
Abstract
Alternative splicing of tau exon 10 generates tau isoforms with three or four microtubule-binding repeats, 3R-tau or 4R-tau, which are under developmental regulation. Dysregulation of tau exon 10 splicing is sufficient to cause neurodegenerative disorders. The RNA-binding Fox3 (Rbfox3), identified as NeuN, regulates RNA processing. However, whether Rbfox3/NeuN regulates tau exon 10 splicing is unknown. In the present study, we found that the developmental expression of 4R-tau coincided with the expression of Rbfox3 in rat brains. Rbfox3 enhanced tau exon 10 inclusion. Tau intron 10 contains UGCAUG, the conservative binding sequence of Rbfox3. Intron 10 of tau pre-mRNA was co-immunoprecipitated by Rbfox3/NeuN. Deletion mutants of the RNA recognition motif (RRM) or three RNA-binding sites of the RRM in Rbfox3/NeuN failed to enhance tau exon 10 inclusion. Rbfox3, specifically expressed in the fetal brain, did not affect tau exon 10 splicing. The level of Rbfox3/NeuN was reduced and was associated with the ratio of 4R-tau/3R-tau in the excitotoxic mouse brains induced by kainic acid. These findings suggest that Rbfox3/NeuN regulates the alternative splicing of tau exon 10 and that decreased Rbfox3/NeuN may lower the ratio of 4R-tau/3R-tau.
INTRODUCTION
Tau is a major neuronal microtubule-associated protein that plays an important role in the assembly and stabilization of microtubules. Tau is abnormally hyperphosphorylated and aggregated into neurofibrillary tangles (NFTs) in the brains of individuals with Alzheimer’s disease (AD) and related neurodegenerative diseases, collectively called tauopathies [1]. Adult human brain expresses six isoforms of tau generated from a single gene by the alternative splicing of exons 2, 3, and 10. Exon 10 encodes the second microtubule-binding repeat of tau. Inclusion and exclusion of tau exon 10 result in tau isoforms with four or three microtubule-binding repeats, called 4R-tau or 3R-tau, respectively. 4R-tau and 3R-tau are approximately equally expressed in adult human brain [2, 3]. Imbalanced levels of 4R-tau and 3R-tau are observed in several tauopathies [4, 5], suggesting that the regulation of tau exon 10 is critical for maintaining normal function of the brain. The RNA-binding Fox (Rbfox) belongs to the heterogeneous nuclear ribonucleoproteins (hnRNPs) family and was characterized recently as a phylogenetically conserved regulator of alternative splicing [6–8]. The Rbfox family in mammals comprises three members: Rbfox1, Rbfox2, and Rbfox3. Rbfox3 was identified as NeuN and is expressed exclusively in neurons [9, 10]. Rbfox3 protein is composed of an RNA recognition motif (RRM), a nuclear localization signal (NLS), and a FOX1-C domain at the C-terminus (Fig. 1A). The truncation of the FOX1-C domain might be associated with its pathogenicity [8]. However, whether Rbfox3 regulates tau exon 10 alternative splicing is unknown.
In the present study, we found that the developmental expression of 4R-tau in rat brains coincided with the expression of Rbfox3. Rbfox3 acted on intron 10 of tau pre-mRNA and enhanced tau exon 10 inclusion. The level of Rbfox3 was positively correlated with the ratio of 4R-tau/3R-tau in excitotoxic mouse brains induced by kainic acid (KA). These results suggest that Rbfox3 may be involved in the regulation of tau exon 10 splicing.

Expression of Rbfox3 and tau in the rat brain during development. A) Diagram of three Rbfox3 isoforms: Rbfox3L (containing a 47-amino acid insert), Rbfox3S, and Rbfox3 (lacking exon 8, 31-amino acid). B) The levels of Rbfox3, 3R-tau, 4R-tau, and total tau in rat brains were analyzed by western blots developed with the indicated antibodies. The blots shown in panel B were quantified densitometrically, and the mean values of Rbfox3 (L + S) (C), Rbfox3 (D), or 4R-tau (E) at each time point were plotted against ages.
MATERIALS AND METHODS
Plasmids, antibodies, and other reagents
pCMV6-Entry/Rbfox3L and S (transcript variants 1 and 2) tagged with Myc and Flag at C-terminus were bought from Origene (Rockville, MD, USA). The plasmids of deletion mutations of Rbfox3 were generated by polymerase chain reaction (PCR) amplification from pCMV6/Rbfox3 and confirmed by DNA sequence analysis. pCI/SI9-LI10 contains a tau mini-gene, SI9-LI10, comprising tau exons 9–11, part of intron 9, and the full length of intron 10 [11]. Short interfering RNA (siRNA) of mouse Rbfox3 (a pool of three siRNAs targeting on sequences 001: CTCATCCCACCCAGGACTA, 002: AGATGAAGCAGCACAGACA, 003: CGACTACATGTCTCCAACA) were synthesized by RIBOBIO Company (Beijing, China). Monoclonal anti-NeuN (Rbfox3) antibody (MAB377), anti-3R tau, and anti-4R tau were from Millipore (Billerica, MA, USA). Polyclonal anti-total tau (R134d) was produced by our laboratory. Monoclonal anti-Flag and anti-actin were from Sigma (St. Louis, MO, USA). Polyclonal anti-GAPDH was from Santa Cruz Biotechnology (Santa Cruz, CA, USA). Protein G beads and the ECL western blotting substrates were from Thermo Fisher Scientific (Rockford, IL, USA). Peroxidase-conjugated anti-mouse and anti-rabbit IgG were obtained from Jackson ImmunoResearch Laboratories (West Grove, PA, USA).
