
Abstract
The accumulation of tau protein aggregates is a pathological hallmark in Alzheimer’s disease (AD) and other neurodegenerative diseases. However, the identity of the toxic tau conformation that propagates and induces neurodegeneration is still unknown. Anti-tau antibodies are a common tool used to differentiate between normal and pathological-associated tau forms or as passive immunotherapy in the quest to interfere with tau-mediated neurodegeneration. Here, we show that Tau13, a tau N-terminal antibody, preferentially enriches high molecular weight tau species produced in a tauopathy mouse model and AD. The data suggest that Tau13 has higher affinity to specific tau conformation presence in higher molecular weight tau species.
INTRODUCTION
The accumulation of aggregated tau proteins is a pathological hallmark of a group of neurodegenerative diseases collectively known as tauopathies [1–3]. The best known tauopathy is Alzheimer’s disease (AD) [1–3]. In AD, progressive accumulation of pathological tau proteins leads to its interneuronal transfer and, consequently, propagation of tau-mediated neurodegeneration, which is directly related to the severity of the clinical presentation [3, 4–6]. However, an important conundrum related to the pathophysiology associated with tau-mediated neurodegeneration is deciphering the conformational changes that convert tau into a toxic molecule.
Oligomeric tau proteins are considered the most neurotoxic species, whereas neurofibrillary tangles may play a neuroprotective role [7, 8]. Aberrant posttranslational modifications and protein-protein interactions may act in concert to promote the formation of pathological tau species [3]. For example, FKBP51/Hsp90 chaperone complex block UPS-mediated tau degradation and induce the formation of oligomeric tau, promoting tau-mediated neurotoxicity [9]. In contrast, previous studies showed that deletion of the RNA-binding proteins (RBPs) T-cell intracellular antigen 1 (TIA1) gene increased survival of mice expressing human P301S tau mutant, decreasing the accumulation of tau oligomers, but increased neurofibrillary tangles [10]. In the absence of TIA1, other protein(s) may associate with tau oligomers to promote the formation of filaments and tangles [10, 11]. However, it is still unclear the sequence of molecular events that promote the transition from normal to pathological tau species.
In AD, the accumulation of high molecular weight (HMW) tau species correlate with the accumulation of pathological tau in specific brain regions [3, 12]. Additionally, specific anti-tau antibodies have been used for the histological identification of specific tau conformations associated with AD pathology [12, 13]. Alz50 and MC1are antibodies commonly used in neuropathological analyses [13]. These antibodies recognized conformational-specific epitopes that are associated with the transition from normal to pathological form of tau [13]. TNT1 and TNT2 are other anti-tau antibodies that recognize an epitope in tau’s N-terminal domain that becomes exposed as pathological tau is formed [14–16]. These antibodies are mostly used in immunohistochemical analyses, where proteins present in the tissue are fixed and, in some instances, specific conditions are used to promote epitope presentation. Here, we show that Tau13, an antibody that binds to amino acids 2–18 and that is normally considered to bind both normal and pathological tau in western blot and immunohistological analyses, preferentially immunoprecipitates HMW tau from a tauopathy mouse model and AD brain. The results suggest that in solution Tau13 antibodies have higher affinity for specific tau conformations present in HMW tau species. Importantly, the results bring to the attention the use of different techniques to study specific tau conformational changes.
MATERIAL AND METHODS
Mouse colony
The transgenic (Tg) JNPL3 tauopathy mouse model was kindly provided by Drs. Michael Hutton and Jada Lewis [17, 18]. This tauopathy mouse model expresses the human tau isoform 0N4R bearing the P301L mutation, commonly found in familial cases of FTDP-17, under the control of the prion promoter. The JNPL3 mouse colony is maintained as hemizygous for htauP301L and inbred on the SW genetic background. JNPL3 mice develop tau-mediated neurodegeneration in age- and brain region-dependent manner, with a severe motor impairment phenotype. Motor impairment is evaluated using three tasks, namely tail hang, rope hang, and righting reflex tests. Mice with a motor impairment score equal to or below 10 are catalogued as normal, whereas mice with a score of 12 or above are catalogued as motor impaired. Motor impaired mice are sub-catalogued in three groups: mild (12–17), moderate (18–23), and severe (24–30) [18]. All experiments were conducted in accordance with Michigan State University IACUC approved protocol #01-15-020-00.
