-brain barrier permeability, apolipoprotein ɛ4 polymorphism (APOE ɛ4), and cognitive reserve. The role of the LC-NA system and sex differences are two of the most rapidly emerging topics in AD research. Current literature either investigates the LC due to it being one of the first brain areas to develop AD pathology or acknowledges the neuroprotective effects of estrogens and how the loss of these female hormones have the capacity to contribute to the sex differences seen in AD; however, existing research has neglected to concurrently examine these two rationales and therefore leaving our hypothesis undetermined. Collectively, this article should assist in alleviating current challenges surrounding female AD by providing thought-provoking connections into the interrelationship between the disruption of the female LC-NA system, the decline of estrogens, and AD vulnerability. It is therefore likely that treatment for this heterogeneous disease may need to be distinctly developed for females and males separately, and may require a precision medicine approach.
Keywords
OBJECTIVE
This article puts forth a new hypothesis that has identified and integrated characteristics in the locus coeruleus-noradrenaline (LC-NA) mechanism in females that distinctly spotlights their precarious nature to disproportionately developing Alzheimer’s disease (AD). Our novel hypothesis endeavoring to explain female AD vulnerability is based upon the differential and detrimental effects of the loss of estrogens and degradation of the LC-NA system in females. The present proposal aims to 1) highlight existing evidence which exhibits how the aforementioned effects may have an impact on female AD susceptibility; 2) bring awareness to the need of an in-depth investigation into the loss of estrogens and diminished production of the LC-NA system to combat AD sex differences; and 3) promote further research in the field in order to advance this hypothesis forward.
BACKGROUND
Evolution of locus coeruleus involvement in AD
The locus coeruleus (LC) (Fig. 1) has become the focus of numerous investigations due to its influential functions throughout the brain (Box 1) and its role as the earliest host of AD pathology [1, 2]. Renowned discoveries by Braak and colleagues have identified intraneuronal abnormal tau in the human LC prior to LC neuronal loss and subsequent LC neuronal death as tau burden increases [3, 4]; this research has since been validated in a more recent rat model [5]. Consequently, the presence and aggregation of extracellular amyloid-β protein (Aβ) in the human LC has been shown to accompany neurofibrillary tangles in the late-stage processes of Braak and Braak (BB), particularly stages V and VI [3, 4]. Moreover, the afferent and efferent projections of human LC neurons are thin, vastly arborized, and poorly myelinated, causing them to be eminently fragile [6]. This is particularly significant regarding the afferent nerve fibers that permeate through the gut to the vagus nerve and central nervous system (CNS) microvasculature; this causes the LC-NA system to be particularly vulnerable to being exposed to harmful metabolites, pathogens, and viruses [7]. These noxious microorganisms can then intensify tau hyperphosphorylation, each of which can be transmitted throughout the brain, not only degrading the LC-NA pathway but also causing detrimental alterations of more extensive neuronal function. Surviving LC neurons containing abnormal pathologies spread trans-synaptically to other brain regions exacerbating synaptic dysfunction, resulting in cognitive decline and memory impairment, and neurodegeneration [7–10]. These pathological changes can also depress neurovascular coupling and increase inflammation and blood-brain barrier (BBB) permeability [2, 12]. The association between severe neuronal loss in the LC and AD is strongest in the rostral and medial-dorsal projecting neurons [13–17], which has a major impact on disease outcome given that the loss of LC neurons reduces the availability of NA in the forebrain, hippocampus, and other target-regions; degree of LC cell loss has been shown to be highly correlated with global cognitive score, neuropathological accumulation, duration and severity of illness in elderly patients [16, 19]. Consequently, the disruption of the LC-NA pathway may very well be at the epicenter of the development of AD [1, 20] (Fig. 2).

Location of the locus coeruleus. The LC (circled) is located on the lateral floor of the fourth ventricle and upper dorsolateral pons.
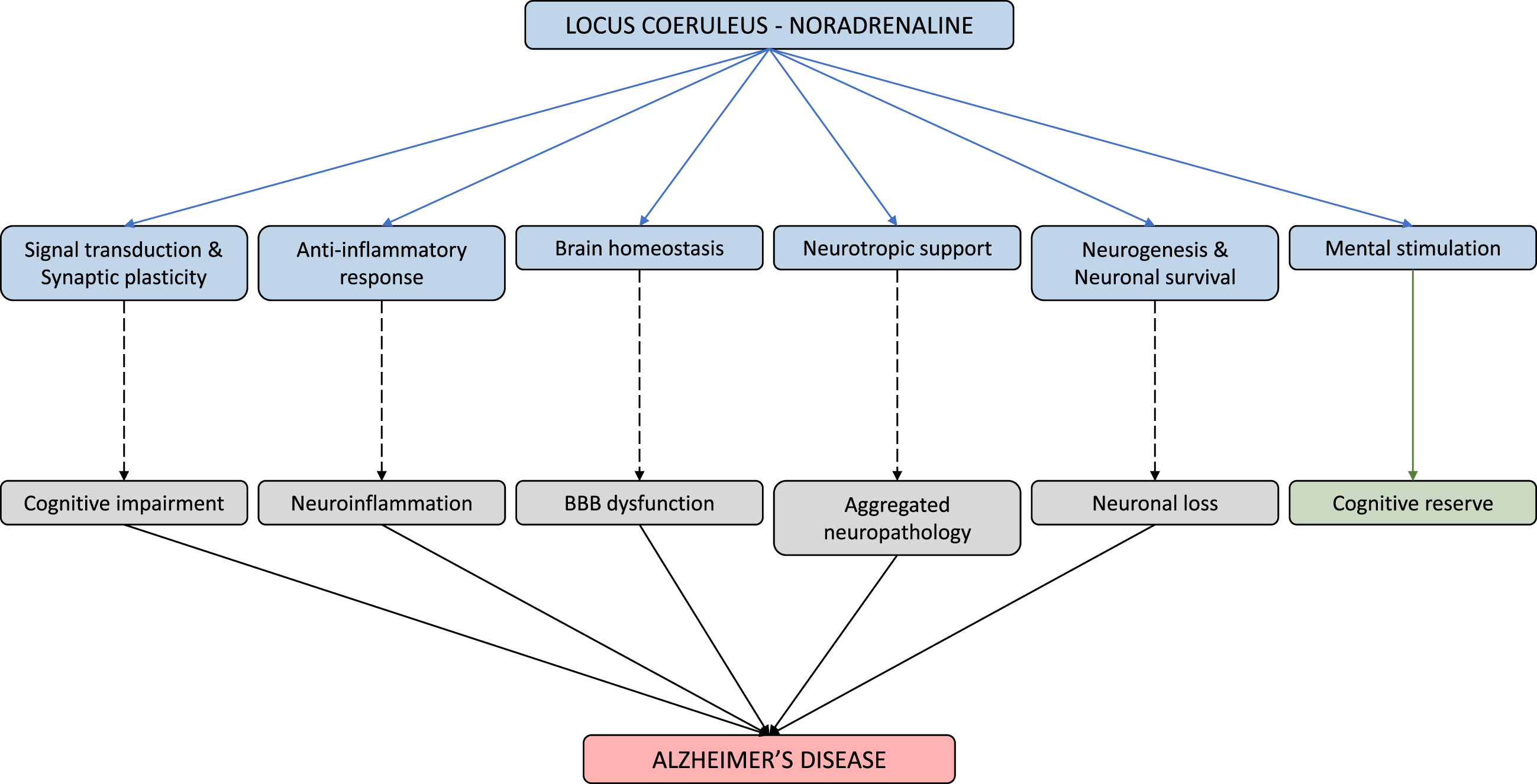
Physiological and pathophysiological implications of the LC-NA system. The LC-NA system is involved in a host of physiological functions (shown via blue line). However, as the LC starts to falter, these mechanisms begin to weaken resulting in various AD symptomology (shown via dashed line). In reverse, mental stimulation of the LC, provided by anything novel and salient, leads to improved cognitive reserve (shown via green line).