Rats and mice brain tissues
Developmental rat brain homogenates were from previous study [12]. Rat brain tissues were collected from rat fetuses at gestation day 15 and 19 (E15 and E19), rat pups on the day of birth (P0), at postnatal day 5 (P5), P15, P30, and postnatal 3 months (3M), 6M, 12M, and 24M [12].
Excitotoxic mouse brain homogenates were from a previous study [13]. The mice (male, 25–30 g body weight, 12 weeks old) were injected intraperitoneally with KA (TOCRIS, Minneapolis, MN, USA; dissolved in isotonic saline, 20 mg/kg of body weight), and sacrificed 0, 2.5, 6, 10, 24, and 36 h after injection, and the forebrains were immediately removed. Tissues were stored at –80°C until used.
All animal handling and use were as per the protocol approved by our Institutional Animal Care and Use Committee, in accordance with the PHS Policy on Human Care and Use of Laboratory Animals.
Brain tissues were homogenized in 9 volumes of cold buffer consisting of 50 mM Tris-Cl, pH 7.4, 8.5% sucrose, 50 mM NaF, 1 mM Na3VO4, 2.0 mM EDTA, 10 mM β-mercaptoethanol, and 10μg/ml each of aprotinin, leupeptin, and pepstatin. Aliquots of the homogenates were mixed with the same volume of 2×Laemmeli buffer (125 mM Tris-Cl, pH 6.8, 4% sodium dodecyl sulfate (SDS), 20% glycerol, 2% β-mercaptoethanol and 0.005% bromophenol blue), followed by heating in boiling water for 5 min.
Cell culture and transfection
HEK-293T cells, mouse neuroblastoma N2a cells, or human neuroblastoma SH-SY5Y cells were maintained in Dulbecco’s modified Eagle’s medium (DMEM) or DMEM/F12 supplemented with 10% fetal bovine serum (FBS) (Invitrogen, Carlsbad, CA, USA) at 37°C (5% CO2). All transfections were performed with FuGENE ® HD (Promega, Madison, WI, USA) or Lipofectamine™ 2000 (Invitrogen) according to the manufacturer’s instructions.
Knockdown of TDP-43 with RNA interference
For inhibition of Rbfox3 expression, N2a cells were transfected with certain amounts of siRNAs, siRbfox3 (RIBOBIO), using Lipofectamine 2000. After 48 h transfection, RNA and proteins were extracted as described below. The same amount of scramble siRNA was used as control.
Quantitation of tau exon 10 splicing by reverse transcription-PCR
Total cellular RNA was extracted from cultured cells by using the RNeasy mini kit (Qiagen, GmbH, Germany) per the manufacturer’s instructions. One microgram of total RNA was used for first-strand cDNA synthesis with oligo-(dT) by using the Omniscript reverse transcription kit (Qiagen). PCR was performed by using PrimeSTAR HS DNA Polymerase (Takara Bio Inc., Otsu, Shiga, Japan) with forward primer 5′-GGTGTCCACTCCCAGTTCAA-3′ and reverse primer 5′-CCCTGGTTTATGATGGATGTTGCCTAATGAG-3′ to measure the alternative splicing of tau exon 10. The conditions of PCR were: 98°C for 5 min, 98°C for 10 s, 68°C for 40 s for 30 cycles, and then 68°C for 10 min for extension. The PCR products were resolved on 1.5% agarose gels, visualized by ethidium bromide staining and quantitated using the Molecular Imager system (Bio-Rad).
RNA immunoprecipitation
The RNA immunoprecipitation (RNA-IP) experiment was performed as described [14, 15]. Briefly, N2a cells co-transfected with pCMV6/Rbfox3 and pCI/SI9-LI10 or SH-SY5Y cells transfected with pCMV6/Rbfox3 were crosslinked with 1% formaldehyde for 10 min at room temperature. After quenching with 125 mM glycine, the cells were lysed in lysis buffer (16.7 mM Tris-Cl, pH 8.1, 0.01% SDS, 1.1% Triton X-100, 1.2 mM EDTA, 167 mM NaCl, 1×Roche protease inhibitors cocktail and 50 U/ml RNasin Plus RNase Inhibitor) on ice for 10 min and centrifuged at 2,000 g for 5 min to pellet nuclei. The nuclear fraction was sonicated in buffer B (50 mM Tris-Cl, pH 8.1, 1% SDS, 10 mM EDTA, 1×protease inhibitors cocktail and 50 U/ml RNasin Plus RNase Inhibitor). After centrifugation at 16,000 g for 10 min, the supernatant was subjected to immunoprecipitation with anti-Flag in IP buffer (16.7 mM Tris-Cl, pH 8.1, 167 mM NaCl, 0.01% SDS, 1.1% Triton X-100, 1.2 mM EDTA, 1×protease inhibitors cocktail and 50 U/ml RNasin Plus RNase Inhibitor) for 2 h. Immuno-complex was washed sequentially with low-salt buffer (20 mM Tris-Cl, pH 8.1, 150 mM NaCl, 0.1% SDS, 1% Triton X-100 and 2 mM EDTA), with high-salt buffer (20 mM Tris-Cl, pH 8.1, 500 mM NaCl, 0.1% SDS, 1% Triton X-100 and 2 mM EDTA), with LiCl buffer (10 mM Tris-Cl, pH 8.1, 250 mM LiCl, 1% NP-40, 1% deoxycholate and 1 mM EDTA), and with TE buffer (10 mM Tris-Cl, pH 8.0, 1 mM EDTA). Immuno-complex was eluted with elution buffer (1% SDS, 0.1 M NaHCO3 and 50 U/ml RNasin @ Plus RNase Inhibitor). The crosslinking was reversed by incubation with 200 mM NaCl at 65°C for at least 2 h. After digestion with 0.4 mg/ml Proteinase K (Invitrogen) at 42°C for 45 min and 1 mg/ml of RQ1 RNase-free DNase (Promega) at 37°C for 15 min, respectively, RNA was extracted by miRNeasy Micro Kit (Qiagen) and was subjected to first-strand cDNA synthesis with random primers by using the Omniscript Reverse Transcription Kit (Qiagen). The cDNA was amplified by PrimeSTAR HS DNA Polymerase (Takara) with primers against tau intron 10: Forward 5′-AAAATCAGGGGATCGCAGCGGCTAC-3′, Reverse 5′-GACATTCTTCAGGTCTGGCATGGGCAC-3′. The conditions of PCR were initial denaturation at 98°C for 5 min, 98°C for 10 s, 68°C for 40 s for 30 cycles, and a final extension at 68°C for 10 min. PCR products were separated on a 2% agarose gel and were visualized by ethidium bromide staining.