Human tissue
Human samples were derived from University of Michigan Brain Bank. Normal aging and AD temporal cortices were used in the experiments. The normal aging (NA) cohort had a Braak stage of 0, while AD cohort had a Braak stage of VI. The PMI was less than 6 h. Tissue samples from temporal cortex were excised, avoiding blood vessels and white matter.
Immunoprecipitation
Transgenic JNPL3 brains (500 mg) and AD temporal cortex (300 mg) were homogenized in Buffer A (20 mM Tris base, pH 7.4, 150 mM NaCl, 1 mM EDTA, 1 mM EGTA, 1 mM phenylmethylsulfonyl fluoride, 5 mM sodium pyrophosphate, and supplemented with 1x protease inhibitor cocktail). After homogenization, the protein extract was centrifuged at 18,400 g for 10 min to generate a post-nuclear lysate. The supernatant was transferred to a clean tube. Post-nuclear lysates (2 mg) from JNPL3 (n = 2) and human temporal cortex (n = 2) were incubated with Protein A-conjugated magnetic beads for 3 h at 4°C with constant agitation. The beads were allowed to settle at the bottom of the tube by gravity and then place on magnetic field. The supernatant was transferred to a clean tube. Tau13 (2μL) (BioLegend, San Diego, CA; cat # MMS-520R) was added to the pre-cleared post-nuclear lysate, followed by an overnight incubation (∼16 h) at 4°C. Protein A-conjugated magnetic beads were added and incubated for 4 h at 4°C. The beads were collected as described above and washed four times with Buffer A. The buffer was completely removed and sample buffer containing N-ethylmaleimide instead of reducing agent were added. The samples were not boiled. The assays were repeated using brain tissue from different mice and AD.
Western blot
Immunoprecipitates from AD and JNPL3 mice were resolved in 4–20% SDS-PAGE and transferred to pure nitrocellulose membrane (0.45μm BIO-RAD). The membrane was blocked using 5% dry-milk solution in 1x TTBS (2.5 mM Tris-Base, 15 mM NaCl, 30 mM KCL, 0.02% Tween 20 detergent) for 1 h at room temperature. The nitrocellulose membrane was incubated with 43D pan tau (1:2000) or Tau13 (1:100,000) antibody overnight at 4°C. After incubation with the corresponding secondary antibody for 1 h at room temperature, protein bands were visualized in Li-COR Odyssey Imaging System, using Image Studio Software (ver 5.2).
RESULTS
Tau13 preferentially immunoprecipitated 64kDa tau
The accumulation of aberrantly posttranslational modified tau proteins with an apparent 64kDa molecular weight is associated with pathological tau in specific brain regions and motor impairment phenotype in transgenic JNPL3 mice [17]. The 64kDa tau proteins are also enriched in sarkosyl insoluble fraction, indicating that this tau specie is present in tau aggregates [17, 19]. This shift in molecular weight is mainly due to aberrant phosphorylation of tau proteins [19]. However, other posttranslational modifications, such as ubiquitination, can also contribute to the mobility shift. In western blot analyses of total brain lysate using 43D pan-tau antibody, the 64 kDa tau represents a small fraction of total tau that is almost undetectable in mice without motor impairment phenotype (Fig. 1A; TgN, Input). The main 50 kDa tau protein band detected corresponds to physiological tau. However, as pathological tau progressively accumulates in JNPL3 mice, the 64 kDa protein band can be readily detected in total brain lysate, but the 50 Da tau protein band continues to be the most prominent (Fig. 1A; TgS, Input). Using Tau13 antibodies for immunoprecipitation, we found that the ratio of 50 kDa and 64 kDa tau protein bands was reverted (Fig. 1A; IP). Despite 50 kDa tau protein being the most abundant species in brain lysate of both normal and motor impaired transgenic JNPL3 mice, western blot of the Tau13-immunoprecipitated proteins showed that 64kDa tau proteins were enriched (Fig. 1B). Even in normal transgenic JNPL3, the 64kDa tau protein band was readily detectable after Tau13 immunoprecipitation (Fig. 1A; TgN-IP). Since 64 kDa tau proteins represent a highly posttranslational modified tau specie that could form aggregated tau structures, we incubated brain lysate with beads alone to determine nonspecific precipitation. The results showed that background levels of nonspecific precipitation do not account for the Tau13 enrichment of 64kDa tau proteins, suggesting that Tau13 antibodies can detect normal and aberrantly modified tau specific in denaturing conditions (Supplementary Figure 1), but in solution this antibody preferentially binds aberrantly modified tau species.