Evolution of sex as a risk factor for AD
Studies examining patients have highlighted the disproportionate AD vulnerability of females over males, a ratio of approximately two to one [21], has brought forth valuable discoveries of sex differences in areas such as onset and conversion [22], rate of cognitive decline [23, 24], and brain atrophy [25]. Significant findings from these studies emphasizes the substantial consequence that disregarding sex may have for the vast range of current studies that do not sufficiently consider sex as a moderating variable in experimental, clinical, and therapeutic studies, and in identifying potential gender-specific prevention and treatment approaches to AD. Ongoing research to explain higher female incidence AD rate is primarily interested in risk factor profiles and prevention of AD, specifically heeding the effects of the apolipoprotein ɛ4 allele (APOE ɛ4) variant and the diminution of ovarian hormones; however, current studies have yet to determine or justify why the ratio exists.
RATIONALE
Today, there are a total of 50 million dementia patients across the globe, including 6.2 million AD patients that reside in the United States (U.S.) alone [26]. Among the patients in the U.S., 3.8 million are female and 2.4 million are male, a matter of substantial interest regarding our theory and AD research as a whole. Considering the significant number of AD patients in the U.S., which is projected to double by 2050, and the two to one ratio it yields, this hypothesis has the potential to help in early identification and intervention of AD for 3.8 million females currently, and 8.3 million females by 2050 [26]. However, in order to do so, it is pivotal that we reassess the approach that is taken for treating this heterogenous disease, particularly given the fact that no effective treatment exists. A substantial amount of evidence acknowledges the role of the LC in AD pathogenesis and progression. Rapidly becoming one of the most emerging issues in AD research, sex differences and why females disproportionately develop AD has recently received greater attention. However, to our knowledge, there are no existing investigations examining an association between sex differences and the LC in AD diagnosis, thus highlighting a major gap in the current understanding of risk and prevention of AD.
NEW HYPOTHESIS
Upon reexamination of disseminated animal and human research alike, we hypothesize the escalated vulnerability of female AD is caused by hormonal changes effecting the LC-NA system that first results in a loss of estrogens and noradrenaline (NA), and secondly, a consequential cascade of neuroinflammation, loss of BBB integrity, and neuropathological changes contributing to the onset and progression of AD.
For consideration of estrogens research
Female and male sex hormones include progesterones, androgens, and estrogens; however, the levels in which these hormones are produced differ depending on their sex, whereby androgens are more relevant for males, estrogens are more critical for females. Although all aforementioned sex hormones have their role to play, estrogens are of primary concern regarding females, considerably more so when females begin approaching the age range of perimenopause, menopause, and postmenopause. In females’ reproductive years, estrogens are predominantly produced by the ovaries and adrenal glands. Although estrogens are primarily known for their capabilities regarding female reproduction, these estrogens also circulate through the BBB in order to reach the brain and become a “master regulator” of biological functions as well as aid in neuroprotective effects [27]. The classification of estrogens contains three forms of the sex hormone: estrone (E1), estradiol (17β-estradiol or E2), and estriol (E3). Of these three estrogens, 17β-estradiol is the most prominent and highly researched form of estrogens. In addition to circulating estrogens produced by the females’ reproductive organs, 17β-estradiol can also be produced endogenously in the brain, also known as brain-derived estrogen [27].
In the latter part of a female’s lifetime, females will undergo stages of perimenopause, also known as menopausal transition, menopause, and postmenopause, respectively. Perimenopause typically begins at an average age of 47 and persist for approximately 5–8 years [28] and is denoted by females experiencing irregularity of duration and time between menstrual cycles, reproductive senescence, as well as, a gradual decline in ovarian hormones [29, 30]. Menopause typically occurs among females who fall in the age range of 48 to 52 years of age on average [31], and is defined as the 12-month period of amenorrhea where estrogens (evaluated by 17β-estradiol levels per STRAW+10) begin to rapidly decline [29]. After 12 months of cessation of menses, the occurrence of the final menstrual period can be confirmed, as well as the beginning of postmenopause, a stage in which females cease to menstruate and estrogens (evaluated by 17β-estradiol levels per STRAW+10) continue to decline for approximately 1 year, after which the estrogens begin to stabilize at the lowest levels in a female’s lifespan [29, 30]; it is also of note both brain-derived and circulating 17β-estradiol mirror the same amount of significant decline when postmenopausal females were compared to females who were premenopausal [32].