Western blots
Cells or brain homogenates were lysed in 1×Laemmli sample buffer (62.5 mM Tris-Cl, pH 6.8, 2% SDS, 10% glycerol, 5% 2-mercaptoethanol, 0.002% bromophenol blue) and boiled for 5 min. The protein concentration of cell lysates was measured by using the Pierce 660 nm protein assay kit (Thermo Fisher Scientific). Equal amounts of protein were separated by SDS-polyacrylamide gel electrophoresis (PAGE) and were electrically blotted onto polyvinylidene difluoride membrane. The membrane was blocked with 5% fat-free milk in TBS for 30 min and then incubated with primary antibody in 5% fat-free milk and 0.5% NaN in TBS overnight at room temperature. After washing three times with TBST (TBS with 0.05% Tween 20), the membrane was incubated with horseradish peroxidase-conjugated secondary antibodies for 2 h. After three washes with TBST, the membrane was incubated with ECL for 1 min and then exposed to X-ray films (Kodak, Rochester, NY, USA).
Statistical analysis
The data are presented as mean±SD and were statistically analyzed by the unpaired two-tailed Student’s t test for two-group comparison and by one-way ANOVA for multiple-group comparison. Non-linear regression was used for analysis of the correlation between the ratio of 4R-tau/3R-tau and level of Rbfox3. A value of p < 0.05 was considered statistically significant.
RESULTS
Expression of Rbfox3 coincides with 4R-tau expression in rat brains
It is well known that alternative splicing of tau exon 10 is regulated developmentally. We observed that embryonic and neonatal rat brain expressed 3R-tau (Fig. 1B). In adult rat brain, 3R-tau was not detectable (Fig. 1B). Rat brain started to express 4R-tau at P15. 3R-tau was decreased and 4R-tau was increased in rat brain during development (Fig. 1B, E).
Three variants of Rbfox3—Rbfox3L, Rbfox3S, and Rbfox3 (Fig. 1A)— are expressed in the brain [10, 16]. To determine developmental expression of Rbfox3, we analyzed the levels of Rbfox3 in rat brains from embryonic to aged rats by western blots developed with anti-NeuN. We found that the expression of Rbfox3 was also regulated developmentally (Fig. 1B). Rbfox3L and Rbfox3S were undetectable in embryonic rat brain (Fig. 1B). Rat brain started to express Rbfox3S at P0 and Rbfox3L at P30 (Fig. 1B). The levels of Rbfox3S and Rbfox3L in rat brains gradually increased during development (Fig. 1B, C). Rbfox3 was highly expressed in embryonic rat brains, but decreased after birth (Fig. 1B, D). Thus, the expression of Rbfox3L/S coincided with the expression of 4R-tau in the rat brain during development (Fig. 1B, C, E), suggesting that Rbfox3 may be involved in regulation of the alternative splicing of tau exon 10.
Rbfox3 enhances tau exon 10 inclusion
To determine whether Rbfox3 regulates the alternative splicing of tau exon 10, we transfected various amounts of pCMV/Rbfox3L and pCMV/Rbfox3S together with pCI/SI9-LI10 (mini-tau gene consisting of tau exons 9, 10, and 11, and part of intron 9 and the full-length of intron 10) (Fig. 2A) into HEK-293T cells. We analyzed the splicing products of tau exon 10 by reverse transcription PCR (RT-PCR) and the expression of Rbfox3L or Rbfox3S by western blots 48 h after transfection. We found that overexpression of both Rbfox3S (Fig. 2B) and Rbfox3L (Fig. 2C) enhanced tau exon 10 inclusion in a dose-dependent manner. Both variants showed similar function in promoting tau exon 10 inclusion (Fig. 2B, C).