Tau13 immunoprecipitates 64kDa tau from JNPL3 brain. A) Representative western blot of Tau13-immunopecipiatated (IP) tau proteins, from brain lysate derived from non-transgenic (NTg), transgenic without (TgN) or with (TgS) motor impairment phenotype, using the 43D pan-tau antibody. Beads alone incubated with brain lysate were used to determine the level of non-specific binding. The arrow indicates 64 kDa tau specie. Input (IN) represents 5μg of lysate before immunoprecipitation. B) The graph illustrates the high molecular weight (HMW) and low molecular weight (LMW) band intensity ratio. HMW encompasses the 64 kDa tau band and LMW is the tau band at 50 kDa (n = 3). *p = 0.01, ****p = 0.0001.
Tau13 immunoprecipitates high molecular weight tau from AD
High molecular weight (HMW) tau proteins accumulate in AD at specific brain regions. These tau species represent aberrantly posttranslational modified tau proteins [1–3, 12]. In contrast to JNPL3 mice, in human, six tau isoforms contribute to these HMW tau species. Western blot analysis of temporal cortex protein lysate from normal aging (NA) and AD cases showed an increase of HMW tau in AD compared to NA (Fig. 2A; Input, NA versus AD). This expected increase in aberrantly modified tau proteins in AD coincide with the neuropathological assessment (Braak stage VI). To determine if Tau13 preferentially immunoprecipitates HMW tau species, temporal cortex protein lysates from NA and AD were incubated with Tau13 antibodies. Western blot was used to detect the immunoprecipitated proteins. The results showed that Tau13 immunoprecipitation enriched HMW tau presence in AD temporal cortex (Fig. 2B). Beads alone used as negative controls showed low levels of HMW tau proteins, indicating that Tau13 preferentially binds to HMW tau in solution.
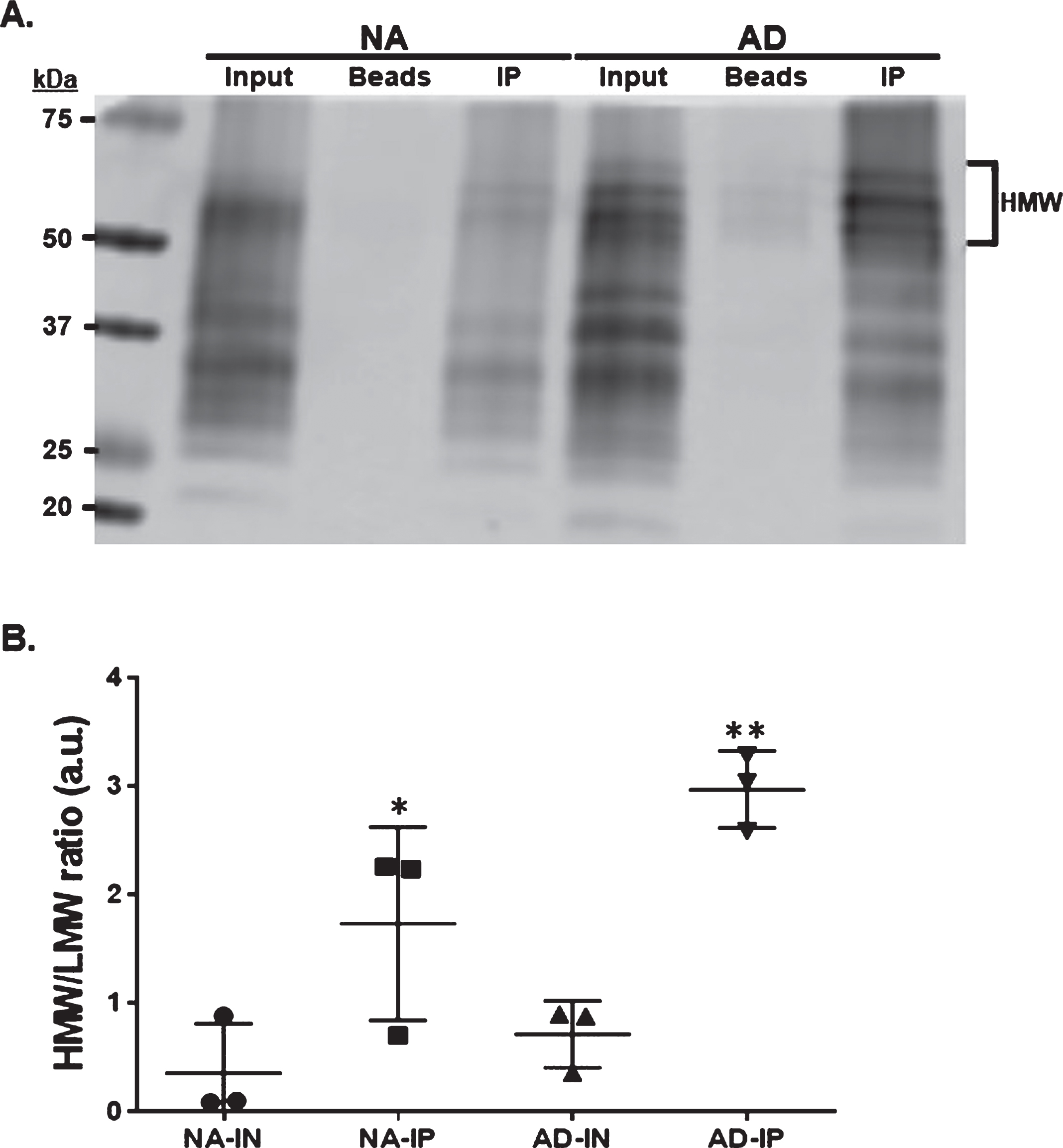
Tau13 immunoprecipitates HMW tau from AD. A) Representative western blot of Tau13-immunopecipiatated (IP) tau proteins, from temporal cortex protein lysate derived from normal aging (NA) or AD, using 43D pan-tau antibody. Beads alone incubated with protein lysate were used to determine the level of non-specific binding. Input represents 5μg of lysate before immunoprecipitation. Bracket indicates high molecular weight (HMW) tau species. B) The graph illustrates the HMW and low molecular weight (LMW) band intensity ratio. HMW encompasses tau bands above 50 kDa tau band and LMW is the tau band at 50 kDa or lower (n = 3). *p = 0.01, **p = 0.001.