As previously mentioned, the classification of estrogens contains E1, 17β-estradiol or E2, and E3; however, it has been brought to our attention that the field of neuroscience commonly uses the umbrella term “estrogen” as an all-inclusive term for estrogens. As a consequence of the utilization of this umbrella term, the type of estrogen being referred to in numerous studies consulted for this prospective paper was ambiguous and resulted in our assumption that studies were most likely discussing 17β-estradiol when using “estrogen.” Therefore, it must be mentioned that although we suspect this report is highly representing the effects of 17β-estradiol, the potential contributions and effects of other hormones, such as, E1, E3, and progesterone, should not be neglected and must be considered when pondering females and AD. Furthermore, this limitation accentuates an area of research that neuroscience must improve upon. Moving forward, it is critical for researchers to 1) be more specific and provide more thorough descriptions when discussing sex hormones including estrogens, as well as 2) conveying whether they are referring to circulating or brain-derived estrogens in order to prevent obscurity of information, and 3) identify if females fall within reproduction, perimenopause, menopause, or postmenopause by using an outlined set of criterion, such as the STRAW+10 criteria [29]. This issue should be addressed promptly considering the upward trend in research regarding sex differences in AD. Finally, it must be noted that the specific form of estrogen will be documented in this review if it was identified; however, due to the forgone impediment, the majority of the evidence provided will use the term “estrogens.”
Estrogens influence on LC-NA regulation
By way of various regulatory mechanisms, gonadal hormones, such as estrogens, have significant effects on human brain function, including cognition, bioenergetic systems, and homeostasis [27, 33–35]. Estrogens’ direct ability to modulate LC activity is promoted by the presence of both nuclear estrogen receptors (ER) ER-α and ER-β and cell-membrane ER subtype G protein coupled estrogen receptor 1 (GPR30) found in wide distribution throughout the LC [36]. Additionally, studies looking at various male and female rat species have depicted that estrogens plays a significant role in facilitating sex differences in LC-NA morphology, from embryonic development into adolescence [37, 38] (for more details, see section Selective vulnerability and multiple clinical phenotype). Likewise, studies using ovariectomized female rats have exhibited that estrogens have a considerable impact on LC function by acting as a presynaptic modulator of NA release by altering the level of enzymes synthesizing NA, such as tyrosine hydroxylase and dopamine β-hydroxylase [39, 40]. In addition, estrogens are a key regulator in the degradation of NA by reducing catechol-O-methyltransferase levels, an essential enzyme needed for the breakdown of NA [41]. Given the evidence regarding estrogens’ effect on humans, and if the findings from the aforementioned rat studies are translatable to humans, this suggests the females could have an increased capacity for NA production and release.
Considering estrogen’s strong influence on LC-NA regulation [42, 43], and menopause causing a rapid loss of the ovarian sex-hormones in all females’ midlife, we hypothesize the loss of estrogens may explain the decline of the female LC-NA system, resulting in an increased risk of developing AD. This perspective piece highlights three key determinants of AD in which estrogens and the LC-NA make significant contributions that require further examination: namely neuroinflammation, BBB, and APOE ɛ4.
Effects of LC and estrogens activity on neuroinflammation
Mounting evidence implicates glial-mediated inflammation as a major contributor to cognitive decline and neurodegenerative processes [44]. When reacting to neuroinflammation, microglia cluster around amyloid deposits and can become chronically activated [45–47]. Overactive glia cells are unable to effectively clear Aβ. Aβ accumulation upregulates pro-inflammatory release and exacerbates the inflammatory cascade leading to synapse elimination, further amplifying neurodegeneration, and cognitive function declining [48–50], along with increased neurotoxicity and neuroinflammation wherefore AD neurodegenerative processes are amplified [51–55]. The above immunological cascade of events is gaining traction as having a fundamental role in the AD neurodegenerative processes [56–58].
Current evidence in animal studies favors the idea of an early fundamental link between neuroinflammation and LC degeneration that brings about AD pathogenesis [55]. These studies and similar findings in humans have demonstrated that NA acts as an endogenous CNS immunomodulator by helping maintain homeostasis [52, 60], influences on other neurons and glia to exert potent neuroprotective effects as an anti-inflammatory, and operates as a suppressor on pro-inflammatory gene induction [12, 62]. As AD develops, as seen in BB stages V, there is a progressive neuronal degeneration and cell loss in the LC thereby decreasing NA availability [4]. This decrease may carry negative implications given that in vivo amyloid precursor protein mice models suggest deterioration of LC varicosities prevents the release of extra-synaptic NA to surrounding glial cells, and consequently inhibits phagocytosis and interferes with the amyloid pathology generating cycle [63], thus promotes chronic inflammation and the formation of Aβ in areas usually innervated by the LC [53].
Estrogens, such as 17β-estradiol, exert influential modulatory effects on microglia and are shown to directly attenuate pro-inflammatory release from these cells, as well as, indirectly prevent the transcription of pro-inflammatory cytokines [64]. As females undergo menopause, the decline in 17β-estradiol synthesis and changes in the immune system significantly increase the levels of pro-inflammatory markers brought on by the loss of anti-inflammatory and pro-resolution activity, thus suggesting a pro-inflammatory phenotype in females [64]. Non-infectious diseases such as obesity, diabetes, and depressive illnesses are all associated with the same pro-inflammatory phenotype [65]. This is not coincidental, as those diseases can also be classified as female driven risk factors that increase the likelihood of developing a comorbid diagnosis of AD [65].
Taken together, we can infer that postmenopausal females are more likely exposed to additional inflammation due to higher levels of Aβ and immuno-stimulated glia. A noteworthy study detected the presence of Aβ pathology in 1,031 postmortem human brains when using immunocytochemistry (4G8) and Campbell-Switzer staining [4]. Of these cases, 4% exhibited Aβ deposition in the fourth decade of life; however, the most prominent levels of Aβ plaques were observed in the sixth decade and beyond. More indicative to our proposal, a significant correlation between Aβ deposition and age was found to progress more rapidly in females [4]. Without estrogens, notably 17β-estradiol, and LC-NA neuroprotection, the ability to downregulate the transcription of inflammatory genes in microglia and reduce amyloid neuronal toxicity is decreased [66–68]. Consequently, substantial depletion of NA increases the magnitude of neuroinflammation and gives rise to a relentless inflammatory mechanism that accelerates the neuropathology it was created to eradicate, leading to more rapid development of AD pathology in females [63, 70].