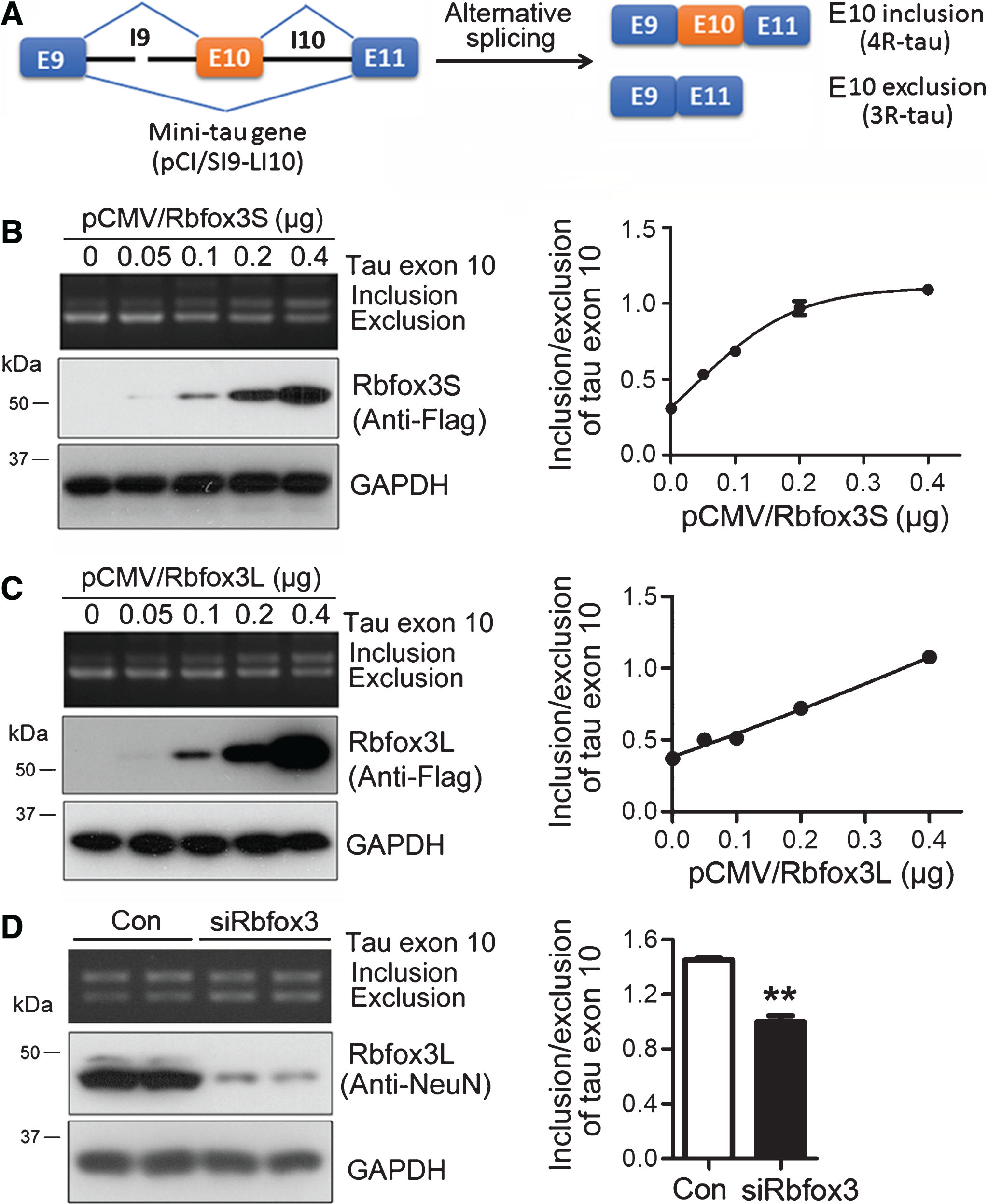
Rbfox3 enhances tau exon 10 inclusion. A) Ideograph of tau mini gene SI9-LI10. B,C) Various amounts of pCMV/Rbfox3S (B) and pCMV/Rbfox3L (C) were transfected together with pCI/SI9-LI10 into HEK-293T cells for 48 h. The alternative splicing products of tau exon 10 were analyzed by RT-PCR, and the ratio of inclusion/exclusion of tau exon 10 was plotted against the corresponding amounts of pCMV/Rbfox3 transfected into the cells. The expression of Rbfox3 tagged with Flag was detected by western blots. D) siRbfox3 and scramble siRNA (Con) was transfected together with pCI/SI9-LI10 into N2a cells. The splicing products of tau exon 10 were determined by RT-PCR, and the expression of Rbfox3 was detected by western blots 48 h after transfection. The ratio of inclusion/exclusion of tau exon 10 was analyzed and is represented as mean±SD (n = 3). *p < 0.05.
To confirm the role of Rbfox3 in tau exon 10 splicing, we knocked down Rbfox3 with its siRNA in neuroblastoma N2a cells-transfected with pCI/SI9-LI10 and analyzed the splicing products of tau exon 10. We found that knockdown of Rbfox3 suppressed tau exon 10 inclusion (Fig. 2D), supporting the premise that Rbfox3 enhances tau exon 10 inclusion.
Rbfox3 binds to UGCAUG in intron 10 of tau pre-mRNA
Rbfox proteins regulate alternative splicing by binding to a specific UGCAUG element [6, 18]. We found that tau intron 10 contains the hexa-nucleotide sequence (Fig. 3A). To determine if Rbfox3 binds to the tau intron 10, we performed RNA-IP. We co-transfected pCI/SI9-LI10 with pCMV/Rbfox3L into N2a cells and immunoprecipitated Rbfox3 with anti-Flag. The pre-mRNA of tau co-immunoprecipitated with Rbfox3 by anti-Flag was amplified with RT-PCR by using random primers for reverse transcription and primers against intron 10 of tau for PCR, as indicated in Fig. 3A. We found that intron 10 of tau pre-mRNA was co-immunoprecipitated by Rbfox3L (Fig. 3B), suggesting that Rbfox3 may interact with intron 10 of tau pre-mRNA.