DISCUSSION
The progressive accumulation of tau aggregates is directly associated with the severity of the clinical presentation in AD and other neurodegenerative diseases [12]. Biochemical characterization of tau proteins has led to our current understanding of its structural dynamics leading to the formation of pathological conformations. The transition of tau from a monomeric soluble protein to oligomeric forms, that evolves to filaments and ultimately to the ultrastructure(s) known as neurofibrillary tangles, is still poorly understood. Targeting these conformational transitions is an important task in the quest to identify effective treatments against tau-mediated neurodegeneration.
Passive immunotherapy is an approach sought out to interfere with the formation of pathological tau and its interneuronal transmission. Recently, Nobuhara et al. showed that antibodies can be used to block the uptake and transmission in vitro [20]. They use different anti-tau antibodies to immunodeplete pathological tau derived from a tauopathy mouse model (rTg4510) and AD brain [20]. One of the antibodies used in this work was Tau13. The results showed that incubation of brain extract with Tau13 depletes HMW tau, significantly reducing the uptake and propagation of pathological tau [20]. However, they tested other anti-tau antibodies (e.g., 6C5 and 40E8) for their affinities to total and HMW tau [20]. These antibodies immunoprecipitated both normal and HMW tau, suggesting that have equal affinity for both tau species in solution [20]. Here, we show, for the first time, that Tau13 in solution preferentially immunoprecipitates HMW tau produced in a tauopathy mouse model (JNPL3) and AD brain. In contrast, Tau13 does not differentiate between normal and pathological tau when used in western blot and immunohistochemistry analyses (Supplementary Figure 1). This suggests that, in solution, Tau13 epitope is more exposed or available in HMW tau. However, in denaturing conditions, Tau13 binds equally to both normal and HMW tau.
The result presented here points out, at least, two important experimental considerations. First, anti-tau antibodies need to be tested using different techniques before determining their specificity. Second, a combination of anti-tau antibodies that recognized different tau conformations could be considered as a therapeutic strategy to target pathological tau biogenesis at different stages. Thus, a re-evaluation of anti-tau specific antibodies is warranted.