Effects of LC and estrogens activity on BBB
Studies using various species of rats have shown the LC-NA to have a role in BBB maintenance, including the regulation of BBB permeability and neurovascular coupling [71–73], and has also been shown to modulate morphological and functional properties of the BBB [71, 75]. More particularly, a morphometric analysis indicated intracortical astrocytes being one of the direct targets of LC-NA terminals thus implying that NA has a regulatory influence on astrocytic functions of BBB permeability and homeostasis [74, 77]. Furthermore, NA, by means of an association with cyclic adenosine monophosphate, regulates the formation and stability of tight junctions (TJs) [78–81]. This can be seen in the way TJ disarrangement and gliosis is caused by noradrenergic fiber degeneration [75].
In addition, the emergence of neuroinflammation and neurodegeneration in the LC, due to diverse pathogens, are two physio-pathological aspects promoting BBB dysfunction [11, 82]. As the BBB begins to lose functionality, a decrease in barrier tightness is observed due to the degradation of TJs [83]. This maladaptive increase in permeability allows for the transport of cytokines and immune cells into the CNS and cerebrospinal fluid [84]. As a result, a secondary inflammatory response creates an environment suited for a pro-inflammatory take-over that compromises Aβ clearance, affects TJ integrity, causes endothelial cells (EC) damage, and insufficient nutrient supply; comparable to what is identified in AD [85, 86]. In addition, defective neurovascular unit cells that prompt neurovascular impairment can result in neurodegeneration and the deterioration of cognitive abilities and advance the inception and progression of AD [83, 87–94].
Animal studies using endogenous estrogen have shown 17β-estradiol’s ability to protect the vasculature of intercellular junctions via targeting and regulating claudin-5, which is specifically needed to enhance permeability [95], and annexin A1 protein [96], an anti-inflammatory regulator of TJ formation, thus, governing BBB integrity [97, 98]. Moreover, 17β-estradiol has the capability to act as a homeostatic signal by limiting the disturbance of lymphocytes and enhancing TJ function and has a profound impact on the BBB, thus marked differences may occur as females age and the associated circulating 17β-estradiol declines [99].
Another important target of both NA and estrogens is the cerebral microvasculature [100–102]. LC-NA deafferentation impairs forebrain cerebrovascular function during the preclinical and prodromal stages of AD [18]. NA’s ability to regulate vascular related functions, such as cerebral blood flow, can be seen in anatomical evidence of rat studies depicting LC ascending fibers delivering direct innervation to the microvascular endothelium [74, 102]. Additional rat studies have observed estrogens’ exposure guards against apoptosis of ECs, alters mitochondrial function, increases cerebral blood flow, as well as, induces anti-inflammatory actions on cerebral blood vessels, and in turn suppresses endothelial inflammation [100, 103]. Due to the relatively high energy demands of these specialized ECs, cerebral vascular endothelium contains more mitochondria than endothelium in any other vascular beds, thus increasing energy production needed to maintain the BBB efficiency while also reducing oxidative stress that is associated with AD [104, 105]. Collectively, the presented animal literature exhibits how the LC-NA and estrogens play a critical role in protecting and regulating the BBB, as well as, how the degradation of the BBB results in a significant decline in cognitive abilities and an increase in neurodegeneration. Taking this evidence into account advances the implication that a combined loss of estrogens and NA leads to females becoming more susceptible to consequences brought on by having a more permeable and less governed barrier. Nevertheless, considering the fact that present data is predominantly based on animal models, replicating the findings of these studies in humans will be advantageous to our current hypothesis.
APOE ɛ4 interaction with LC and estrogens
To date, it is suggested that the strongest genetic risk factor for AD is the presence of the APOE ɛ4 allele [106]. Mice studies have linked APOE to BBB dysfunction via ɛ4 alleles’ conduciveness for vascular atrophy resulting in increasing permeability and reducing cerebral blood flow [107, 108]. In addition, carriers of the ɛ4 allele have been shown to have altered astrocyte bioenergetics that restrict astrocytes from allowing proper lipid and glucose transport, storage, and utilization [109]. APOE ɛ4 carriers display a disease-associated microglia profile, including increased pro-inflammatory cytokine production with impaired phagocytosis, deficient debris clearance, and impaired migration capability. These detrimental functional effects predispose carriers to neuropathological features associated with AD, such as impairments in cerebral glucose metabolism, neuronal signaling, neuroinflammation, and mitochondrial function [110, 111].
APOE ɛ4 status is also instrumental in the mediation of neuronal maintenance, growth, and synaptic repair. Synaptic disturbances in the LC-NA tract in APOE-deficient mice were more evident the farther away the axon nerve terminal is from the cell body [112]. These findings enhance the likelihood that the LC-NA projecting pathways composed of longer axons and branch formations could potentially be dependent on APOE for normal function. A supplementary analysis in patients that assessed the role of APOE ɛ4 on the compensatory mechanism neurons undergo when in distress resulted in LC neurons experiencing insufficient dendritic remodeling in addition to neuronal degeneration[113]. Taken together, this illustrates that LC neurons are likely compromised in APOE ɛ4 carriers and this could contribute to the breakdown of the LC-NA pathway more rapidly than in non-carriers, and potentially results in an earlier age at onset and a more expeditious development of the disease [112, 113]. In addition, a recent female-only rodent study indicated APOE ɛ4 inhibition of vesicular monoamine transporter 2 (VMAT2) in LC exacerbates tau pathology in AD, signifying that the LC’s initial role in contributing to AD may conceivably be due to APOE ɛ4’s pathological effect [114].
APOE ɛ4 also impacts females independent of AD diagnosis. Studies using non-cognitively impaired ɛ4 allele carriers reported an APOE ɛ4-by-sex interactions on human brain metabolism, structure, and functional connectivity [115–117]. This indicates an unestablished mechanism of action between sex and APOE ɛ4 [116]. Ultimately, APOE ɛ4 has a multifaceted effect on cell types through diverse pathways, consequently disrupting the homeostasis and leading to greater neurotoxicity [109]. However, knowing that females’ more complex dendritic structure leaves them susceptible to harsher synaptic disturbances, combined with APOE ɛ4 influencing dendritic remodeling, we speculate that the extent of synaptic degeneration of the LC-NA pathways in AD are more pronounced in female ɛ4 allele carriers. Furthermore, females have an increased risk in mild cognitive impairment and AD occurrence around menopausal age ranges [31], we believe it can potentially be explained by a loss of estrogens; this hypothesized interaction with APOE ɛ4 has yet to be investigated.