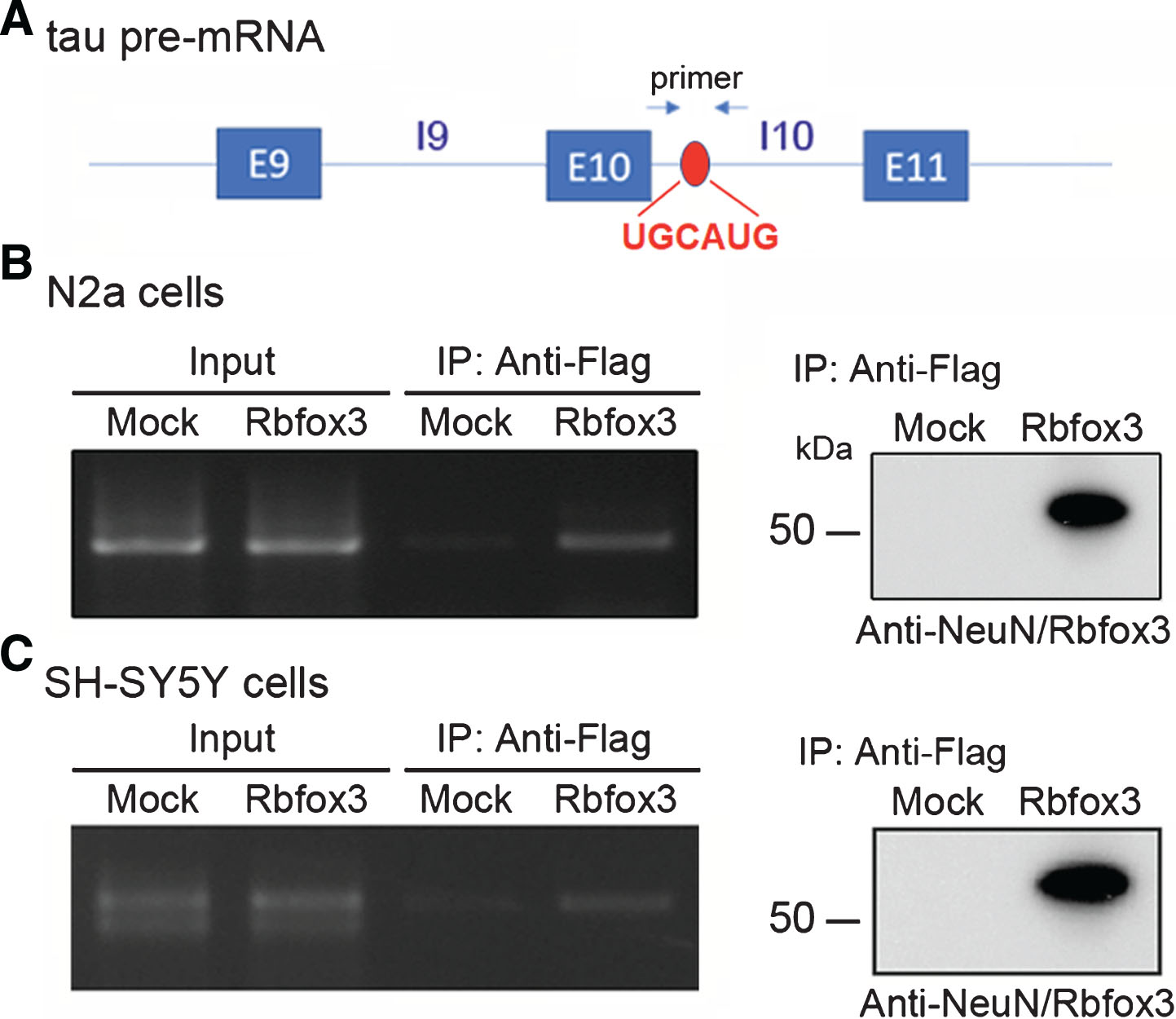
Rbfox3 binds to intron 10 of tau pre-mRNA. A) Schematic showing Rbfox3-binding element UGCAUG (red) of tau pre-mRNA. B) pCMV/Rbfox3L-Flag transfected together with pCI/SI9-LI10 into N2a cells, followed by immunoprecipitation with anti-Flag antibody. Co-immunoprecipitated pre-mRNA of tau with Rbfox3L was determined by RT-PCR with primers specific to intron 10, as indicated. The RT-PCR products were separated by agarose electrophoresis (left). The expression of Rbfox3L-Flag was determined by western blots with anti-NeuN (Rbfox3). C) pCMV/Rbfox3L-Flag was transfected into SH-SY5Y cells, the RNA-IP was carried out with anti-Flag, and the intron 10 of endogenous tau pre-mRNA was analyzed by RT-PCR, as described in panel B.
To confirm the interaction of Rbfox3 with tau intron 10, we overexpressed Rbfox3L in human neuroblastoma SH-SY5Y cells and performed RNA-IP as described above. The results showed that intron 10 of endogenous tau pre-mRNA was co-immunoprecipitated by Rbfox3L (Fig. 3C), confirming the interaction of Rbfox3 with intron 10 of tau pre-mRNA.
RRM is required for enhancing tau exon 10 inclusion
To determine the role of different domains of Rbfox3 in tau exon 10 splicing, we constructed several deletion mutants of Rbfox3 (Fig. 4A) and co-transfected them into HEK-293T cells with pCI/SI9-LI10 for 48 h. We found that the full-length Rbfox3 enhanced tau exon 10 inclusion, and the RRM deleted mutants Rbfox3 and Rbfox3 failed to enhance tau exon 10 inclusion. Rbfox3 and Rbfox3 showed similar ability to full-length Rbfox3, whereas Rbfox3 and Rbfox3 which do not have an NLS, showed less ability in the promotion of tau exon 10 inclusion (Fig. 4B). Although the expression levels of Rbfox3 mutants were various and very lower levels of Rbfox3 and Rbfox3 were detected (Fig. 4B), Rbfox3 could not enhance, and Rbfox3 and Rbfox3 enhanced to a lesser degree, tau exon 10 inclusion, suggesting that the RRM is required for, and the NLS may affect Rbfox3 to promote, tau exon 10 inclusion.