Future experiments and validation studies
Hormone replacement therapy (HRT) studies are of great popularity given the link between estrogens depletion and the increased risk of AD; although previous trials are not without debate and limitations [118], their contributions regarding HRT and female AD are invaluable assets that have facilitated ideas for future research. The past two decades have provided us with evidence that has not strongly supported the hormonal loss hypothesis in relation to cognitive function and/or dementia risk [119]. These studies have done well to incorporate various sex hormones in multiple combinations through multiple application methods. The Women’s Health Initiative (WHI) conducted a study on postmenopausal females 65 years and older [120]. Upon examination of the effects of estrogens alone (conjugated equine estrogens (CEE)), estrogens plus progestin (CEE plus medroxyprogesterone (MPA)), Premarin, a CEE that contains estrone and 17²-estradiol, as well as MPA, the WHI concluded that not only are the use of these HRTs not recommended but that the risks (of cancer, cardiovascular, and dementia) outweighed the benefits [121, 122]. A similar study, the Cache County Study (CCS), included postmenopausal females of 65 years of age or older (oophorectomized and non-oophorectomized) who reported uses of either “unopposed” or “opposed” estrogens [123]. The results of CCS indicated that postmenopausal females who began HRT 1–5 years from menopause exhibited a trend toward AD risk reduction, whereas participants who began HRT closer to or after menopause showed no association to lower AD risk [123]. A third study, The Kronos Early Estrogen Prevention Study (KEEPS), examining postmenopausal, healthy, non-hysterectomized females found no cognitive benefits after participants initiated HRT for four years via oral CEE plus micronized progesterone or transdermal estradiol plus micronized progesterone [124]. Additionally, The Early Versus Late Intervention Trial with Estradiol (ELITE) study looked to identify if utilizing oral micronized 17β-estradiol with progesterone gel for five years would have any effect on cognitive abilities in females who were more than 10 years postmenopausal [125]. ELITE concluded with no affects being evident in verbal memory, executive functions, or global cognition [125]. Taken together, the major critique of the previous four studies includes participants being in the postmenopausal stage, thus prompting the notion that future experiments must consider hypotheses such as estrogen administered into a healthy environment, “healthy cell bias” [119], and the efficacy of HRT in relation to the timing and onset of menopause, “window of opportunity”[126]. There has been some evidence supporting the timing hypothesis of HRT in observational studies where females have self-reported menopause status, timing of HRT initiation, and if opposed or unopposed estrogen was utilized [126]. Together, these observational studies provide support for the timing hypothesis given that each study’s results showed reduced risks of AD in groups that began HRT in earlier stages of their life [123, 128]. These findings stand in stark contrast compared to the previous four studies mentioned, particularly a follow-up study to the CCS which identified females who began HRT 1–5 years before menopause had a 30% reduced risk of AD, and a 37% reduced risk when taking HRT for 10 or more years [123].
In order to validate the present hypothesis, it is essential for future studies to persist in identifying sex differences in the LC-NA system. Moreover, it is imperative that studies are translated from rodents to humans so that we may discontinue drawing inferences from these studies and confirm the effects in humans. Additionally, our investigation of previous publications determined that a majority of animal research and experimental treatment trials are largely dominated by work carried out in males. In order for scientific research to excel, experimental and clinical design must undergo considerable modifications such as routinely stratifying data by sex and ensuring studies are statistically powered to detect interaction effects between sexes. Furthermore, a set list of guidelines must be established for reporting sex differences [129], otherwise inconsistencies and uncertainties will remain in our understanding of female AD.
Future experiments may embrace the goal of working towards establishing an interrelated timeline connecting the dysregulation of the LC-NA system, the decline of sex hormones, and AD vulnerability together. This will allow for studies conducted to utilize a more relatable timeline that associates the females’ stages of menopause with the advancement of neuropathology in the LC using the pre-established BB stages. The institution of this timeline will enhance the investigation of the hormonal loss hypothesis when testing how HRT affects the LC-NA system and enable studies to place consistent, corresponding timeframes for their findings regarding “window of opportunity” and “healthy cell bias.” Taken together, these future investigations are instrumental in exploring methods to upregulate estrogens and LC-NA mechanisms in order to deter further neuropathological accumulation and neurodegeneration. It is vital to address this hypothesis promptly seeing that the increase in today’s lifespan could indicate that on average females would spend up to one-third of their life without production of estrogens [130].
CHALLENGES ADDRESSED BY OUR HYPOTHESIS
Aging
The increased longevity of females is one of the most argued justification for the increased risk of AD and has recently come under more scrutiny as of late [131]. Evidence indicates aging alone is not a suitable determinant for LC degeneracy [132]; however, our present theory insinuates that aging is notable given the association of age and onset of menopause in the majority of females. As mentioned previously, the current mean age of the onset of menopause is approximately 51 years of age, with a mean range of 48 to 52, and perimenopause typically begins at an average age of 47. This is significant due to these age ranges of the emergence of perimenopause and menopause, respectively, coinciding with BB stages Ia, Ib, I, and II. At these ages, a vast majority of human autopsy brains were classified in BB stages Ia and Ib whereby prominent AT8 immunoreactive pretangle material was displayed, noted as the source of argyrophilic neurofibrillary lesions, thus facilitating neuronal vulnerability [3]. Additionally, approximately half were classified as BB stages I and II at which point Gallyas-positive neurofibrillary lesions were manifesting in the entorhinal regions, indicative of the potential initiation of neuronal deterioration that could occur in future BB stages [3]. A highly significant correlation between AD-related pathologic findings and age has been found in females, yet no current significant difference has been seen between females and males in early and mid BB stages. However, more significant variations are seen between males and females as individuals increase in age. Given the strong influence estrogens have on LC-NA regulation [42, 43], and the fact that females suffer from greater implications than males the further away they get from perimenopause and menopause age, it must be considered if a decrease in estrogens may have a disproportionately negative effect on the LC-NA system of females, thus resulting in an increased risk of developing AD.