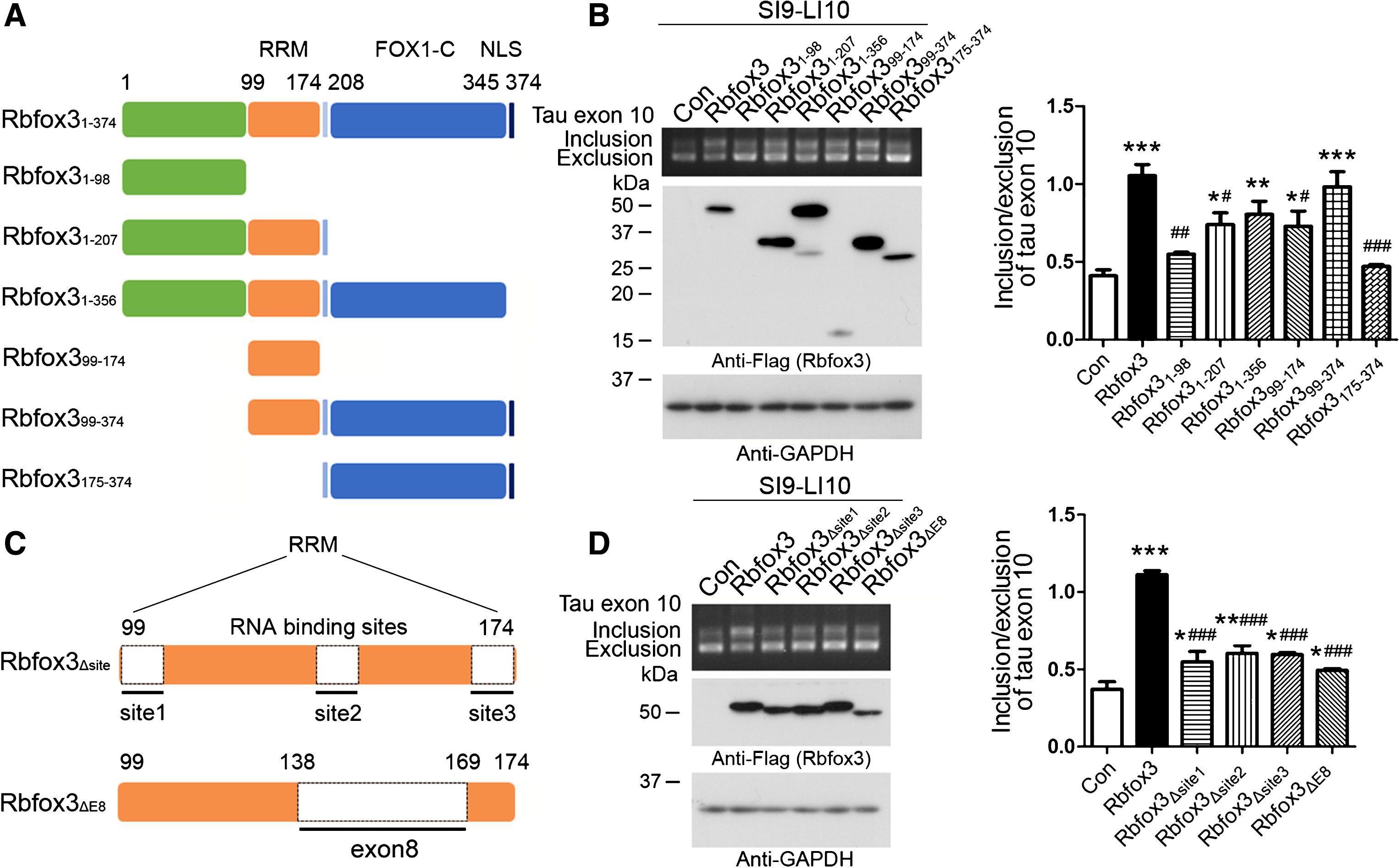
RRM of Rbfox3 is required for promotion of tau exon 10 inclusion. A) Schematic diagram of Rbfox3 deletion mutants. Rbfox3 protein is composed of an RRM in the central region of the molecule, an NLS and a FOX1-C domain at the C-terminus. B) The plasmids of the deletion mutants of Rbfox3 tagged with Flag were co-transfected with pCI/SI9-LI10 into HEK-293T cells. The splicing products of tau exon 10 were analyzed by RT-PCR. The expressions of Rbfox3 mutants were analyzed by western blots developed with anti-Flag. C) Schematic diagram showing the deletion mutations of the RNA-binding sites of RRM and exon 8 of Rbfox3. D) The deletion mutants of RNA-binding sites of RRM or exon 8 of Rbfox3 were co-transfected with pCI/SI9-LI10 into HEK-293T cells. The splicing products of tau exon 10 and Rbfox3 mutants were analyzed by RT-PCR. The ratio of tau exon 10 inclusion and exclusion are represented as mean±SD. p < 0.05; p < 0.01; p < 0.001; *versus Con; versus Rbfox3.