Selective vulnerability and multiple clinical phenotypes
Animal studies have depicted sexual dimorphism in LC morphology [37, 41], with sex differences in LC size, make up, and neuronal number, depending on the strain of rat used. Furthermore, studies using Wistar rats show developing females to have a higher volume and neuronal count [38, 133], and this, together with sex differences in dendritic structure [37], may be due to the influence of estrogens during LC neurogenesis [38, 134]; female dendritic trees have been shown to be longer and more complex, with greater numbers of branch points and higher-order branching [37]. If these findings could be reproduced in human studies, this may support the prospect that females receive higher levels of synaptic input and process additional information coming into the peri-LC, from regions such as the periaqueductal gray and nucleus tractus solitarius (NTS) [135]. The NTS can influence NA activity both directly via synapses on neurons in the LC, and indirectly via connections linking the LC to the amygdala and hippocampus [136]. Alternatively, the NTS is also involved in the regulation of gastrointestinal activity along with the vagus nerve, which, as mentioned previously, could result in exposing the LC to toxic agents [7].
Due to the LC mediating arousal and autonomic function, the potential multifaceted dendritic structure of the female LC and the increase in NA induced by estrogens allows for the potential of heightened emotional arousal response [37]. A repercussion of the increased stimulation seen in the female LC could be the rationale for hyperarousal resulting in and contributing to the increased rates of disorders seen in females [41], such as post-traumatic stress disorder, depression, and sleep disturbance, all of which share clinical phenotypes with, and are comorbid risk factors of AD and coincidentally are more predominant in females [137]. This connection between their modified LC-NA network and the presentations of behavioral manifestations in females emphasizes the need for further examination into the causes-and-effects of selective vulnerability in female AD.
An additional consideration regarding LC vulnerability stems from results of a recent mice study investigating the effects of 3, 4-dihydroxyphenylglycollaldehyde (DOPEGAL) on the LC indicated that monoamine oxidase A (MOA-A) metabolizes NA into DOPEGAL through oxidative stress, which in turn increased asparagine endopeptidase, and resulted in tau N368 cleavage which is susceptible to hyperphosphorylation, aggregation, and propagation, leading to noradrenergic cell death [138]. This study contributes significant information when considering our hypothesis given that estrogens and progesterones are both regulators of oxidative stress [139]. As mentioned previously, females begin to experience perimenopause and menopause, respectively; approximately when BB stages Ia and Ib, I, and II occur in humans, coinciding with the timing when pretangle and tangle material is observed, and neuronal vulnerability is facilitated. This is significant on account of the process of NA oxidizing to DOPEGAL and ultimately resulting in tau cleavage and noradrenergic cell death is conceivably protracted, and thus are not observable until later BB stages, leading us to speculate if the loss of estrogens and progesterones in early BB stages due to perimenopause and menopause is potentially one of the explanations of NA metabolizing to DOPEGAL due to the lack of the female’s ability to regulate oxidative stress.
Furthermore, findings from a recent study indicated similar results where increased levels of NA-metabolism were connected to greater cerebrospinal fluid phosphorylated tau in both female and male memory clinic patients [140]. Additionally, these findings were affiliated with a decrease in learning ability that was tracked over a 6-year timeframe; this observation is in line with the idea that changes in the LC prior to the accumulation of neuropathology takes time to intensify to a measurable degree which allows us to discern behavioral differences [140]. Collectively, these studies indicate oxidative stress, which is regulated by estrogens and progesterones, metabolizing NA ultimately results in neuropathology. Upon consideration of these results and taking our hypothesis into account, it is of substantial importance to advance this research by investigating the effects of sex differences, such as females undergoing the menopausal transition, may have on NA oxidization and if this results in increased neuropathology load and neuronal death in the LC.
Modifying risk factors
Potential preventative measures that have vastly emerged in AD literature are those of brain reserve and cognitive reserve [141]. Brain reserve pertains to the individual’s anatomical brain structure, and suggests a more complex morphological framework that includes increased synaptic and neurotransmitter receptor densities that would potentially be able to withstand consequences of neuronal loss or tolerate more detriment. Alternatively, cognitive reserve spotlights individuals’ functional resilience and suggests increased cognitive processes or more intricate neural networks that aid in creating a higher degree of build-in redundancy [142]. Currently and collectively, both brain reserve and cognitive reserve highly favor males as opposed to females. For instance, brain reserve strongly suspects that males increased cerebral brain volume is superiorly capable of withstanding the accumulation of pathology to that of females [143, 144]. This is further supported by evidence indicating females had significantly greater odds of AD given the identical amount of pathology to males [145], as well as female brain volumes declining faster in patients with mild cognitive impairment and AD [146]. However, bearing in mind the previously discussed preeminence of the female rodent LC morphology and if this is translatable to female human LC morphology, what it means for female brain reserve remains to be answered.
Conversely, a link between the characteristics of cognitive reserve, such as education, occupational attainment, and lifetime experiences, and the upregulation of the LC-NA system in both animal models and in vivo human studies has been alluded to [147]. An upregulated LC-NA system allows for an increase in the number of receptors on the target cell, permitting more effective signal transmissions needed for peak cognitive performance, while simultaneously releasing a protective mechanism against threats of inflammation and neuropathology [12, 149]. While the mechanism of cognitive reserve is still difficult to define, the model suggesting NA being the fundamental pillar that underpins a delay in AD pathology is something to consider. It is widely acknowledged that males/men surpass females/women in cognitive reserve on the basis of both sex and gender differences due to previous societal practices which resulted in less opportunity for cognitive enrichment for females/women. Moreover, a study comparing LC signal intensity and the established proxies of reserve (i.e., education level and occupational attainment) in a cognitively healthy population, showed older-adult females, on average, have 20% less LC signal intensity than older-adult males [150]. Conversely, a more recent study observed no sex-differences in age-related LC intensity associations [151]. These conflicting results may be due to different sample sizes and/or varying parameters of magnetic resonance imaging, thus continuation of improving upon the methodological approach is required to validate such findings. Nonetheless, a potential sex difference in LC signal intensity raises the question of what this means in regards to an indisposed LC in menopausal females (i.e., during a marked time of the depletion of estrogens). Does this affect females/women’s day-to-day cognitive capabilities or their ability to undertake cognitive reserve, and therefore losing out on the potential to compensate or modify the AD state? If so, we must discover how we may begin to exploit females’ ability to perpetuate cognitive reserve, thereby upregulating the LC-NA system, earlier on in life, in order to mitigate the foreseeable AD pathology that comes as a result of the decline of estrogens and LC-NA dysregulation.