There are three RNA-binding sites in the RRM domain of Rbfox3 (Fig. 4C). Exon 8 encodes part of the RRM, which contains the second RNA-binding site (Fig. 4C). To determine their effect on tau exon 10 inclusion, we deleted each of three RNA-binding sites of RRM and exon 8, respectively (Fig. 4C) and co-transfected them with pCI/SI9-LI10 into HEK-293T cells for 48 h. We found that deletion any one of RNA-binding sites and deletion of exon 8 significantly reduced the activity to promote tau exon 10 inclusion as compared to the full-length Rbfox3 (Fig. 4D). These results suggest that all three RNA-binding sites of RRM are critical for Rbfox3’s function on tau exon 10 inclusion. Rbfox3 may not regulate the alternative splicing of tau exon 10.
Reduction of Rbfox3 expression is associated with the decrease of 4R-tau in excitotoxic mouse brains
We previously reported that in excitotoxic mouse brains induced by KA, 4R-tau is decreased [19], leading to a decreased ratio of 4R-tau/3R-tau, in a time-dependent manner (Fig. 5C). To determine the relationship between Rbfox3 and dysregulation of tau exon 10 splicing in excitotoxic mouse brains, we analyzed the expressions of Rbfox3 by western blots and found that Rbfox3 was markedly decreased in a time-dependent manner (Fig. 5A, B). The time-dependent reduction of Rbfox3 coincided with the reduction of the ratio of 4R-tau/3R-tau (Fig. 5C). Correlation analysis indicated a strong positive correlation between the ratio of 4R-tau/3R-tau and the level of Rbfox3 (Fig. 5D). These results suggest that the decreased Rbfox3 may contribute to the decreased ratio of 4R-tau/3R-tau in the excitotoxic mouse brains and support the hypothesis that Rbfox3 may regulate tau exon 10 splicing in the brain.
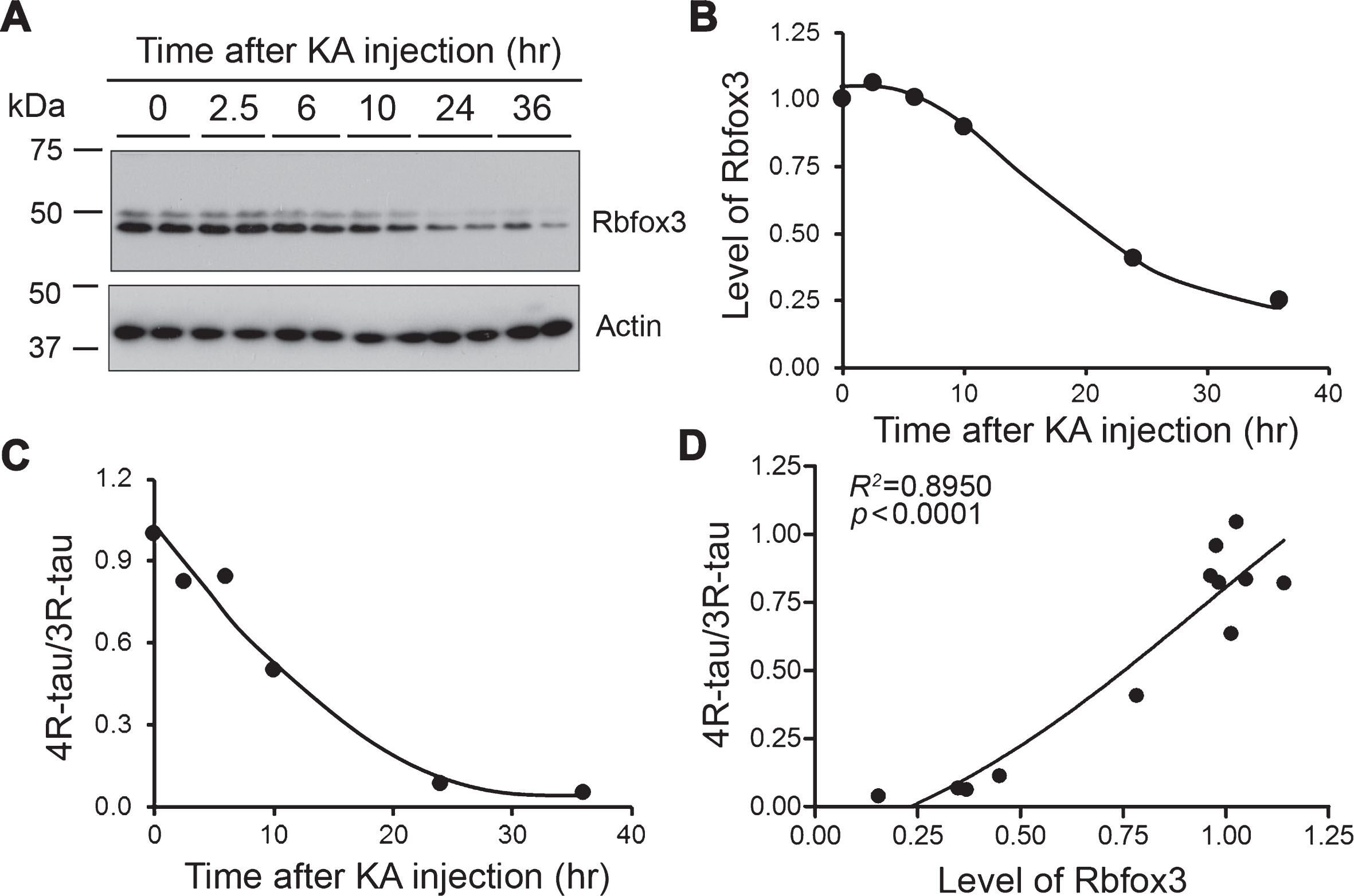
Decreased expression of Rbfox3 level is associated with the reduction of 4R-tau/3R-tau ratio in excitotoxic mouse brains. A, B) The levels of Rbfox3 in the brain homogenates from KA injected FVB mice were analyzed by western blots developed with anti-NeuN (A). The quantifications of the blot are shown in (B). C) Relative levels of 4R-tau and 3R-tau were determined by western blots previously [19], and the ratios were plotted against the time after KA injection. D) 4R-tau/3R-tau ratio was plotted against the level of Rbfox3. Non-linear correlation analysis was performed.
DISCUSSION
Alternative splicing of tau exon 10 results in 4R-tau and 3R-tau, which are equally expressed in normal adult human brain [2, 3]. Dysregulation of tau exon 10 splicing is associated with several neurodegenerative disorders related to tau pathology [4, 5]. In the present study, we report for the first time that Rbfox3/NeuN regulated tau exon 10 splicing. In developmental rat brains, Rbfox3 expression coincided with 4R-tau expression. Tau intron 10 contains UGCAUG, a conservative Rbfox-binding site. Rbfox3 interacted with intron 10 of tau pre-mRNA. Overexpression of Rbfox3L and Rbfox3S, but not of the fetal form, Rbfox3ΔE8, enhanced tau exon 10 inclusion, which required RRM. In excitotoxic mouse brains, decreased Rbfox3 was associated with a decreased ratio of 4R-tau/3R-tau. Thus, Rbfox3 may play an important role in tau exon 10 splicing.
The three Rbfox paralogues are a highly conserved family of alternative splicing regulators, all of which contain a single RRM-type RNA binding domain near the center of the protein [20]. Rbfox1 is expressed in brain and striated muscles, and Rbfox2 is expressed ubiquitously, whereas Rbfox3 expression is restricted to neurons [7, 21]. Rbfox3 is highly conserved among species and is stably expressed during specific stages of development [20]. Rbfox3 contains an RRM and is classified as a member of the Fox-1 gene family that binds specifically to the identical hexa-nucleotide, UGCAUG, and functions as a splicing regulator [10]. UGCAUG is overrepresented in the introns, in which splicing is regulated in a cell type- and developmental stage-specific manner [22–25]. Alternative splicing occurs most frequently in the brain [26, 27]. It was reported that Rbfox3 can activate neuron- and muscle-specific alternative splicing of a cassette exon, N30, of the NMHC II-B pre-mRNA [10], exon EIII-B of fibronectin [28], exon N1 of c-src, exon 33 of the L-type calcium channel Cav1.2 [29] and that Rbfox3-regulated alternative splicing of Numb promotes neuronal differentiation during development [8]. Alternative splicing regulated by Rbfox3 would play a key role in the regulation of neural cell differentiation and development of the nervous system. We found here for the first time that as a neuron-specific splicing factor, Rbfox3 enhances tau exon 10 inclusion.
Tau exon 10 splicing is developmentally regulated. Fetal and neonatal brains only express 3R-tau, and mature murine brains mainly express 4R-tau. Three isoforms of Rbfox3 are expressed in the brain, which are developmentally regulated in rat brain as well. We found here that fetal and neonatal rat brain expressed Rbfox3ΔE8, which was dramatically decreased after birth. Rat brain started to express Rbfox3S at P5, and Rbfox3L at P30. Both Rbfox3S and Rbfox3L were gradually increased during development. The total level of Rbfox3 was increased during development and coincided with 4R-tau expression, suggesting that Rbfox3 may be involved in the developmental regulation of tau exon 10, which remains to be determined.
Alternative splicing of tau exon 10 is regulated by several splicing factors, including 9G8, SC35, ASF/SF2, SRp55, Tra2-β1 and SRp30c, and TDP-43 [15, 30–35]. In the present study, we found that both Rbfox3L and Rbfox3S promoted tau exon 10 inclusion. The deletion of RRM domain or RNA-binding sites of RRM abolished or diminished its function in promoting tau exon 10 inclusion, suggesting that RRM is required for Rbfox3 to regulate tau exon 10 splicing. Exon 8 of Rbfox3 encodes part of RRM. Rbfox3, an isoform expressed in fetal and neonatal brain, failed to regulate tau exon 10 splicing. Tau intron 10 contains an UGCAUG element, a specific binding site of Rbfox [8]. We found that Rbfox3 bound to the UGCAUG in intron 10 of tau pre-mRNA. Thus, we speculate that Rbfox3 enhances tau exon 10 inclusion by acting on the UGCAUG element of tau pre-mRNA by its RRM.
Glutamate receptor-mediated excitotoxicity has been implicated in the pathogenesis of AD [36]. Rbfox3 in KA-induced excitotoxic mouse brain was decreased in a time-dependent manner, and decreased Rbfox3 was associated with a reduced ratio of 4R-tau/3R-tau. In AD brain, Rbfox3/NeuN is reduced as a result of neurodegeneration. We previously reported that the ratio of 4R-tau/3R-tau was decreased in AD brain [19]. Thus, we speculate that a reduced level of Rbfox3 might contribute to dysregulation of tau exon 10 splicing and tau pathogenesis in AD brain.
In conclusion, we found that Rbfox3 bound to UGCAUG in intron 10 of tau pre-mRNA and promoted tau exon 10 inclusion. Fetal Rbfox3 had no effect on tau exon 10 splicing. Developmental expression of Rbfox3 coincided with 4R-tau expression in rat brain. Reduction of Rbfox3 was associated with the decrease of 4R-tau/3R-tau ratio in excitotoxic mouse brain. Thus, Rbfox3 might be involved in the regulation of tau exon 10 splicing and reduction of Rbfox3/NeuN may play a role in tau pathogenesis in AD.