Pathogenic predisposition and sequence of progression
Structural and metabolic characteristics of female APOE ɛ4 gene carriers contribute to a predisposition to pathophysiological dysfunction, resulting in neuropathological components of AD. The composition of phenotypic, LC morphological framework induced by estrogens, and demographic attributes establish the substructure that influences LC dysfunction and neuronal loss that accelerates AD pathogenesis. In the course of female AD, it is more likely that the combined neuroprotective functions of NA, the inflammatory response system, and the regulation of the BBB, with the additional support of estrogens, are able to hold off the disease until a critical threshold. We speculate a drastic decrease in estrogens and NA production results in the pathologic process of tau pathology and Aβ so severe that it can be observed in BB stage V and VI through LC cell loss and impaired cognitive function. Moreover, an overall loss of tropic support occurs, thus leaving the female brain incapable of producing an anti-inflammatory response and governing BBB integrity, in turn conducing a toxic neuronal environment. Neuropathologies intensifying, BBB dysfunctioning, and neuroinflammation rising creates a continuous fed forward cycle that contributes to the progression of the disease, and causes clinical signs of AD such as a reduction of neuronal plasticity and neurodegeneration to become more apparent, and are currently unable to be halted or reversed (Fig. 3).
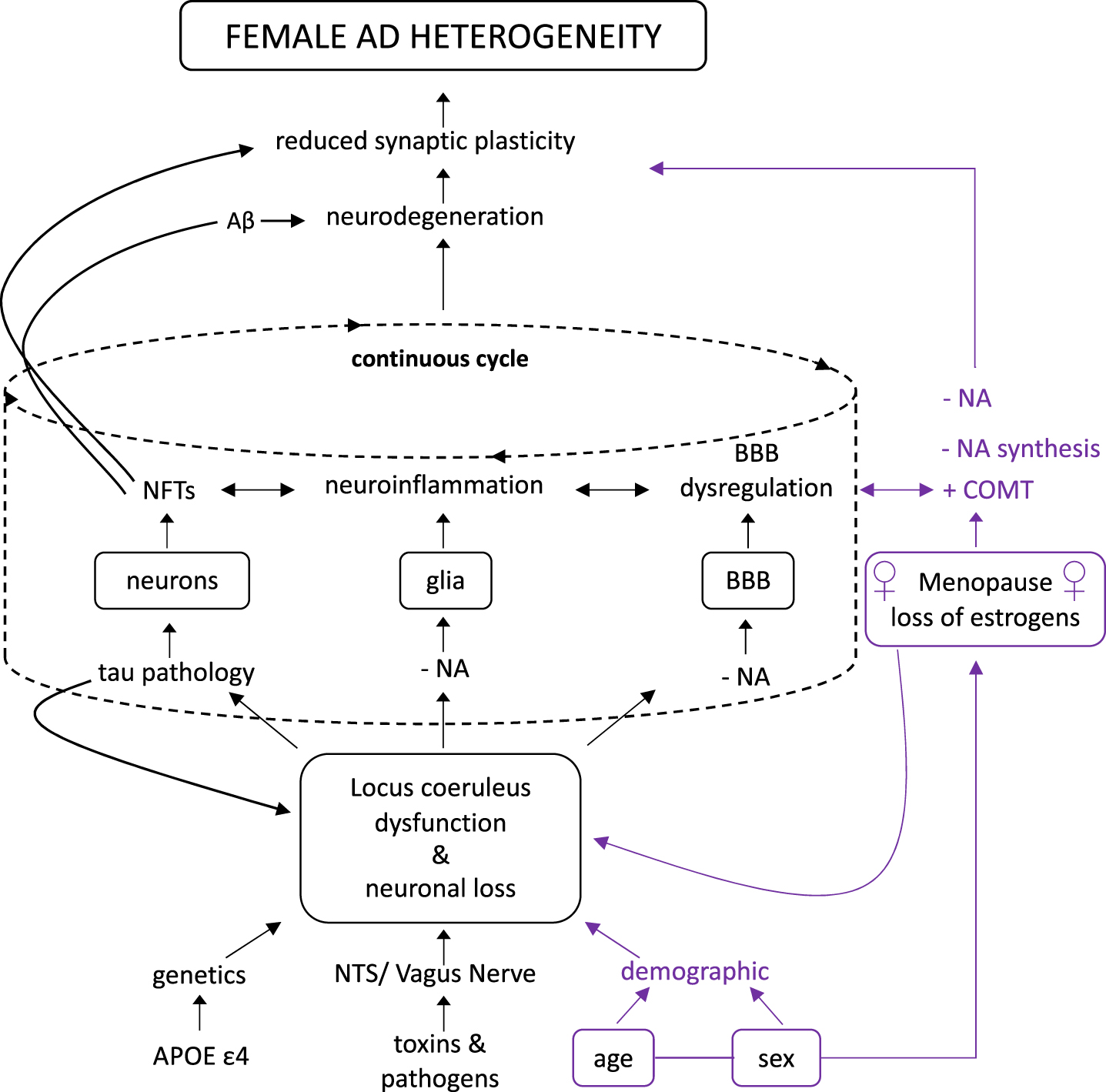
Hypothesized route of female ad heterogeneity. We propose that sex and age, as it relates to the onset of menopause, increases female vulnerability due to the rapid loss of estrogens and its effects on the LC-NA system, thus accelerating the load of neuropathology that give rise to neurodegeneration and reduced synaptic plasticity. This hypothesized route not only plays a critical role in AD development, but also disproportionately effects females. Redrawn from Mravec et al. 2014 [1].
Mixed pathology
The exploration into sex difference and their frequency patterns of mixed neuropathologies have only recently begun [137]. However, according to a recent study involving autopsy-based investigations on human brains, females significantly exhibited signs of mixed AD and cerebrovascular pathology compared to males [152]. This finding raises intriguing questions regarding the nature and extent of the role in which the two critical components of our present hypothesis have given our previous explanation of the vitality of the BBB being heavily dependent on favorable LC-NA signaling the production of estrogens. It is therefore likely that a further connection may exist between the downfall of both the female LC and estrogens and an increase in the comorbidity of mixed pathologies, thus warranting future studies.
Biomarkers
Currently, the amyloid/tau/neurodegeneration (ATN) classification framework has seen a significant rise in hopes that it will assist in the diagnostic procedure, development of preventative measures, and treatment strategies for AD. ATN classification for AD focuses on the presence of Aβ pathology, tau pathology, and neurodegeneration. Prior research utilizing the ATN biomarkers, as well as, most LC investigations have yet to report any significant difference among males and females; however, sex differences have not been the predominant variable of consideration. In line with ongoing research, we support moving in the direction of focusing investigations on the LC as a new biomarker potentially capable of tracking prognosis and progression of AD, with a specific interest in sex differences. It is essential that further efforts utilize various methodologies, such as unbiased stereological analysis to assess LC cell population, volume, and neuronal death [132]. Additionally, capitalizing on enhancements new technologies provide to current techniques such as in vivo imaging [153] and positron emission tomography imaging [154] will be a critical component in measuring LC integrity and functionality, allowing us to overcome previous obstacles such as its small size and location. Combining these various methodologies will further facilitate our ability to validate the LC as an early-stage biomarker for AD, and incite further exploration into sex differences.
Translation potential (diagnostic implications)
Future research in pursuance of advancing early disease detection and improving upon investigational therapeutic approaches are of utmost priority. Neuropsychological testing upon exhibitions of cognitive deficits is one the primary courses of action to assess if AD has begun to manifest. However, the current assessment strategies not adjusting for sex differences indicates a significant shortcoming in the clinical diagnosis of AD, and may contribute to a myriad of misdiagnoses or late diagnoses in females.
Cognitive abilities have recently been recognized in both animal and human studies as being molded by gonadal steroids and the differences by which LC-NA mechanisms modulate the amygdala and hippocampus regions, and throughout the brain [155]. These factors contribute to males and females having unique patterns of activation and performance on emotional autobiographical and episodic memory, spatial ability, and perceptual processing tasks [155]. It has been proposed that the mechanism of action by which the LC assists males and females differently is in how they distinguish and redirect attention to salient stimuli and how they encode and retrieve specific information. Such discrepancies become behaviorally apparent as males outperform females on recall tasks for central information, spatial rotation, and navigation tasks, while females excel on recall tasks for ancillary information, objection location, and verbal memory tasks [130, 155].
Acknowledging the role the LC and estrogens play in the sex-specific strengths and disadvantages consistently present in cognitive processes may have direct bearing on neuropsychological testing, so much so that it ought to be accommodated for in the formation of sex-adjusted normative data and sex-based cut-off scores used to differentiate normal aging and preclinical AD. Accordingly, this will increase the likelihood of preventing AD symptoms being masked during examination and further improve validity of AD diagnosis. Sex-specific neuropsychological assessments will have vast implications for the monitoring of AD progression, as well as, measuring the effectiveness of experimental treatments.
LINKAGE TO OTHER MAJOR THEORIES
Currently, there is no disease-modifying treatments available in spite of an enormous amount of research focused primarily on the role of Aβ plaques and neurofibrillary tangles in neurodegeneration in patients [156–158]. However, the underlying cause/causes of AD pathology remain at large, highlighting the critical urgency we must devote to researching new hypotheses such as the one presented in this article. The substantial amount of AD heterogeneity has led to the proposal of numerous theories with various focal points ranging from genetic predisposition, abnormal accumulation of pathogenic properties, dysregulation of the CNS, and environmental risk factors, implying a high probability of multiple pathways being involved in the clinical presentation and progression of AD. However, prior postulations have overlooked sex differences in AD heterogeneity. We put forth a hypothesis that we believe has pinpointed the LC-NA system as the hub of etiology in female AD, and furthermore distinguishes the decline in LC-NA and the production of estrogens as the initiating drivers that spur the primary concentration of the aforementioned hypotheses. Although our theory is not fully supported by experimental research, dissecting and analyzing formerly disseminated publications provided the evidence needed for this defining contribution that will now require others in the field to embrace and utilize to stimulate ideas for future studies. Conducting experiments aimed at maintaining estrogens and LC-NA production may prove to provide the pertinent evidence needed in order to find suitable therapeutic interventions.
CONCLUSION
To our knowledge, this is the first article that has provided a deeper insight into the interrelationship between the disruption of the female LC-NA system, the decline of estrogens, and AD vulnerability, seeing that no studies have been conducted considering their involvement to date. Accounting for sex as a biological variable in the LC as our hypothesis suggests, may propel AD research forward. These future studies will lay the groundwork for identifying sex differences and understanding their impact on AD etiology and progression. Furthermore, this will allow prevention trials to target the LC and enhance our ability to delay the onset or slow the progression of AD before it causes irrevocable neurodegeneration.
The LC contains 40,000–60,000 neurons that have a remarkably extensive axonal diffusion throughout the brain [159, 160], with its efferents consisting of two ascending fiber systems innervating 1) the cortex and 2) cerebellar forebrain, as well as, a descending pathway in the periventricular bundle projecting to the spinal cord [161–163]. The LC receives afferent projections from brainstem structures, in addition to inputs from the prefrontal cortex. These connections allow the LC to integrate low-level autonomic stimuli with high level cognitive information and to transfer this signal throughout the brain [164, 165].
Upon activation, NA is discharged from the LC and acts on both α- and β-adrenergic receptors present in neurons and glial cells, thus facilitating signal transduction and diverse functions within the central nervous system (CNS) [166], ranging from regulation of arousal and autonomic function to influencing episodic memory, attention, and working memory [6, 167–171]. NA also acts as a neuromodulator regulating synaptic connections and neuroplasticity, while also promoting neurogenesis and survival [70, 172]. NA additionally provides neuroprotection by suppressing neuroinflammation, and in under certain conditions, defending neurons from amyloid-induced toxicity, excitotoxicity, metabolic, and oxidative stress [12, 20, 52, 148, 149]. LC neuron terminals also possess and release co-transmitters, such as neuropeptide Y and galanin, concurrently with NA [173], both of which have been proven to contribute to neuronal plasticity and neuroprotection [174–177]